



二、微生物的趋化性
微生物的趋化性是细胞趋向吸附剂化合物或离开排斥剂化合物的行为。实际上每一种运动生物都展现出某些类型的趋化性。趋化性是微生物趋向有利环境,避开不利环境的行为,对微生物在生态环境中的分布有重要意义。大肠杆菌的趋化性被深入研究,这里介绍的基本都是以大肠杆菌为材料的研究成果。
1.趋化性行为
在一个稳定的环境中,细菌的运动是随机的,其在一个方向作直线运动时会不时随机改变运动方向,一般是向前直线运动几秒钟就停下来翻筋斗,然后又以不同方向做直线运动,循环往复。趋化性行为是这种随机运动的一种偏离,是环境信号对细菌细胞在运动方向上随机改变的一种调整(图1-12(a))。加入吸附剂使细胞连续做直线运动而减少改变运动方向,平均的直线运动距离要长于随机运动时直线运动的距离。这种情况一直到细胞适应于吸附剂,运动又回到原先的随机行为;移去吸附剂则使细胞频繁改变方向(翻筋斗)而更少直线运动,直至适应新的情况,又回复到随机行为。加入排斥剂会产生和移去吸附剂同样的应答情况,而移去排斥剂和加入吸附剂可以有同样的结果。
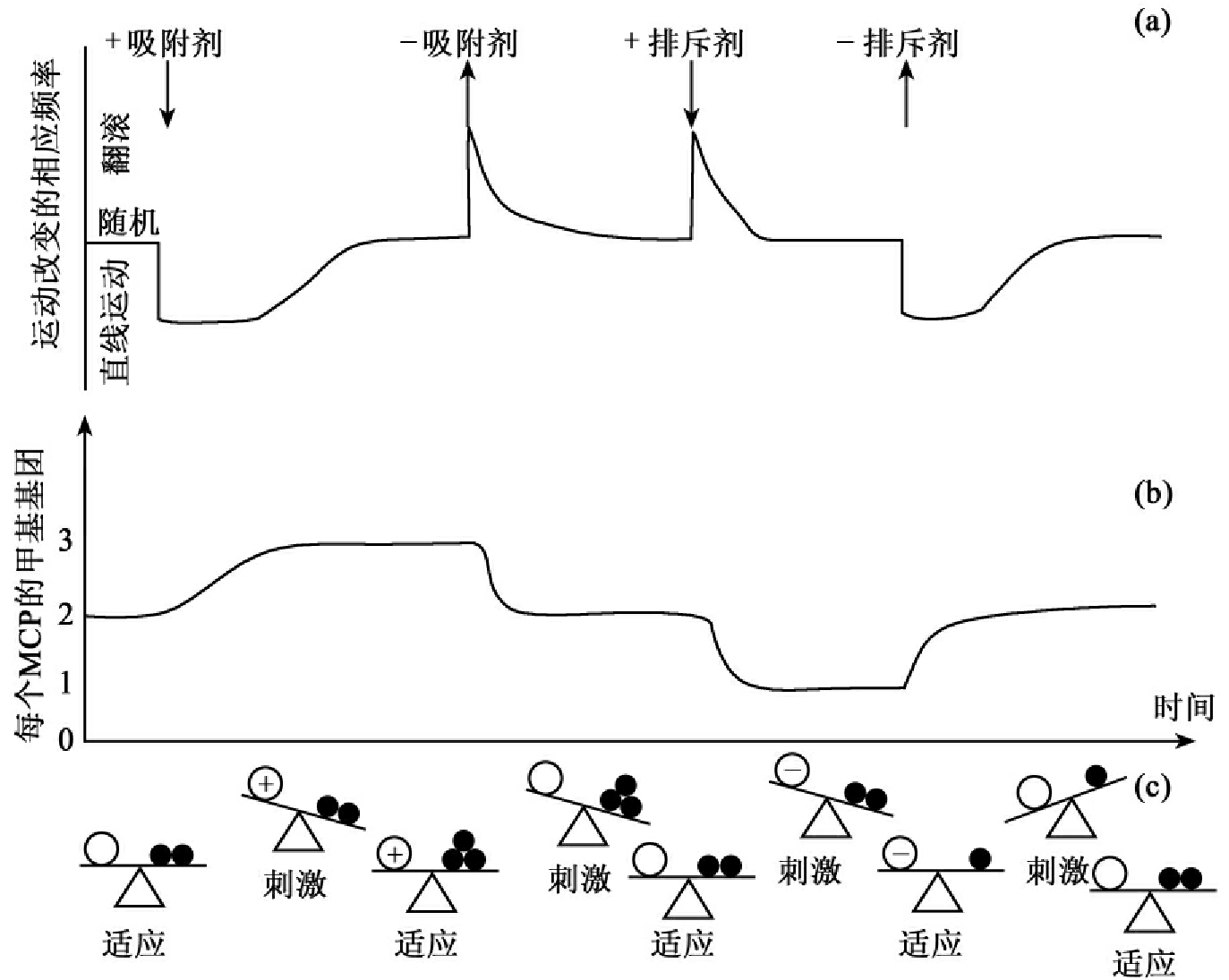
图1-12 细菌趋化性的刺激与适应
(a)加入吸附剂使细胞连续做直接运动而减少改变运动方向,直到细胞适应于吸附剂,运动又回到原先的随机行为;移去吸附剂则使细胞频繁改变方向(翻筋斗),而减少直线运动,直至适应又回复到随机行为。加入排斥剂会产生和移去吸附剂同样的应答情况,而移去排斥剂和加入吸收剂有同样的结果。
(b)在没有吸附剂或排斥剂时,一个典型的MCP中4个可能甲基化的谷氨酸中有约2个被甲基化,加入一种饱和的吸附剂刺激可增加大约一个甲基基团,移去吸附剂则会失去这个基团,加入饱和的排斥剂刺激会失去一个或一个以上的甲基基团,而移去排斥剂会增加一个甲基基团。
(c)MCP信号取决于变动(○,没有变动;+,吸附剂;-,排斥剂)和甲基化水平(○,一个甲基基团;●●两个甲基基团等)之间的平衡。一个突然产生的变动会导致失衡而产生一种刺激,而后甲基化水平的改变又会回复到平衡。
