



第六章 科赛尔、利文和四核苷酸假说
“四核苷酸”最初是指核酸分解产物中,出现等分子百分数的四种核苷酸或相应的含氮衍生物,因此用来描述那时所知道的核酸,当时认为核酸就是这样简单地组成的一种分子。现在已确定核酸是一种多核苷酸,性质很复杂,但这个名词仍在原来意义上用于这类多核苷酸,因为它们所含的核苷酸符合四核苷酸的比率。
后来,由于认识了这种复杂性质而取得了进展,这个名词就用来表明一个单位,即由四种核苷酸中的每一种核苷酸分子所组成的单位,这种核苷酸通过自身的反复组合形成了多核苷酸,这似乎意味着这样结合的模式在整条多核苷酸中是完全一致的。最后,已推测在每一个这样的单位里,四种核苷酸总是以固定方式组成不变的序列。作为这些假说的合乎逻辑的结果,提出了某些多核苷酸作为“多聚四核苷酸”的概念。
这样,“四核苷酸”这个名词有了一系列递变的含义,从分析结果的统计学表达到一种确切的化学结构的定义,为避免混淆起见,使用这个名词时一定要规定其含义;“统计学的”和“结构的”四核苷酸就是由此而提出的两个术语。现在正是综述和评价这些解释所依据的证据的时候,特别是因为多核苷酸在生物学领域中的重要作用,所以必须清楚地认识不同来源的多核苷酸之间相似或趋异的种种可能性,因此涉及核酸的各种猜测也许只是以公认的化学知识为依据。
(古兰德、巴克和乔丹,1945,184~185)
尽管在奥尔布雷克特·科赛尔领导下,在十九世纪对核酸作了研究,但当菲伯斯·利文于1900年进入这个领域时,有关核酸的化学还远远没有弄清楚。在以后的三十年间,这位伟大的化学家使这个课题从混乱中摆脱出来,使核酸不再在概念上同蛋白质纠缠在一起。1931年,他出版了一本书,综述了核酸的化学。这是他聪明地采用有机化学技术的一块“纪念碑”:特别是戊糖。但是,这项成就的代价是接受了一个简单的结构公式,即碱基有一个固定位置,有机化学家认为这是可以辨认的。
从本章里我们将看到怎样根据在核酸分子中只有四种碱基的发现,提出这个概念的。当同根据经验公式得到的分子量一起考虑时,结论是四核苷酸。我们也将看到,利文如何成功地把核苷和核苷酸——肌苷和肌苷酸——的一级结构于1909年应用于RNA,从而有力地支持该假说;当二十年后利文把肌苷酸结构推广到DNA时,这个假说得以解释各种现象。这个假说是如此深入人心,以致后来利文和孚尔根认识到DNA和RNA的大分子性质后也不能动摇它。四核苷酸假说是在开始探索未确定领域时的工作假说的一个范例:它是开掘这个课题的工具。这些假说取得了成功并变得如此牢固,以致要有一次革命才能消除它们。四核苷酸假说则有双重革命。一是改进由哈默斯坦引进、由米尔斯基和西格纳所采用的抽提技术,同时使用测定高分子量的可靠方法。再一次革命则是查格夫把层析法用于核酸。在查格夫的工作之前,核酸化学家对碱基分析常常达不到四核苷酸的要求,通常都漠不关心。他们总是可以说分析方法过于粗放了。这里就像大分子概念的情况一样,我们看到了技术革新所产生的有力影响。
与此同时,日益认识到蛋白质的惊人的复杂性,结果出现了我们可称之为“中心法则的蛋白质版本”。这个学说认为,生物的特异性来自蛋白质,特异性的各种变化来自蛋白质里的氨基酸序列。
这不是说核酸不重要。绝非如此。科赛尔把核酸同新的原生质的形成,同生长——合成联系起来。但是,对于每一种生化分子来说,他明确断定组蛋白更为重要;关于组蛋白,他建立了一种生化分类学以及有关蛋白质结构和合成的理论。那么,科赛尔的生化个性的概念是什么?怎样构思出四核苷酸假说的?为什么这个假说能保持如此之久?
科赛尔的生化个性概念
“自然科学家和医生联合会”访问他家乡罗斯托克镇时曾带领过植物园游览队的这位学生积极分子,曾在德国施特劳斯堡大学受业于著名植物学家门下,但他父亲讲明了要他去念医学而不念植物学,然而他不是看不到自己工作的生物学意义的那种人。当霍佩-赛勒争取到科赛尔从事生理化学研究去分离核素的水解产物时,难道我们能相信科赛尔会无视他的化学研究的生物学意义吗?绝非如此。他的论文总是可隐约地看出其生物学意义,一有可靠的化学数据就准备表达出来。他的同事S.埃德尔巴歇曾说过科赛尔渴望从“统计学的研究”转向“动力学的研究”(埃德尔巴歇,1928,13),就是说,从化学分析转到代谢路线。其实,对生物学意义的关心,正是他继续探究从拟核素(磷蛋白)到核酸的代谢路线的主要原因!
