



第四节 细菌的遗传与变异
细菌的遗传是指亲代与子代细菌的相似性,它使细菌的性状保持相对稳定,且代代相传,是物种存在的依据。细菌的变异是指亲代与子代以及子代细菌之间存在的性状上的差异性,它是细菌得以进化的依据。
一、细菌的遗传物质
细菌的遗传物质包括细菌的基因组和基因组以外的遗传物质。一般细菌的基因组控制细菌的主要性状,而基因组以外的遗传物质控制着细菌的次要性状。
(一)基因组
细菌基因组(genome)位于拟核区,称为拟核,又称为细菌染色体,是遗传的主要物质基础。细菌基因组由两条共价闭合的、环状双股DNA组成,控制细菌的遗传与变异。DNA呈超螺旋状态存在,基因组中约有1%的碱基被甲基化;细菌的基因是连续的,无内含子,且功能相关的基因高度集中。细菌基因组以半保留方式进行复制,新形成的DNA双股分子所携带的遗传信息与亲代的完全相同,故子代与亲代细菌的性状相同。
(二)质粒
质粒(plasmid)为细菌基因组外的遗传物质,绝大多数细菌质粒为闭合环状双股DNA分子。质粒是自主复制单位,编码产物赋予细菌某些非必需的生物学特性,随分裂传给子代。细菌质粒带有遗传信息,可决定细菌的一些独特的生物学特性。质粒具有以下特性:
1.非细菌生存所必需:细菌失去质粒后仍能生存,但失去基因组则不能生存。
2.具有独立复制能力:质粒的复制可不依赖于基因组,而在细胞质内进行。
3.可自行消失或人为消除:细菌在培养和传代过程中,质粒可自然丢失,也可在高温、紫外线、诱变剂等作用下丢失。丢失的质粒可使宿主相关的表型也消失。
4.可以传递或转移:质粒可通过接合、转化或转导等方式进行转移。获得质粒的细菌可同时获得质粒所携带的相应生物学特性,如毒力、细菌素、耐药性等。
5.具有相容性与不相容性:因质粒可独立复制,又能转移和自然丢失,因此宿主有机会出现几种质粒共存的现象,此为质粒的相容性;但在没有选择压力的条件下,两种结构相似、密切相关的质粒不能稳定共存于同一宿主菌的现象称为质粒的不相容性。了解质粒的不相容性,就有可能把功能不同的基因分别克隆到两个相容的质粒上,转入同一宿主细胞内,研究基因在体内的表达情况。
质粒具有与DNA重组的功能,所以在基因工程中被广泛用作载体,为遗传工程的应用开辟了广阔的前景。
(三)其他遗传物质
细菌中还存在一类独特的DNA片段,不依赖于同源重组,可以在细菌的基因组中从一个位置转移到另一位置,称为转位因子。转位因子插入细菌基因组后,因在插入部位影响了细菌基因组DNA的正常结构,可使细菌失去某些功能,如致病性、耐药性等。