细菌的趋化性行为与细菌对环境条件的适应、记忆以及环境条件改变时的感觉、刺激有关。如细菌种群处于一种稳定环境条件下,它们会逐步适应,并在适应过程中就会产生对这种条件的记忆,适应了环境的运动细菌其运动都是随机的。当向环境投入吸附剂、排斥剂时,细菌就会感知浓度变化,现在的感知和过去的记忆的比较就会产生一种刺激,正是这种刺激改变细菌随机运动的轨迹,产生一种趋化性行为。实际上细菌的这种趋化性行为(因吸附剂、排斥剂引发的)可以看成是一种对环境条件改变的应答。把适应于吸附剂环境的细菌种群(随机运动)移到缺乏吸附剂环境,细胞会以为它们在错误的方向上运动而改变行为,这和暴露在排斥剂下的行为是同样的。当细胞已适应于排斥剂时,相反的情况也会发生。常见的吸附剂是微生物的营养物,如核糖、葡萄糖、半乳糖、麦芽糖、天冬氨酸、丝氨酸、谷氨酸、丙氨酸、甘氨酸,而排斥物则是一些对微生物的有毒物质,如重金属镍和钴等。
2.趋化性行为的生理遗传调控过程
趋化性行为的生理遗传调控过程是通过一个十分复杂的调控系统实现的。概括地说是吸附剂或排斥剂的环境信号首先被细胞膜上的跨膜受体蛋白接收,信号再通过信号传导系统的蛋白最终传导到控制细菌细胞运动的鞭毛传动器上,控制细菌的运动。
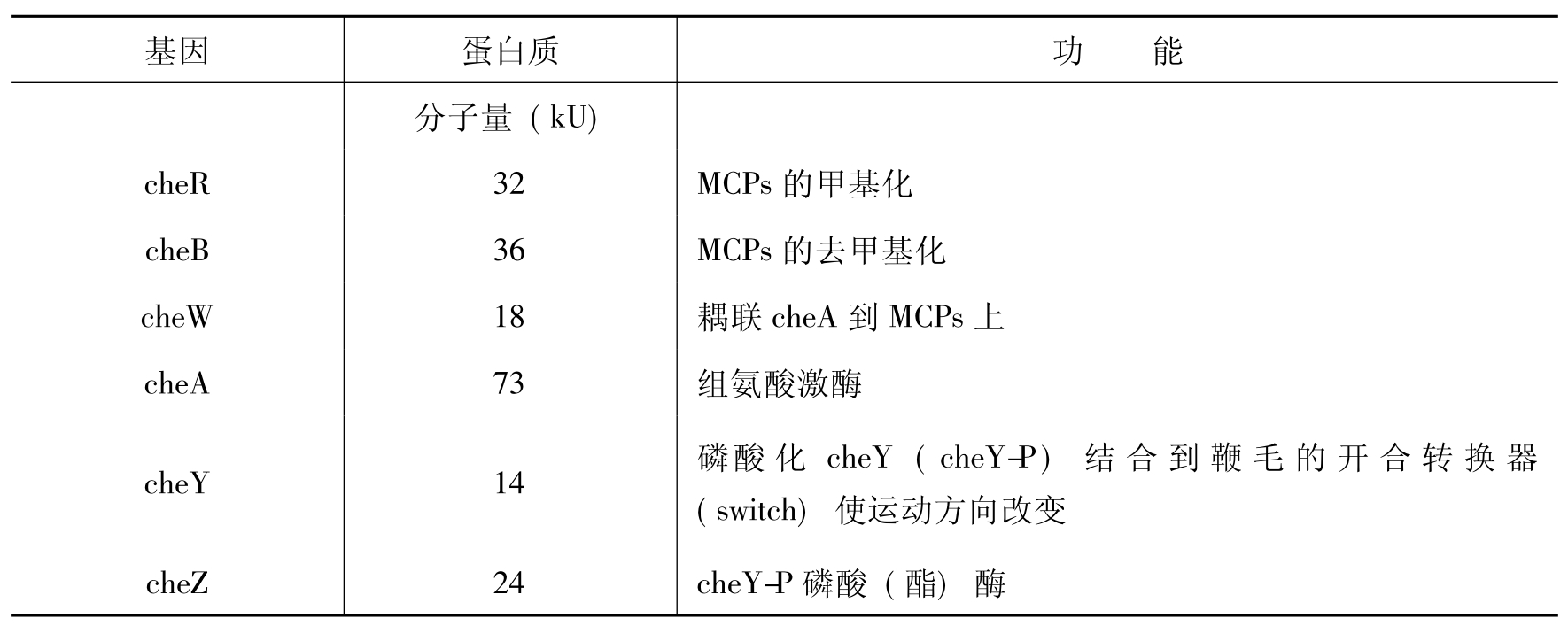
对E.coli趋化突变体(che mutants)的研究证明,在大肠杆菌中有两个主要的che互补基因座(comple mentation group),分别为cheA和cheB。后来的研究证明cheA由二个基因cheA和cheW组成;cheB由4个基因cheR、cheB、cheY和cheZ组成。缺陷cheA、cheW、cheY或cheR的菌株表现出直线运动的表型,很少改变它们的运动方向,相反cheB和cheZ突变体总是翻滚式改变方向。从这些研究结果可见cheA、cheW和cheY蛋白是应答刺激所必需的,而cheR、cheB和cheZ则与适应有关。che基因组和它们的蛋白质产物功能见表1-12。