科赛尔从生理学研究中得出了正确结论,即核酸既不是作为贮存的化合物,也不是作为肌肉收缩的能源,而是同新鲜组织的合成作用有密切关系。早在1885年,他承认查卡里乌斯提出染色质像是核素的化学特征的证据,科赛尔说:“……这种物质除了白蛋白和磷酸外,生成次黄嘌呤、鸟嘌呤和黄嘌呤。”(1885,1929)八年后,他把染色质是游离酸的可能性包括在内:“组织学家所说的染色质主要是带有一点白蛋白的核酸化合物,在某种程度上这也是完全‘游离的’核酸。”(1893,158)
刊登在《现代文化》上的一篇论文中,他论述了化学同生理学的关系。
核酸只出现在细胞核里,其实只出现在细胞核的一个部分里,很久以来组织学家称之为“染色质”,以说明它倾向于吸收碱性染料,与细胞核其余的形态学组成成分适成对照。这个事实对化学同细胞学说的关系来说有重大意义,因为它使我们除了认识细胞的基本器官的形态学特征外,还了解其化学特性。对于构成细胞核染色质的特定基本单位的了解,必将成为研究这个器官的化学活性的基础。可是,主要在涉及生长和原生质替换时才可看到这种活性。
(科赛尔,1913,383)
在二十世纪的读者看来,这真是突然由正确转入了错误,但这是了解科赛尔想法的一条线索!后面我们将评论这种思想方式;它带有五十年代以前的生物化学的标记——代谢路线的局限性,这是从“统计学”化学到“动力学”化学的自然发展,也是从有机化学进人生理化学的有机合成的逻辑延伸。那时的计划是要弄清反映细胞内部活动的合成化学和降解化学,遗传信息是自然界的既定事实,研究它被认为为时尚早。
那时,科赛尔很高兴地让学生海因去研究有丝分裂的化学,希望在染色体的染色性同核酸与组蛋白的结合状态——当与组蛋白“解离”或联结疏松时被碱性染料深染,“不解离”时则淡染——之间确立平行关系,这很难说是对有丝分裂作为复制染色体化学信息的一种机制怀有很大兴趣。不,刺激细胞分裂和形成新鲜原生质的化学反应确实令人感兴趣。
另一方面,科赛尔是细胞化学的“构造单位”概念的创始人。根据这个概念,细胞代谢可分解成许多初始分子——“构造单位”的相互连接和分离,由此合成和降解为次级复杂分子。在胚胎中,许许多多“构造单位”连接在一起产生了复杂蛋白质;在精子里,这些复杂的蛋白质已被除去,留下了鱼精蛋白——最简单的蛋白质。核酸(在嘌呤环和嘧啶环中)以及鱼精蛋白和组蛋白(在精氨酸和组氨酸中)都有大量C=N键,这给科赛尔以深刻印象。什么地方出现生长,什么地方就大量生成这些化合物,这个事实向他表明了这些C=N键的特定的合成活性。在这里,帕弗吕格的多聚体理论的影响是值得注意的。
可是,科赛尔怎样使“构造单位”排列在多聚体分子里?他是否像艾米尔·费希尔那样,根据链的顺序看出了化学个性呢?无疑他是这么做了。他说,在了解蛋白质时涉及结构化学,“但这不是原子在分子里的相对位置问题,而是把分子或‘构造单位’集成一个较大单位——蛋白质的问题”(1913,383)。他没有想到去抽提短链多肽,这是可以理解的;但费希尔鼓吹的这条研究路线,在四十年代末最早的序列分析中达到了顶点。科赛尔发现蛋白质分解产物之间有恒定的定量关系后,坚信“构造单位”存在这种排列。他曾引进银盐技术,但1914—1918年战争期间白银稀缺时他发展了萘酚-黄-S法,他采用范·斯里克新的定氮法。他说,得到的数据
……迫使我们得出结论:不同性质的蛋白质的化学结构也完全不同。我们也必须作出结论说,各种分解产物的原子基团,确实是这样存在于蛋白质分子中;因为同一种蛋白质水解后,总是得到数量一定的产物。
(科赛尔,1912,67)
这是1911年科赛尔在琼斯·霍普金斯大学所作赫脱基金讲演中的一段。1911年他作的哈维讲演更为精彩,下面引用其中的一段,他在谈到用食物分解的蛋白质在体内重新构成蛋白质时说:
我想把在动植物体内蛋白质的重新排列比作编成一列火车车厢。 当蛋白质在身体里运行时,有些部分会落在后面,有些则在这里或那里添上新的部分。为了充分理解这种变化,我们必须记住蛋白质是由“构造单位”以不同方式构成的……能够参与构成蛋白质的“构造单位”的数目,大约与字母表里的字母一样多。我们考虑到字母的组合可以表达无限量的思想时,我们就能理解在蛋白质所占有的一个很小空间里可以记下生物体的极其大量的属性;就能理解性细胞的蛋白质可在某种程度上包含物种的乃至个体的全部性状。我们也就能理解决定蛋白质结构的任务是何等重大,以及生物化学家为什么如此锲而不舍地从事蛋白质的分析。
(科赛尔,1911,45)
不知道核酸的情况又是怎样?科赛尔及其学生们从酵母和胸腺的核酸中分离出嘌呤和嘧啶,证明羟基嘌呤(黄嘌呤和次黄嘌呤)是由鸟嘌呤和腺嘌呤脱氨基生成的,并发现来自动物——无脊椎动物和脊椎动物的精子、血球细胞和胸腺——的核酸十分相似,但同酵母的核酸却有明显区别。1902年,奥斯本和哈里斯第一次从高等植物中制备出核酸,证明像酵母的核酸一样含有尿嘧啶,以代替动物核酸中的胸腺嘧啶;含有五碳糖而不是六碳糖(认为这是动物核酸里的糖),显然至少有两种核酸。也许还不止两种;也许是鸟苷酸、胸腺苷酸和腺苷酸;如果是这样,则在这种同聚核苷酸与酵母核酸或胸腺核酸之间可能有复合核酸。十九世纪末,就是用这些可能性来解释从胰脏得到的古怪的核酸。此时,一位富有进取心的年轻的瑞典化学家伊瓦尔·克里斯琴·班进入这一领域,他除了对胰脏核蛋白的同一性提出许多疑问外,还怀疑科赛尔的一些工作。科赛尔发现自己正处在一直想避免的公开争论的情况之下。这位出生于波罗的海罗斯托克港的普鲁士人性恪温和而内向,不善于当众讲话(埃德尔巴歇,1928,4)。他可能因为同伊瓦尔·克里斯琴·班争论,较少注意到核酸而更多地去注意蛋白质。另一方面,米歇尔死于1895年,留下的鲑鱼鱼精蛋白是一种没有确定的物质。这些物质以及第一次由科赛尔报道的组蛋白使他未能得到核酸——这是各种“构造单位”的丰富来源。