此外,细菌的遗传物质还包括基因盒-整合子系统、毒力岛等。基因盒是较小的、可移动的DNA分子,常以环状独立状态存在,只有被整合子捕获并整合到整合子中才能转录。基因盒-整合子可整合到转座子内,能整合不同的耐药基因盒,且一个整合子可捕获多个基因盒,因此可表达出对抗药菌的单一甚至多重耐药性。毒力岛除含有编码细菌毒力的基因簇外,一些毒力岛还编码细菌的分泌系统、信号传导系统和调节系统等。
二、细菌的变异
细菌可以自发地或在人为条件下发生变异。在细菌生长繁殖过程中变异现象常常出现。
(一)细菌变异的类型
按细菌变异的表现形式可分为以下几种。
1.形态结构的变异:细菌的大小和形态在生长过程中可受外界环境的影响而发生变异。例如,细菌在抗生素作用下可出现细胞壁缺陷的L型;有的细菌可失去荚膜、芽胞、鞭毛(H-O变异)等特殊结构。
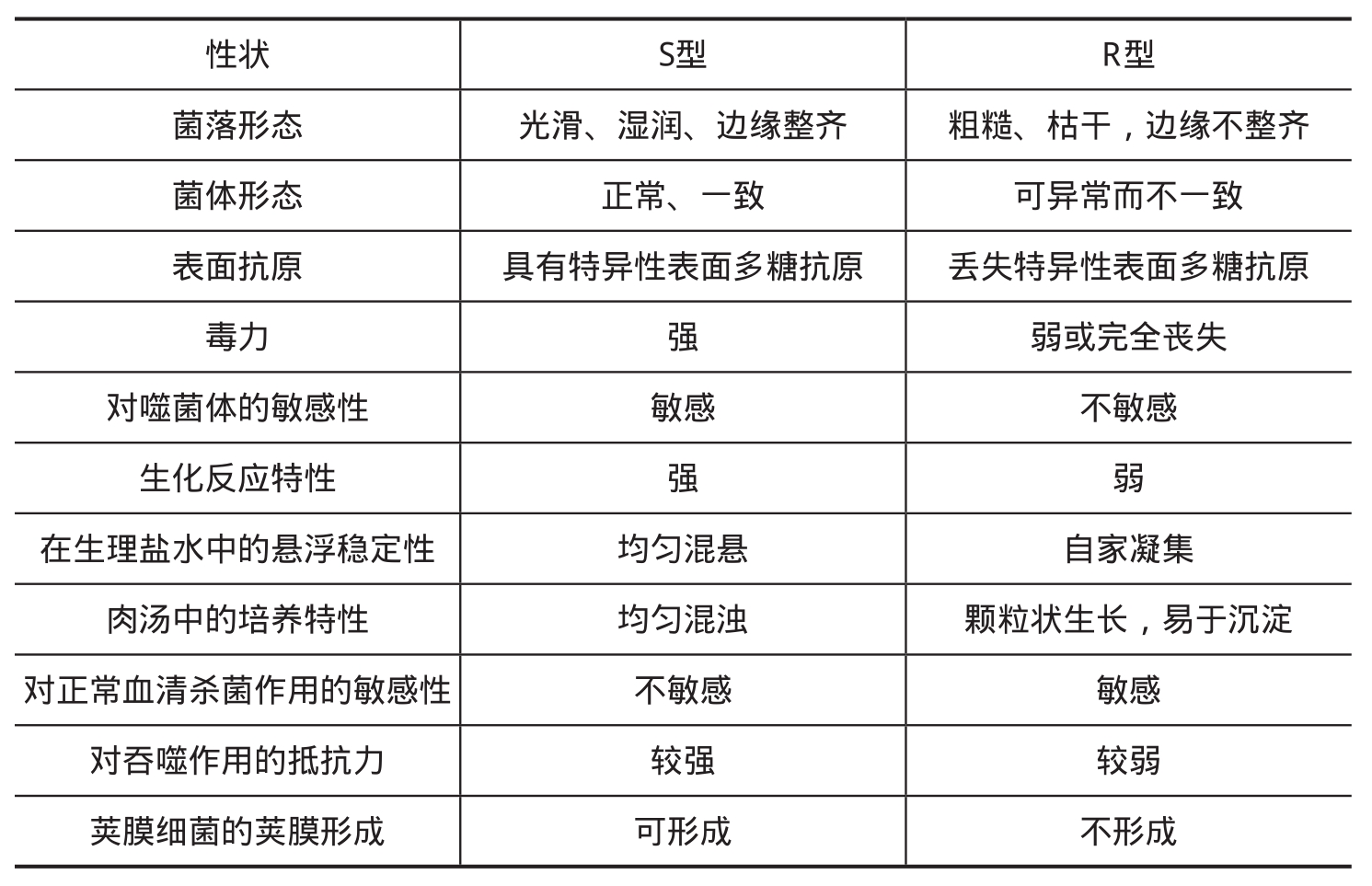
2.菌落变异:细菌的菌落主要有光滑型(S型)和粗糙型(R型)两种类型。S型菌落表面光滑、湿润、边缘整齐;R型菌落表面粗糙、枯干,边缘不整齐。绝大多数新分离的菌株为S型。在一定条件下(如长期人工培养或培养于含有特异抗血清的培养基中等)S型菌落可变为R型菌落,称为S→R变异。S→R变异常见于肠道杆菌,这种变异经常伴随着一系列其他性状的改变,包括表面S型抗原的丧失和病原菌毒力由强变弱等。也有极少数细菌(如炭疽杆菌、结核分支杆菌),其新分离的菌落正常为R型,在一定条件下则变为S型。光滑型与粗糙型菌落的比较见表2-3。
表2-3 S型与R型菌落的性状比较
3.抗原变异:当编码细菌的基因突变时,就会引起抗原结构发生突变。如菌体抗原、鞭毛抗原、荚膜抗原等的基因发生突变时,细菌形成相应抗原结构的能力丧失,引起细菌抗原性变异,从而不能被特异的抗体所凝集。肠道杆菌中如沙门菌属、志贺菌属中常发生鞭毛抗原以及菌体抗原的变异。
4.毒力变异:病原菌的毒力可以由弱变强或由强变弱,在自然情况和人工诱变下均可以发生。连续通过易感动物可使病原菌毒力增强,而通过非易感动物可使细菌毒力减弱。
5.耐药性变异:指细菌对某种抗菌药物由敏感变为耐药的变异。由于抗生素的广泛应用,细菌对抗生素耐药性越来越普遍,有些细菌还表现为多重耐药。细菌的耐药性变异给临床治疗带来很大的困难,并成为当今医学上的重要问题。
6.代谢变异:细菌代谢变异的现象较为常见的有以下两种。