表1-12 E.coli che基因组及其产物
受体蛋白是跨膜的接受甲基趋化性蛋白(methylaccepting chemotaxis proteins MCPs),其具有对吸附剂、排斥剂的专一性。这些蛋白是Tar、Tsr、Trg和Tap基因的产物,也称为MCP受体或Tar、Tsr、Trg和Tap受体。它们可以直接刺激配位体或通过周质结合天冬氨酸、谷氨酸、排斥剂(如镍、钴),还可以通过麦芽糖结合蛋白(MBP,maltose binding protein)与麦芽糖结合。已知的大肠杆菌的趋化性受体蛋白如图1-13所示。Tar和Tsr在细胞中的水平比Trg、Tap高10倍,因此Tar和Tsr被称为主要受体,而Trg、Tap为稀有受体。
受体MCP同时有感觉和信号结构,具有N-末端的胞质外感觉区,通过一个疏水性跨膜序列连接胞内的信号区。感觉区具有专一性,其专一性等同于MCP受体的专一性,这是由N-末端决定的。对Tar MCP感觉区的X射线结晶结构分析证明感觉区有两个α-螺旋束(α-helical bundles)的二聚体,当存在天冬氨酸时,天冬氨酸可以结合到二聚体的亚单位的表面。信号区也是α螺旋占优势,有高度保守的中心区,这个区域的α螺旋两侧有4个或更多的谷氨酸甲基化和去甲基化结合位点。中心区结合cheW、cheA,从而把受体和趋化信号系统连接起来。
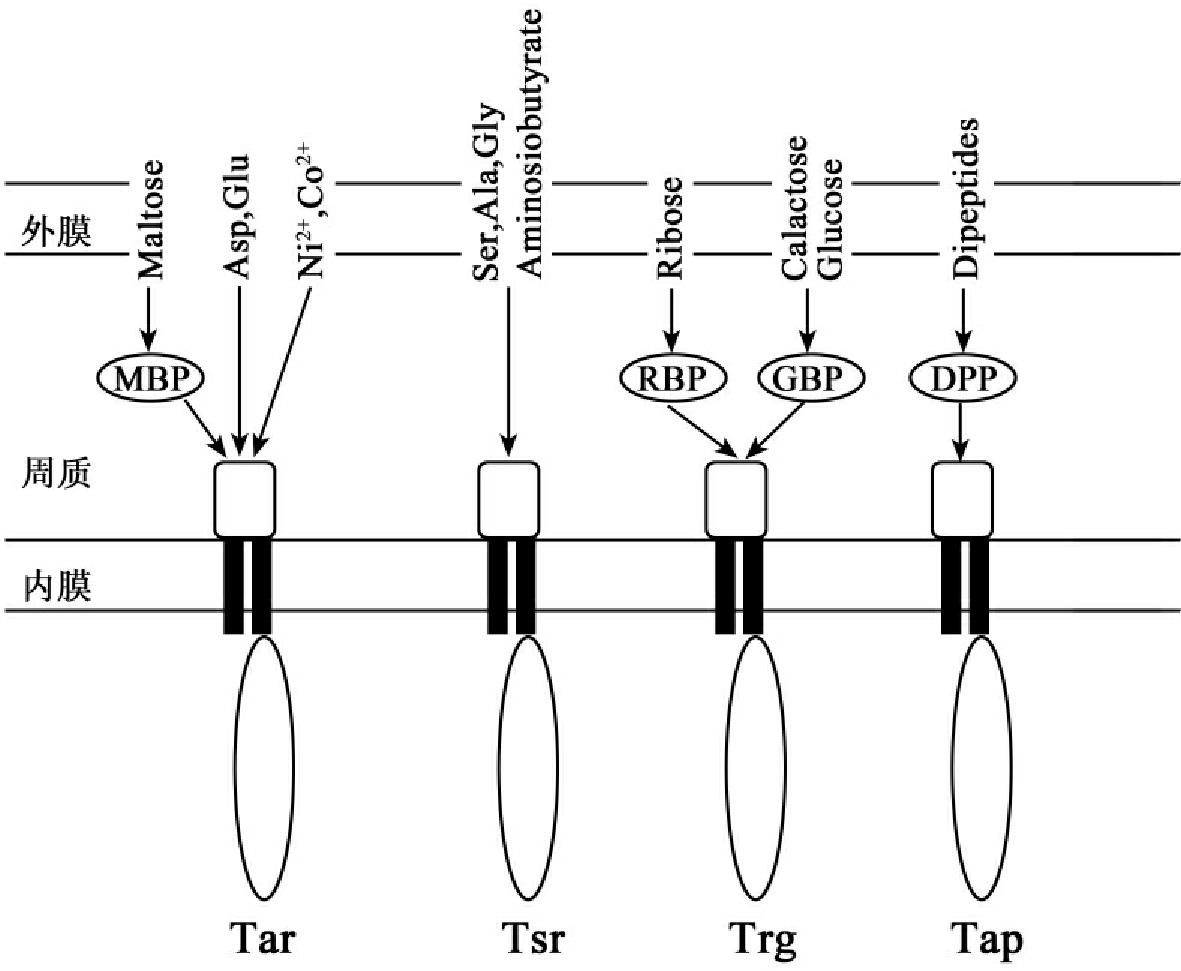
图1-13大肠杆菌趋化性受体。四种MCP受体:Tar、Tsr(taxis to serine and repellent)、Trg(taxis to Ribose and galactose)、Tap(taxis to dipeptide)。RBP:核糖结合蛋白(ribose binding protein)。GBP:半乳糖结合蛋白(galactose binding protein)。DPP:二肽结合蛋白(dipeptide binding protein)。