科赛尔并未把核酸看成极小的分子。他确实认为核酸的每一个磷原子有一个碱基和一个糖的“构造单位”,而且有4种碱基。因此,他的结论是核酸分子含有12个或12的倍数的“构造单位”(科赛尔,1913,383),在他所作的诺贝尔奖获奖演说中,把核酸描述为“至少有12个构造单位的一种复合物,但在活细胞里的结构可能更大些,因为有些观察指出在器官里,几个这样的复合物彼此结合一起”(科赛尔,1910,399)。可是,在降解产物中鱼精蛋白和组蛋白竟如此之多!他用以计算碱性氨基酸的银-氧化钡法,比估计嘌呤和嘧啶时所需的粗放而剧烈的方法又可靠得多!正是在这方面,科赛尔认为有希望达到他铭记在心的目的,即用化学转化来说明生长和分化,根据化学上的区别来说明形态上的区别(参阅他的专著《鱼精蛋白和组蛋白》,1928,vii)。
科赛尔最有成效的工作是在柏林马堡大学的十个愉快的年头里完成的。科赛尔住在富丽堂皇的生理学研究所中的教授住宅里,这是普鲁士文化部长为科赛尔的前任屈尔茨建造的。在他的研究生中间,英国化学家T.H.米尔洛研究类核素,海因研究有丝分裂的化学,美国人A.P.马修斯研究分析鱼和无脊椎动物的核酸,库契研究鱼精蛋白,P.A.利文和F.P.H.斯托伊德尔也来到马堡大学。此时,科赛尔仍积极参加他的学生们的研究工作,而且在他的公寓里非正式地款待他们(M.E.琼斯,1953,85)。问他为什么于1905年离开马堡赴海得堡时,据说他的回答是因为不喜欢他的同胞——普鲁七人的趾高气扬的方式(马修斯,引自琼斯,1953,86)。1913年他的妻子去世;1914年第一次世界大战使他与国际科学界以及在美国的亲友断绝了往来。对他来说没有必要保持“德意志的显要地位”(达金,引自琼斯,1953,93),也没有必要追随加强潜艇的政策。他日益趋于内向,他为德国所发生的事件感到羞愧。他在柏林时期的主要技术助手海尔·皮特里慢慢变成了海得堡实验室的负责人,这是一位具有“兴登堡式的外貌、体质和方法”的人物(肯纳韦,1952,394)。科赛尔的研究方式和态度
……是粗鲁的;他会嚓一下立正并说“嗳,您的研究工作怎么样了”?然后,人们要征询他的意见;不管提出哪一种工作方针,肯定会引出他的一句口头禅——“这有利也有弊”。人们得到的印象是他对任何一种化合物都不感兴趣,或是对没有得到“十分有趣的盐类”或某种晶体的研究人员不感兴趣。
(肯纳韦,1952,394)
阿尔伯脱·马修斯回忆他与科赛尔在马堡相处一段日子的情况却全然不同。对他来说,科赛尔是良师益友(马修斯,1936,ⅷ)。
科赛尔的影响
氨基酸分析以及嘌呤和嘧啶分析这一对孪生技术,无疑是由科赛尔建立的。据说他曾把鱼精蛋白画成碱性氨基酸的链,同淀粉的多糖链相类似。当马修斯问他为什么不打算用氨基酸去合成这种链时,他的回答是:“曾经有过这种想法,但把它转告了艾米尔·费希尔,因为他的设备较好,可做这一工作,而且对有机合成也比自己更在行。”(琼斯,953,87)从这一说法无法断定在多大程度上由于科赛尔的促进,费希尔才去从事这一重大研究。当我们提出遗传物质是什么这个问题时,科赛尔的著作中未作明确的讨论。他至多只是寻求一种化学分类学。正是这种希望促使他向马修斯提出建议,他要研究无脊椎动物海胆的核蛋白。这种低等动物的精子的化学成分,应比鲑鱼、鲱鱼的精子更简单些。但并非如此,马修斯发现海胆的核酸,完全像鱼的精子中的核酸,它的鱼精蛋白(海胆组蛋白)从氨基酸的种类来看,非但不少而且更为复杂(马修斯,1897,411)。至于鲱鱼的精子,其头部的鱼精核蛋白——他认为是精子的染色质——的组成可以写成C30H57N17O6·C40H54N54P4O27。可是奥古斯特·魏斯曼认为,染色质“应是身体里最复杂的化合物,因为它含有(身体里)所有其他分子的干分子”(同上,411)。因此,精子头部里的染色质应是这种物质的最复杂的形式,但事实上,这是迄今所知的最简单的形式!马修斯由于笃信核酸是遗传物质,所以对此持极端怀疑的态度。现在引用他给考德里的《普通细胞学》所写文章中的一段话:
细胞学家称之为染色体的结构,大多数人认为是所有遗传性状的载体,有些人甚至认为每一个性状是由一个明确的单元或基因所代表;这种结构在固定的经染色的切片中显现出来,它只不过是经常用以染色的碱性染料与核酸形成的盐而已。除了染料以外,还不能肯定是否还有某种蛋白质 然后,核酸又是什么呢?有许多种核酸呢,还是只有一种?
常常在讨论染色质的组成但又很不完备,因为我们了解得太少,无法支持染色体是由基因所组成的假说。从化学观点来看,检查鱼类精子得到的资料是这一理论最好而又最令人信服的证明。在身体里应是最复杂的染色质,但在这里我们找到的却是最简单的;就是说,这是一种可以给出分子式的鱼精蛋白和核酸的盐类……如果染色体是由完全不同的基因所组成,那就根本不可能表现为这么简单而确定的组成。各种细胞的核酸似乎是相同的。当然,它可能是不同的,但事实上它有同样的物理性质、分析数目、旋转力等,这些也许表明不存在几种不同的核酸。肯定不会有好多种。因而促使我们去寻找染色质中的另一半——蛋白质中的差别,如果存在这类差别的话。在我们检查过的精子中,找不到这一理论预期会有的差别,这是值得注意的。我们老是找到一种鱼精蛋白,只含3~4种氨基酸。同时,即使在这种鱼精蛋白里,其排列也可能有相当大的差别;但没有化学的证据表明存在这种差别……