(1)营养型变异:即某一菌株由于发生基因突变而丧失合成一种或几种生长因子的能力,因而无法在基本培养基上正常生长繁殖的变异类型。营养型变异菌株可作为杂交、转化、转导和原生质融合等研究中的标记菌种。如变异株丧失对某种糖类、维生素、氨基酸或其他生长因子的合成能力,在补充这些营养物质的培养基上才能生长。这种突变对研究细菌代谢产物的生物合成途径很有用处。
(2)终末产物阻遏:是指细菌合成氨基酸等的酶类,可被自身的合成产物所抑制而不能生成的变化。例如,大肠杆菌可生成色氨酸合成酶类以合成色氨酸,但若于培养基中加入色氨酸时,则其色氨酸合成酶类的生成即被抑制。
7.条件致死突变型:在某种条件下,某菌株经基因突变后可正常生长、繁殖并实现其表型,而在另一条件下却无法生长、繁殖的突变类型。温度敏感突变株(Ts突变株)是一类典型的条件致死突变株。产生Ts突变的原因是突变引起了某些重要蛋白质的结构和功能改变,以致在某特定的温度下能发挥其功能,而在另一温度(一般为较高温度)下则该功能丧失。例如,大肠杆菌的某些菌株可在37℃下正常生长,却不能在42℃下生长等。
(二)细菌变异的机制
1.基因突变:突变是细菌遗传物质的结构发生突然而稳定的改变,导致细菌性状的遗传性变异。基因突变的原因是多种多样的,可以是自发的或诱发的。由于自然界中诱变剂的作用或偶然的复制、转录、修复时的碱基配对错误所产生的突变称为自发突变。自发突变的突变率极低,一般在10-6~10-9范围内,且突变是随机的;由人工利用物理因素或化学药剂诱发的突变称为诱发突变。诱发突变可大大提高突变率。在生产上,人工诱变是产生生物新品种的重要方法。基因突变是细菌变异的主要原因,也是生物进化的主要因素。
2.基因的转移与重组:供体细菌可直接或间接地将部分遗传物质单向传递给受体细菌,从而导致受体细菌发生基因重组。细菌基因转移与重组的主要形式有转化、转导、接合和原生质体融合。
(1)转化:转化是指受体菌直接摄取供体菌的游离DNA片段,通过与基因组重组获得供体菌的部分遗传特性。转化的发生过程,首先是供体菌的DNA片段吸附于处于感受态的受体菌的细胞膜上。细胞上的DNA与一种特异的可以抵御核酸酶作用的蛋白结合形成复合物,然后穿入受体菌细胞内,与其DNA发生整合,受体菌由于获得外源的DNA而改变遗传性状。转化在自然和人工条件下均可发生,而且无论G-菌还是G+菌都有发生。但并不是所有细菌都有转化现象,大多数细菌不能将外源DNA整合到基因组中去。细菌还能通过内切酶识别并破坏进入细胞的外源DNA。能否发生转化与菌株的亲缘关系有关,亲缘关系越近的菌株越容易发生转化。同时,受体菌必须处于感受态时才能转化。