细菌的趋化性与MCP的甲基化有密切关系。细菌的趋化性需要甲硫氨酸,需甲硫氨酸生长的E.coli突变株对吸附剂的趋化反应受到抑制,失去趋化行为能力。甲硫氨酸的作用是变成S-腺苷甲硫氨酸,并为趋化性受体提供甲基。
![]()
吸附剂使甲基水平增加,排斥物使甲基化减少,这种改变是对吸附剂物和排斥物刺激的适应(图1-12(b))。
MCP的甲基化反应受依赖于AdoMet的甲基转移酶(cheR基因编码)的调节,同时也可被特异性的甲基酯酶(cheB基因编码)移去,CheR和CheB是两个溶解性单体蛋白。CheR通过与Tar、Tsr的C-末端4个氨基酸结合而连接在一起。CheR可以直接使主要受体直接甲基化,而使稀有受体甲基化的反应则更为复杂,酶被分泌到一定位置间接使受体甲基化。CheB的活性位点有一个丝氨酸-组氨酸-天冬氨酸催化三联体,催化丝氨酸的水解。一个N-末端调控区可阻断(occlude)活性位点使相应的酶失活。细菌应答排斥剂的加入会产生一个信号,这种信号使CheB主动消除受体的甲基基团,使甲基基团减少,又使细胞适应了排斥剂而进入一个适应期,也造成细胞运动方向的改变。排斥剂的加入和吸附剂的移去降低甲基化,吸附剂的加入或移去排斥剂又提高甲基化水平。受体蛋白的信号系统功能是使刺激配位体和甲基化之间达到一种平衡(图1-12(c))。加入吸附剂或排斥剂所产生的正的或负的刺激信号打破平衡,而由于有效适应造成的甲基化改变又回复到平衡。
控制细胞趋化行为的信号是由受体CheW和CheA组成的趋化信号复合物发出的,通过一系列磷酸化和去磷酸化得以实现。CheA蛋白是一种激酶,这种激酶和ATP结合,并催化它自身的组氨酸残基中一个残基的磷酸化。CheA蛋白的自磷酸化速率受甲基化水平及与刺激配位体结合情况的调控,最高可以提高100倍,也可完全被抑制,但一般是非常慢的。因此这个信号复合物的实际功能是调控激酶以及磷酸化的活性,最终的结果是对结合刺激配位体和受体甲基化水平改变的应答。