(这些化学事实)如能支持这种观点的那也是很少的。笔者认为,这里能得出的有价值的看法是精子的染色质只不过是精子的染色质;而且卵细胞的染色质也只不过是卵细胞的染色质。它们不是神经、肌肉、上皮细胞的染色质。
当这些染色质结合起来时,可以生成身体里所有的染色质,或至少在这些染色质的生成中起重要作用;不能把它们看成是包含着所有能被制作出来的产物样本的博物馆,因为这些产物的数目对于每一种染色质来说实际上是无限的,如发育条件稍有改变细胞就会产生差别,偶然事件如植物的虫瘿等也会产生差别,所以染色体没有空间来容纳所有这些样本。可是,如果染色质能在没有样本指引下制造出另一类染色质,那么为什么不制造出所有染色质呢?为什么总要有某些样本?对这种情况各有各的看法。必须牢记的是,只有持下列主张的人才有举证的义务,这些人主张染色体是包含身体里所有细胞的所有染色质样本的博物馆,所包含的不仅是在生命发育期间的所有染色质,而且还有老祖宗遗传下来的无数珍藏,我们上代还未表现出来的数目无限的样本,以及目前通常所说的潜伏性状。它们无疑是隐藏在染色体的小阁楼里,一有机会就产生出来。
(马修斯,1924,75,89,90)
1936年,马修斯又改变了说法。在来自不同组织的细胞之间和不同物种的精子之间,在鱼精核蛋白的蛋白质部分和组蛋白之中看到了预期的化学差别。“每种鱼的染色质都有自己的那种蛋白质;也许正是蛋白质组成成分的这种差别,使之成为不同物种的蛋白质。”(马修斯,1936,108)科赛尔的学生库尔特·费利克斯保持用这种预料来说明鱼精蛋白的遗传多样性,他是海得堡蛋白质研究所所长,鱼精蛋白的一位大权威。但遗传学家主要受A.P.马修斯的影响,E.B.威尔逊和斯特拉斯伯发表了他的一些观点。如果我们问马修斯的说法的背后是什么,回答显然将是四核苷酸假说和P.A.利文的工作。
四核苷酸假说
根据上面所说核酸未出现足够的变化可用来解释生物化学的特性,就能清楚地表明用还是不用这个假说。蚕丝丝心蛋白主要由2种氨基酸(甘氨酸和丙氨酸)组成,血红蛋白含有所有20种氨基酸,两者呈鲜明对照;核酸则没有这种情况。但核酸概念在确立四核苷酸假说的前后有一明显区别。后来的概念所代表的不止是一种结构,并且是代表研究结构问题的一种特定方法。在四核苷酸假说提出以前讨论核酸结构时,在科赛尔的学生阿尔伯特·纽曼称之为a型和b型凝胶生成和非凝胶生成的核酸钠盐之间有一种区别。纽曼修改了理查德·奥尔特曼的抽提法,很快得到了很好的核酸样品。奥尔特曼、科赛尔和纽曼所引入的这些改进旨在得到真正去除了蛋白质的核酸,用来解决核素、类核素和蛋白质之间关系的争论。他们所作的这些改进使得核酸很容易抽提,所以在1900年左右核酸化学成为非常普遍的课题。
纽曼发现了非凝胶生成的b核酸——他把它看作是凝胶生成的a核酸的解聚产物,对我们来说,这个发现是十分重要的。我们读到他能通过热处理的时间来控制降解程度时,提高了我们对他的观察的热情。而且,我们高兴地发现他把这个过程同在蛋白质和多糖中的类似观察联系起来,因此,“很容易用解聚作用来解释化合物a的胶化牲。在淀粉、树胶和白蛋白化合物的某些方面看来都有类似的关系”(纽曼,1899,375)。
七年后,理查德·伯里安把纽曼的结论概括成下列的核酸逐步降解(伯里安,1906,784):
1.解聚作用,核酸a变成核酸b。
2.除去鸟嘌呤产生施米德伯格所说的半核酸和纽曼所说的胸腺核酸。
3.完全除去嘌呤产生施米德伯格所说的核质磷酸,以及科赛尔和纽曼所说的胸腺核酸。
4.已去掉嘌呤的分子全部分解为基本组成成分。
早些时候,伯里安讨论了核酸可能有的分子量,并断言虽然经验公式得出的相对分子质量为127.2,但米歇尔证明核酸扩散极慢,表明“它的分子量公式可能还要乘上一个数”(伯里安,1904,80)。关于解聚作用的这些讨论被搁置一边,这种情况一直维持到三十年后孚尔根的研究工作发表时止。与此同时,由于奥斯本和哈里斯以及菲伯斯·利文采用剧烈的抽提方法使问题变得更为简单。利文在描述科赛尔和纽曼改进奥尔特曼的技术时,作了如下评论:“对采用强烈方法分离核酸和蛋白质的担心消除了。”(利文和巴斯,1931,251)在同一处他还谈到了他们的早期工作是“在核酸很易变化的传统信念影响之下,因此他们避免用加热……”。从亲本分子里分离嘧啶和嘌呤,采用强烈的方法确实曾经是而且目前仍然是必要的,亲本分子的结构看来只能在牺牲它的多聚性质的情况下才能确定。不用四核苷酸假说,就无法澄清这个问题的惊人的混乱。要是不用同样处理的材料去测定分子量、碱性、一级电离和次级电离的磷酸根数目,那该有多好!这种片面的进展始于奥斯本和哈里斯的工作。
三核苷酸核酸
十九世纪时,核酸的公式都是有四个磷原子的经验公式;但第一个指出分子里四个磷原子同四个碱基相结合的是著名的美国蛋白质化学家托马姆·奥斯本,他同伊萨克·哈里斯一起在纽黑文的康涅狄格农业研究站的实验室里工作。1902年,他们从小麦胚中分离得到大量物质,从而引进了小麦核酸这个名词。因为他们指出这同酵母核酸很相似,所以可断定这是RNA,但他们对抽提出这种RNA的胚组织的描述却令人迷惑不解。他们说,细胞核是如此之大以致细胞不能再容纳别的东西(奥斯本和阿里斯,1992,86)。