转化现象首先由Griffi th在肺炎球菌中被发现(图2-14)。此后,在许多细菌中都发现了转化现象。
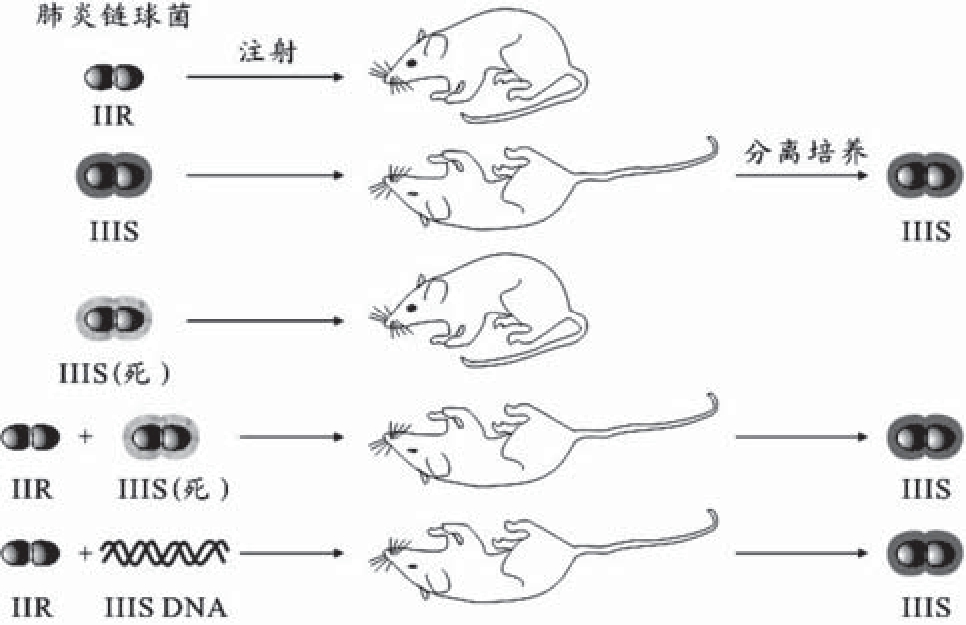
图2-14 Griffith的肺炎球菌的小白鼠实验
(2)转导:转导是以温和噬菌体为媒介,将供体菌的一段DNA转移到受体菌内,通过交换与整合,使受体菌获得供体菌的部分遗传性状。根据转导基因片段的范围,可将转导分为普遍性转导和局限性转导两类。普遍性转导是指温和噬菌体在裂解期的后期,在装配成新的噬菌体时,部分噬菌体误将细菌DNA的裂解片段装入其外壳蛋白中。错误装入的DNA片段可以是供体菌的染色体或质粒的任何DNA片段,所以称为普遍性转导(彩图7)。普遍性转导的结果有两种:若供体菌DNA片段与受体菌的染色体DNA重组,并与染色体一起复制成为稳定的转导子,称完全转导;若供体菌的DNA片段不能重组到受体菌的染色体上,那么由于它本身不具有独立复制功能,随着细胞分裂,供体DNA片段并不能遗传给后代,称流产转导。局限性转导是噬菌体所介导的供体菌染色体上特定基因的转导(彩图8),为前噬菌体从宿主菌染色体切离时发生偏差交换而形成的。
(3)接合:是指两个完整的细菌细胞通过性菌毛相互连接沟通,由供体菌将遗传物质(主要是质粒DNA)转移给受体菌的过程。能通过接合方式转移的质粒主要有F质粒、R质粒、Col质粒等。染色体基因也可被某些质粒诱导而转移给受体细胞。迄今为止已发现的质粒接合传递体系中,F质粒研究的较为详细。F质粒编码性菌毛,首先在大肠杆菌中被发现。染色体外有F质粒存在的细菌称为雄性菌(F+),接合时作为供体菌。无F质粒的为雌性菌(F-),接合时作为受体菌。将F+与F-细菌混合培养,由于性菌毛的作用,F+菌与F-菌接触,F+菌的性菌毛末端可与F-菌表面上的受体结合,结合后性菌毛渐渐缩短,使两者紧靠在一起,F+菌中的F质粒的一股DNA链断开,逐渐由细胞连接处伸入F-菌,继而以滚环模式进行复制。接合过程结束,两个细菌细胞内均形成一个双股F质粒,F-变成F+,也长出性菌毛(图2-15)。另外,R质粒控制细菌耐药性的形成,Col质粒控制细菌素的产生,均可通过接合方式进行细菌间的转移。
(4)原生质体融合:原生质体融合是指选择具有优良性状的两个菌株细胞作为亲本,用人工方法去除其细胞壁,使之成为原生质体,然后在高渗条件下混合,并加入物理的、化学的或生物的助融条件,使原生质体间发生相互融合,通过细胞核融合而发生基因组间的交换、重组,在适宜条件下再生出细胞壁可获得重组子。在重组子中可获得带有双亲优良性状且遗传稳定的融合子。原生质体融合除可在不同菌株间或种间进行外,还能在属间、科间甚至更远缘的细菌细胞间融合。