CheY蛋白的磷酸化水平可以影响细胞的运动行为。CheY是一个14kU的单体酶,其催化作用是把CheA的磷酸组氨酸的磷酸基团转移到它自己的天冬氨酸的残基上。CheY磷酸化诱导蛋白质的构象改变,改变构象的蛋白质被结合到鞭毛运动源(鞭毛传动器)的开关蛋白上。吸附剂的加入(如色氨酸或天冬氨酸)能抑制CheA的磷酸化活性。结合到运动源的CheY-P水平降低,细菌细胞做直线运动。排斥剂诱导造成CheA磷酸化产生能提高CheY磷酸化水平,磷酸化-CheY结合到运动源使细胞做翻滚(翻筋斗)运动。磷酸化-CheY自发的去磷酸化和CheZ对CheY自发磷酸化反应(去磷酸化)的增强作用可以降低CheY的磷酸化水平。此外在趋化行为中还有一种反馈调节功能。像CheY一样CheB也能把CheA的磷酸组氨酸的磷酸基团转移到自身的天冬氨酸残基上。CheB的磷酸化可使受体去甲基化活性戏剧性增加,使受体甲基化和CheA自磷酸化速率同时降低,就造成了一种反馈调节。低CheA磷酸化(加入吸附剂)使CheB磷酸化活性降低,反过来又使受体甲基化活性提高,这导致CheA磷酸化水平提高,这就是趋化行为中的适应,细菌细胞又从直线运动回复到随机运动。同样高CheA磷酸化(加入排斥剂)使CheB磷酸化活性提高,反过来又使受体甲基化活性降低,又导致CheA磷酸化水平降低,趋化行为又进入适应阶段,细菌细胞又从翻滚运动回复到随机运动。细菌(E.coli)的趋化性的调控系统可以概括为图1-14。
目前细菌趋化性的研究成果主要源于大肠杆菌,其他细菌趋化性研究也在不断进行,研究结果说明细菌的趋化性有种系的差异性,值得进一步研究。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