这些学者骄傲地宣称已对小麦核酸作了彻底研究,已能“确定这种分子的可能的构型”。不幸的是,在得出最后结论之前,他们被迫中断了这项研究。由于他们把到那时为止的发现发表在科赛尔主编的《霍佩-赛勒杂志》上,所有学习核酸的学生评价他们的结果必然会对以后的发展产生深远影响,正如琼斯所说,那时是把小麦核酸“作为在所有核酸中研究得最彻底、最精确的一种”(琼斯,1920,49)。
在他们做的10种基本元素的分析中,他们认为最纯的样品给出的经验公式是C41H61N16P4O31。根据银盐,他们认为碱基为6。关于腺嘌呤和鸟嘌呤的百分比,他们写道:
……碱基无疑是以等分子比出现的。小小的差别(鸟嘌呤太多)起因于完全分离碱基时的困难,因为在鸟嘌呤沉淀中混有一定量的腺嘌呤。
(奥斯本和哈里斯,1902,104)
因为每克核酸中得到的这两种碱基相差达9%,所以这个结果不能作为证明或是否定等分子比的依据。
奥斯本和哈里斯根据科赛尔的方法,把小麦核酸放在20%的硫酸里,用150~160℃高压蒸汽处理2小时。他们得到一种嘧啶,它相当于1901年科赛尔的学生阿斯科利在酵母核酸中发现的尿嘧啶,那一年艾尔米·费希尔和哈根巴赫测定了它的熔点。关于它的产量,奥斯本和哈里写道:
核酸分子里的一个尿嘧啶分子,相当于核酸分子的8%在上面提到的实验里,发现尿嘧啶超过11%,我们一定会由此得出结论:核酸分子里至少有两个尿嘧啶分子。所发现的量还低于两个分子所要求的16%,但如果考虑到制备的步骤很多,磷钨酸和氢氧化钡的沉淀很多时,就会明白不可能得到很多的尿嘧啶。
(奥斯本和哈里斯,1902,109)
他们得出的结论是每个核酸分子是鸟嘌呤1:腺嘌呤1:尿嘧啶2。同时承认,有一种物质的量很少,它可能是1894年科赛尔和纽曼发现的胞嘧啶,他们认为核酸里只有一种嘧啶,并计算相对分子质量如下:
相对分子质量
1分子鸟嘌呤 151
1分子腺嘌呤 135
2分子尿嘧啶 225
3分子戊糖 450
6分子羟基 102
4个磷原子 124
3个氧原子 48
1234
减去用于磷酸键的7个氢原子 1227
但是,真正的相对分子质量是1397,“所以那里还有12.2%未分解的残基,这也许是没有辨认出的降解产物,但我们还未能令人满意地证实这一点”(奥斯本和哈里斯,1902,119)。换句话说,他们偏向于认为含有五核苷酸:腺嘌呤、鸟嘌呤和胞嘧啶(?)各一个分子和两个分子尿嘧啶。在图6.1的式子中,他们提出了小麦核酸的结构,胞嘧啶(?)用X表示。
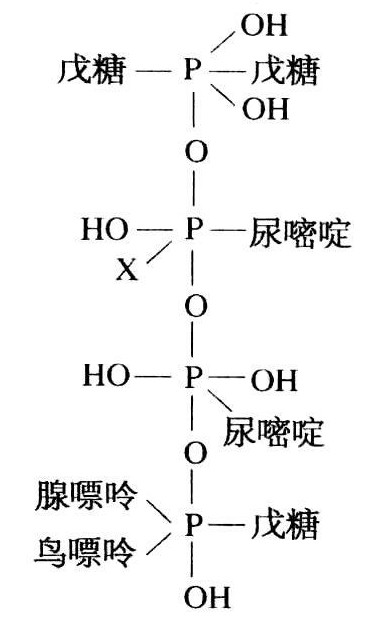
图6.1 小麦核酸(录自奥斯本和哈里斯)。
这个式子——我们认为是很了不起的——使奥斯本和哈里斯得以解释嘌呤比较容易从分子上断裂下来,以及解释他们的错误观察即在这个过程中去除一个磷原子。对我们来说,他们提出的结构正好表明在得出各个基团之间有键连接的现代概念之前,还有多少工作要做。只要仔细读过他们论文的最后一节,就不难看出他们如何强烈地受到真正核素同类核素之间有一条直接代谢路线的信念的影响。他们之所以对核酸结构感兴趣,看来正是由于他们想在小麦胚组织中证明有这种关系——对植物界所做的也正是利文打算在动物界中要做的。
三核苷酸核酸中的胞嘧啶
附近的耶鲁大学谢菲尔德实验室里,亨利·惠勒和特里特·约翰逊在合成尿嘧啶。他们要把合成的产物同奥斯本的产物作比较,所以把奥斯本的晶体除去水分——奥斯本不再把尿嘧啶晶体送给他们了——可是他们说得到的针状晶体是“尿嘧啶和胞嘧啶的等量混合物”(惠勒和约翰逊,1903,305)。这说明奥斯本的和哈里斯的尿嘧啶不纯。他们还看到了一种未经鉴定的成分X并认为它可能是胞嘧啶,他们相信存在两个分子尿嘧啶和一个分子嘌呤,但这无碍于确立四核苷酸,因为另一些事实立刻引起人们注意,而且奥斯本和哈里斯使人们对小麦核酸有一个清晰的印象:每四个磷原子有两个嘌呤分子和两个嘧啶分子——腺嘌呤和鸟嘌呤以等分子百分比存在着。简言之,认识到核酸里只有四种含氮碱基:对胸腺核酸来说是腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶;酵母核酸则是腺嘌呤、鸟嘌呤、胞嘧啶、尿嘧啶,请来了四核苷酸公式。诚然还有另一个证据——每四个一级基团释放出一个次级磷酰基(利文和西姆斯,1925,1926),这是用一条打开的四残基链(利文和西姆斯,同上),或用图6.2中打开成一条链的环状分子(高桥,1932)来解释。早期根据扩散估计出的相对分子质量是1360到1700(米尔贝克和乔帕斯,1935),大体上符合于四核苷酸——1357。当利文鼓吹磷酸酯-糖键构成DNA骨架(利文,1921)和RNA骨架(利文和西姆斯,1925)这一重要阶段时,就有了支持四核苷酸的三方面证据。
人们也许会问,如果早期除了比伯里安、纽曼和米歇尔以外有更多的人认识到核酸的多聚性质,将会产生什么结果。我是怀疑的,因为这些研究者会有我们所说的大分子概念吗?更有可能的是他们会按照我们今天所说的寡核苷酸去考虑问题的。利文和巴斯正是在这个意义上讨论哈默斯坦和孚尔根在专著中所说的“高级”核酸(1931,302~303)。因此,我们完全有理由更多地注意第二章里的大分子概念。
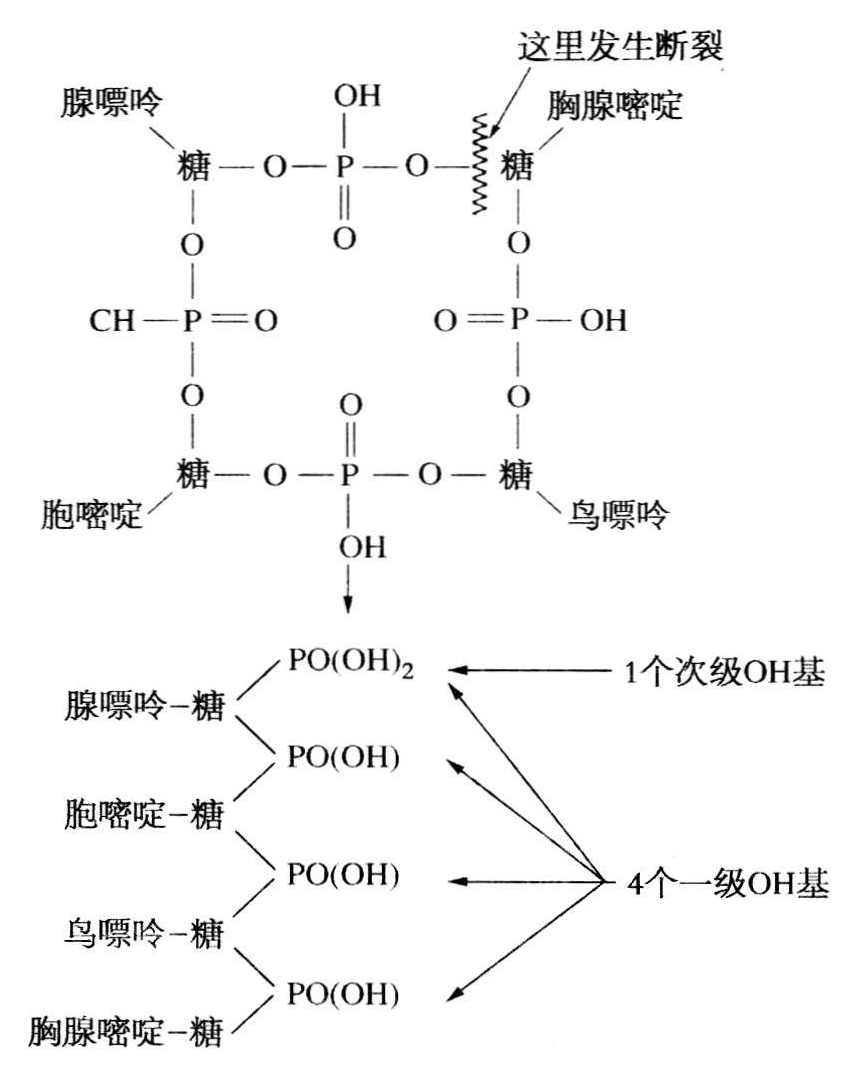
图6.2 设想一个环状四核苷酸如何打开成为一条链,这条链由四个一级、一个次级磷酸根所组成。
很久以前,怀亚特指出以四种碱基盐的溶解度不同为基础,得到的 DNA 组成成分的一些早期分析数据竟是出奇的好。他注意到这些数据偏离查加夫的碱基比的预期值,也偏离四核苷酸结构的预期值,而且偏离程度相同(怀亚特,1950,143)。表6.1列出了1902—1950年间得到的一些数据。