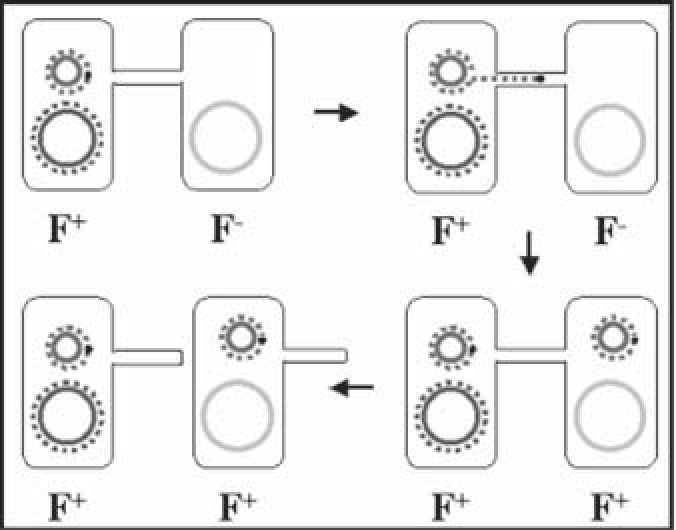
图2-15 F质粒在细菌间的接合转移
三、研究细菌遗传与变异的意义
研究细菌的遗传与变异不仅揭示了细菌本身许多遗传变异的规律,而且推动了整个分子遗传学的迅速发展。在实践方面,细菌遗传与变异在微生物诊断及疾病预防等方面具有重要意义。
1.在疾病诊断、治疗、预防中的应用:由于细菌形态、结构、染色特性、生化特性、抗原性及毒力等方面的变异,使得临床细菌学诊断更加复杂。例如,猪丹毒杆菌在慢性病猪的心脏病变内呈长丝状;伤寒沙门菌有10%不产生鞭毛,检查无动力,无H抗原;血浆凝固酶阴性的葡萄球菌也为致病菌。这提醒我们在对临床传染病进行诊断时要注意了解细菌的变异规律,避免在临床分离菌的鉴定与疾病诊断中造成误诊。
临床耐药菌株,尤其是多重耐药菌株的日益增多,给疾病的治疗带来很大困难。因此,对临床分离的致病菌株必须进行耐药性监测,通过药敏实验正确选择药物,不能滥用抗生素。
用遗传变异的原理诱变强毒株成为减毒株和无毒株可制备预防疾病的各种疫苗,这是预防传染病的有效措施。目前疫苗的研究,更应该采用遗传变异的原理,通过基因的转移和重组或基因的突变获得的减毒活疫苗是理想的预防接种制剂。
2.在测定致癌物质中的应用:凡能诱导细菌发生变异的物质都有可能具有致癌性。例如,通过Ames试验(亦称沙门菌回复突变试验)可以检测污染物的遗传毒性效应。
3.在流行病学中的应用:随着分子生物学的发展,细菌分子流行病学研究越来越受到重视。通过对引起某一细菌性传染病流行的病原菌在核酸水平上进行分析,从而更准确地解决传染源和传播途径及其他有关的流行病学问题。如质粒指纹图谱的分析方法可用于调查实际生产中所感染的各种细菌的某种耐药质粒的传播扩散情况。
4.在基因工程中的应用:基因工程技术可通过基因转移和重组而使细菌获得新的性状。将能编码结构性抗原表位基因结合在质粒或噬菌体载体上,通过载体将目的基因导入受体菌中,制备基因工程疫苗或研制DNA疫苗等生物制品,为提供更加高效、无毒副作用的免疫制剂创造了理想的途径。此外,还可以利用基因工程技术大量生产胰岛素、干扰素、多种生长激素等细胞因子。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