表6.1 1902—1950年间核酸的碱基分析克分子比

第一个声称支持嘌呤和嘧啶的等克分子比例,并以四核苷酸作为工作假说的不是利文而是斯托伊德尔,但他的数据与这个假说不符,胞嘧啶的数值只有该结构所要求的54%(马克姆和史密斯,1954,6)。不幸的是,利文于1908年提出两个嘧啶对三个嘌呤后,翌年就转向斯托伊德尔的观点,为了支持把斯托伊德尔的理论值(四核苷酸的预期值)作为斯托伊德尔的实验结果(马克姆和史密斯,1954,7),在1931年重复了他的错误,当时他在专著里宣称“埃林豪斯根据完全不同的、巧妙的物理学方法,即比较核酸各个组成成分与核酸本身的氧化热,具体证实了四核苷酸结构”(利文和巴斯,1931,277)。人们不知道埃林豪斯的结果实际上是符合于四核苷酸呢,还是符合于一个巨大的大分子。
最后,我们不要忘了利文提出的磷、糖和碱基在核酸(次黄嘌呤核苷酸和次黄嘌呤核苷)中的排列模型。1909年,他已论证了次黄嘌呤核苷酸最可能的结构是一个嘌呤通过糖苷键同糖连接,然后通过羟基以酯键与磷酸连接(利文和雅各布斯,1909a,336)。两年后,他提出了图6.3所示的结构,糖和碱基之间的连接没有放在N9位置上,而是错误地放在N7上。
1909年,利文和雅各布斯还从酵母核酸中分离出鸟嘌呤核苷和腺嘌呤核苷(1909b;1909c),并提出核酸分子是由四个核苷通过缔合的磷酸根连接而成。他把这种四核苷酸命名为“多核苷酸”,且认为胸腺核酸有类似的结构。
我们可称利文和雅各布斯那时提出的核酸分子公式为“四核苷”结构,这种结构中骨架的连接是搞错了,图6.4表示一个二核苷酸。

图6.4 酵母核酸的二核苷酸(录自利文和雅各布斯,1909c,2703)。
直到利文从酵母核酸中得到所有四种核苷酸时,他才引进了现在的RNA中的糖-磷酸骨架(利文,1919)。如果那时他就能证实DNA也是这四种核苷酸,那么就会成功地把这种结构推广到DNA和RNA这两种核酸。可是,DNA比RNA更不稳定,利文和雅各布斯虽然正确地指出这起因于糖(1912),但直到1929年利文才取得成功。
植物核酸和动物核酸
一些评论家最近从琼斯的文章里发现一种有趣的说法,他说核酸“可能是生理化学中了解得最清楚的领域”,以及“……自然界只有两种核酸,一种得自动物细胞的细胞核,另一种得自植物细胞的细胞核”(琼斯,1920,vi,9)。第一个说法提到核酸代谢特别是分解代谢的知识。琼斯的第二个说法一定要同有关核酸的结构和种类的早期文献的混乱背景相对照,例如文献中提到,类核酸和胸腺核酸(科赛尔和纽曼,1893),胰脏的鸟嘌呤核苷酸(班,1898),核素基(施米德伯格,1900)等,当时关于嘧啶——伯里安早期坚持嘧啶来源于嘌呤——特别是尿嘧啶的一级性质和次级性质存在争论,尿嘧啶是否由胞嘧啶脱氨基生成?在琼斯撰写上述文章时,只有胰脏的鸟嘌呤核苷酸尚未得到解答。
最后是E.乔帕斯研究了这种酸,从而打破了植物核酸和动物核酸的分类。1929年,乔帕斯作为洛克菲勒研究员与利文一起工作,利文于1931年的专著中记述了这一点,但在核酸的分类一节仍写了两个标题:酵母核酸(植物核酸)和胸腺核酸(动物核酸)(利文和巴斯,1931,260)。
我们虽可用这本书已在好几年前写成来解释这种情况,书是在1925年开始撰写的(利文,1925),但事实上利文在以后也没有修正,这使我们感到当时那种分类法可说是积习难改。1920年,艾因纳·哈默斯坦指出胰脏核蛋白里的糖是戊糖,他的学生乔帕斯逐渐加强了这种说法(哈默斯坦和乔帕斯,1922;乔帕斯,1924,1928),并在利文的专著出版之前,在洛克菲勒研究所作了明确的验证(利文和乔帕斯,1930)。在这里,我们不是看到了简单假设的诱人特性吗?不是看到了化学家们要在化学基础上区分动植物界的愿望得到实现吗?谁到了1931年还以自己的全部权威充当这种分类的后台,谁在1938年当自己已转向多聚四核苷酸结构时还充当四核苷酸结构的后台,谁不就应该贴上这两个著名假设的标签吗?如果有这样的人的话,那么利文就是要遭到历史学家鞭挞的人。
菲伯斯·阿伦·利文
像利文这样一位核酸的简单结构和分类的主要支持者,看来不太可能在历史上湮殁无闻的。这位来自圣彼得堡(列宁格勒)阿历山大·鲍洛丁化学研究所的出类拔萃的青年俄国犹太人,于1891年22岁时随同家庭来到美国。1896年担任生理化学副教授,在纽约州萨伦内克实验室作为化学家研究结核病,他早年曾在那里患过结核病。最后,应新的洛克菲勒医学研究所所长西蒙·弗莱克斯纳博士的邀请,他找到了安身立命之处,开始时在东五十街127号的研究所里,这所房子还只用了一年;1906年搬到东河的新建方达大厦一楼。处于这样无与伦比的优越地位,利文的研究年复一年地发展,他承担新的研究计划,得到越来越多的经费,希望到研究所来在他指导下工作的年轻科学家也日益增多。
利文同弗莱克斯纳虽然仍是亲密好友,但这位俄国科学家在经费使用上的独断专行激怒了美国人所长,造成了两人之间的不和。从美国哲学学会图书馆保存的信扎中可看出这种情况,弗莱克斯纳给利文的信有几段是这么写的:
昨晚执行委员会会议决定,今年不再增加您的工作人员。
(1916,6,3)
我想来找你了解关于你那个部门的供应和经费。(1918,4,27)
我决不会说……我被你自身同你的设备之间的区别而大受感动。把真正的进展只归因于乙醇、乙醚等是拙劣的判断……
(1919,2,7)
如果考虑到(由于战事)生产可能不正常而多买了酵母和其他一些东西,以致过分影响到(你的经常预算),那么你必须预见到意外支出还会使你的预算发生麻烦……你已遭到了麻烦,你又在麻烦执行委员会,它要你解释准备怎样去完成年度工作。任何一个部门的预算用完时,仓库是不会满足任何要求的。
(1919,2,17)
今后我要澄清关于你订购预算之外的东西,以及订购没有拨款的东西的疑团……在有了将近二十年的采购经验后,我想你应该知道,我跟你一样无权花费不归我们支配的钱。
如果我通知你去订购(价值600美元的干燥器),那是我越权了。我决心不再发生这种情况。
(1923,6,19)
每个人都应遵守同样的规矩……设想在德国会是怎样的情况,那是徒劳的。
(1924,10,22)
遗憾的是,雇用的工人多于你的支付能力。
(1931,2,30)
1929年左右,弗莱克斯纳对于利文谈到的计划的规模深表关注。弗莱克斯纳写道,过去“你处理了一个(问题)然后处理另一个,当这些问题是有益的时候就继续搞下去,然后把它们放下去搞另一些问题”,现在他发现利文“一门心思”研究维生素达五年之久。“在这段时间里,你自然会发现是否值得对这个问题作出更大努力。”(弗莱克斯纳,1929)1929—1930年,使弗莱克斯纳担心的另一项花钱很多的工作是利文对胰脏核酸(班所说的鸟嘌呤核苷酸)的酶学研究,以及利文打算鉴别胸腺核酸中的糖。弗莱克斯纳指出,1926年到1930年间,利文那个部门一共花了将近34.3万美元!1929年利文的年薪为1.6万美元,而在1929年到1930年间,他的整个部门的费用增加1万美元。稍可告慰的是利文可告知弗莱克斯纳,他好不容易才成功地鉴定出胸腺核酸里的糖是2-脱氧核糖;他在1929年6月时写道:
……明年有希望弄清其余的细节。经费用光对我来说是可怕的——单是甜面包的账单就达1500美元,装备伦敦博士的操作室花了250美元。不包括伦敦博士的薪金在内,整个实验室的费用超过3000美元。
1929年,对于利文来说是个什么样的年头!他同来自斯德哥尔摩哈默斯坦实验室的乔帕斯合作,从有很多争论的胰脏核酸中得到碱基分析和核苷。1928年时乔帕斯曾称之为真正核酸的这种物质——最初是在动物组织中鉴别出来的——使利文得到了鸟嘌呤和腺嘌呤的核糖苷,使他确定胰脏核酸是一种多聚核糖核酸(或用现代术语来说,是一种寡聚核糖核酸)。
利文同来自列宁格勒的教授E.S.纶敦一起进行酶学研究,从而证明在胸腺核酸中的糖是脱氧核糖。1912年在纽约和1924年在列宁格勒,他曾有两次机会取得这项伟大成就,但都错过了。在这两次机会中,他研究了在狗的肠瘘管中核酸的生理性消化,希望在温和和控制的条件下使核酸水解,所用的试剂是1911年他同梅德格里西发现的核苷酸酶和核苷酶。1924年利文在巴甫洛夫实验室里无法使胸腺核酸降解,但纶敦来到洛克菲勒研究所四年后给几条狗做了胃瘘和肠瘘,这样利文就能让胸腺核酸通过胃而在肠里收集。这种方法只要用很短一段肠道,利文和纶敦不仅得到了两种嘌呤和两种嘧啶的核苷,还得到这些核苷的糖的组成成分。这样就弄清了这既不是以前普遍认为的六碳糖,也不是孚尔根称之为葡聚糖的那种糖,而是1895年基利安尼首次分离到的糖,他还为这种糖设计了“松枝”试验,以及用铁离子和硫酸进行的试验(基利安尼,1896)。
利文和纶敦无法得到这种糖的脎——在他们之前基利安尼也失败了——这是因为在C2位置上没有羟基。因此,他们以苄苯腺作分析。由此得到了脱氧核糖衍生物的经验公式。
利文也是在1929年同国际教育部的研究员T.莫里一起,比较了他曾称之为“胸腺糖”的胸腺核酸里的脱氧核糖同3种合成的2-脱氧核糖。这4种糖的希夫反应都是正反应,都被硫酸转化成乙酰丙酸;这样,在胸腺核酸降解产物中发现这种酸也就得到了说明。
孚尔根这样伟大的核酸化学家因为化学家们无法应用适合于酵母核酸的技术去从胸腺核酸中抽提核苷和核苷酸而感到沮丧,“表达了一种坚定信念,认为这种化合物(DNA)的构造完全不同于植物核酸”(利文,1931,264;孚尔根,1923,296),人们可看出这些成就的重要性。
利文分离核苷和核苷酸的早期成就使他享有盛誉,他还提出了无可辩驳的证据表明胸腺核酸和酵母核酸都是嘌呤和嘧啶通过糖-磷酸-酯骨架彼此连接成的直链,如图6.5所示。
如果利文和雅各布斯(1909c,2703)以及斯托伊德尔(1912)的磷酸-酯骨架过时了,那么利文和琼斯(1912)提出的这些结构的多糖骨架将不会生成四种核苷酸和核苷。这一成就的重要因素在于利文把酶学工具——这方面是依赖于巴甫洛夫实验室的技巧——同他拿手的降解产物的有机分析结合了起来。他熟谙的立体化学技术也起了无可估量的作用。像利文这样一位机敏的化学家想必一定会在他的酶制剂中发现解聚活性的证据,这样,他也一定已认识到天然的胸腺核酸是一种高聚物吗?是的,虽然应该说最后促使他抓住这种活性的早期发现深入下去的也许是《自然》上的一篇报告,这篇来自斯德哥尔摩的报告说相对分子质量在500000~1000000之间(西格纳、卡斯帕森和哈默斯坦,1938)。

图6.5 利文提出的RNA和DNA的四核苷酸的“酯”结构(录自利文,1919;1921)。
胸腺核酸的一种解聚酶
人们会想起利文虽然成功地从酵母核酸中分离出核苷和核甘酸,但1909年从胸腺核酸中分离时却失败了,这促使他继续对核苷酸感兴趣。1930年,R.T.狄龙和利文很惊奇地发现,如果他们用经过沉淀的肠道核酸酶制剂代替未经处理的制剂,则从胸腺核酸中释放出磷酸的速度有所不同,一个样品的释放速度只有另一个样品的十分之一,尽管它对磷酸甘油的活性很高。利文猜测在这份样品里缺少一种“核酸酶”(一种解聚酶),另一份样品中则有这种酶;因而设想核酸降解成核苷酸时必定经过脱磷酸化作用(利文和狄龙,1930,756)。八年后,利文和施米特表明,核苷酸酶(从核苷酸中释放出磷酸)纯度越高,胸腺核酸制备得越小心,脱磷酸化酶的活性就越低。利文在同事J.H.鲍尔和E.G.皮克尔斯的帮助下,用超离心方法表明用纽曼方法制备出来的核酸(α-胸腺核酸)的相对分子质量在五万到一二百万之间,而孚尔根的酶制剂则始终沉淀不下来。利文的结论是,纽曼型核酸是链长度不等的分子混合物,而孚尔根型物质则是已高度降解的。利文正确地强调了这一发现的重要性,他在《科学》上写道:
……这个发现很重要,不仅因为它揭示出核酸生物降解过程中的另一个步骤,而且在于它提供了一种方法,一方面测定“天然”核酸的纯度,另一方面利用天然核酸去测定核磷酸酶的纯度。
对于同核蛋白结构有关的其他问题来说,这也有重要意义……
(施米德和利文,1938,173)
最后,利文同胸腺核酸[1]的多聚a型及其降解产物b型的老概念取得一致;b型是纽曼于1899年报道,孚尔根在吉森比较了旋光性和凝胶生成后加以证实的(孚尔根,1914)。孚尔根进一步验证通过胰液(商品制剂)的作用a型转变成b型。因为这破坏了a型生成凝胶的活性,他称这种抽提物中的活性因子为“核胶酶”(孚尔根,1923,276;1935,237)。
这不是核酸解聚作用的唯一证据。在比斯德哥尔摩和吉森近得多的地方——在洛克菲勒研究所里——利文知道胰脏抽提物很容易使核酸解聚。这是伦敦药物学会研究员雷内·杜博斯和R.H.S.汤普森发现的。线索来自杜博斯和科林·麦克劳德的工作,他们发现高温杀死的肺炎球菌失去了刺激抗体生成的能力,而且这同细菌从革兰氏阳性变为革兰氏阴性相联系着。他们从许多组织里分离出一种抽提物,它造成染色性质的这种改变,在检测一些纯化合物对该抽提物的降解作用是否敏感时,只有利文提供的酵母核酸受到该抽提物的作用(杜博斯和麦克劳德,1937,697)。杜博斯把这种酶称为多聚核糖核苷酸酶(杜博斯,1937,550),并断定染色性质的改变同RNA的破坏有关。报道这一工作的详细论文,同利文和施米德关于脱氧核糖解聚酶(杜博斯和汤普森,1938)的论文同时发表。杜博斯会记得曾应邀在利文的办公室里共进午餐,所以那位伟大的核酸化学家可能知道杜博斯所发现的酶(杜博斯,1969)。一种发现往往在发表以前就有了报道,现在的情况就是如此。那就是沃尔特·琼斯曾报道胰液抽提液对酵母核酸有作用(琼斯,1920),但以后打算重复他的工作却失败了。
如果杜博斯、孚尔根、卡斯帕森、西格纳和利文都认识到天然状态的胸腺核酸就是现代所说的高聚物,那么四核苷酸又是什么?四核苷酸完整地保留下来,作为通过聚合作用生成天然核酸中的多聚物的基本单位。1938年,施米德和利文写道:“天然核酸是四核苷酸的一种聚合物。”(利文和施米德,1938,425)关于胸腺核酸,他们的评论是:“至今还未能用化学方法使(天然)核酸完全解聚成单个四核苷酸。”(施米德和利文,1938,173)很清楚,他们认为这是可能做到的。孚尔根确信:“引用的所有证据都谈到用核凝胶酶得到的b胸腺核酸仍显示出四核苷酸结构,而且酵素的作用是解聚……”(孚尔根,1935,266)最后,免得读者们认为孚尔根已得到支持这一结论的碱基比,有必要指出,他在1935年记录下来的a和b胸腺核酸的碱基量为0.5G: 0.9A:0.8C∶1.9T!毋需多说,孚尔根并未去研究这些分子比。他所关心的是要表明a型和b型的碱基成分是相同的。
1874年到1962年期间,DNA分子量的图表(图6.6)表明,1935年到1940年间估计数的变动何等之快。DNA和RNA现在成了一种大分子,但“中心法则的蛋白质版本”基本仍未动摇,因为如同古兰德所说,多聚四核苷酸“将限制可能出现的异构体的数目,从而减小生物学特异性的可能性”(古兰德,1947,12)。1944年,这位伟大的核酸化学家喜欢把多聚四核苷酸“作为一种实用的工作假说,如果充分认识到它的局限性,将有助于解释实验结果”(古兰德,1944,216)。他说这个假说还没有确凿证据,一年后,他感到有必要引进“统计上的四核苷酸”这一术语用以描述这样一种多核苷酸:它的碱基组成等于四核苷酸,但碱基序列是“随机的”(古兰德、巴克和乔丹,1945)。1946年,当他在实验生物学学会组织的核酸讨论会上作报告时,他认为他的假定数据可以得到解释,但同DNA中碱基等分子量假设相抵触。

图6.6 从1874年到1973年间,DNA分子量的估计值(T.H.=四核苷酸的数值)。
我的结论已被证明是正确的,即除了细菌转化研究之外(见第十二章),导致认识大分子并进而注意到化学序列的化学中的革命,总有一天会破坏中心法则的蛋白质版本。但不是让化学家单枪匹马去完成这一过程。细胞化学家起了决定性作用,而且我们在下一章里将看到,他们的研究结果只是部分地认识了DNA的生物学重要性,直到艾弗里的工作促使他们重新评估自己的资料以前,还一直在保护中心法则的蛋白质版本不被完全推翻。
【注释】
[1] 利文和巴斯:1931年第289页中,对DNA可能是一种多聚状态持勉强同意的态度。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










