



第一节 重组DNA技术中所需的工具酶
基因克隆的核心内容是对基因进行人工切割、连接组合,构建DNA重组体。在这些操作中,工具酶(tool enzyme)有着广泛的用途,特别是限制性内切酶和DNA连接酶的发现与应用,才真正使DNA分子的体外切割与连接成为可能。
一、限制性核酸内切酶
限制性核酸内切酶(restriction endonuclease)简称限制性内切酶,是一类能识别双链DNA分子中特定核苷酸序列并将其切断的核酸水解酶。主要是从原核生物中分离纯化出来的。
细菌中存在限制(restriction)与修饰(modification)现象,也称为寄主控制的限制与修饰现象,简称R/M体系。这是细菌的一种自我保护机制,其功能类似于高等动物的免疫系统,它能识别自己的DNA和外来的DNA,并使后者降解掉。在具有限制与修饰机制的细菌中,含有对某一DNA序列位点专一的核酸内切酶和修饰酶(通常为甲基化酶),由于甲基化作用,细菌自身DNA的限制性内切酶识别位点被修饰而免于降解,而外源的DNA上的内切酶识别位点未被修饰,因而进入宿主菌后即被降解。
1968年,Meselson和Lim等首先分别从大肠埃希菌(E.coli)B株和K株中分别获得了EcoK和EcoB两种限制性内切酶,到目前为止,人们已经从300多种不同的微生物中分离出了2000多种限制性内切酶。这些酶已经成为重组DNA技术的重要工具。
(一)限制性内切酶的命名
目前对限制性内切酶通用的命名系统是1973年H O.Smith和D.Nathans提出的,其命名原则包括:
(1)限制酶的第一个字母(大写、斜体)代表分离出该酶的微生物(宿主菌)属名(genus)。
(2)第二、三个字母(小写、斜体)代表微生物种名(species)。
(3)第四个字母代表宿主菌的株或型(strain)。
(4)最后的罗马数字表示从一株菌株中发现和分离限制酶的顺序。
如EcoRⅠ:E=Escherichia属
co=coli种
R=RY13株
Ⅰ=该菌株第一个被发现的核酸内切酶
(二)限制性内切酶的分类
根据限制性内切酶的识别切割特性、催化条件及是否具有修饰酶活性,分为3种类型,即Ⅰ、Ⅱ和Ⅲ型酶。3种不同的限制性内切酶具有不同的特性(表3-1)。在基因工程中普遍使用的是Ⅱ型限制性内切酶。
表3-1 限制性核酸内切酶的类型和主要特征
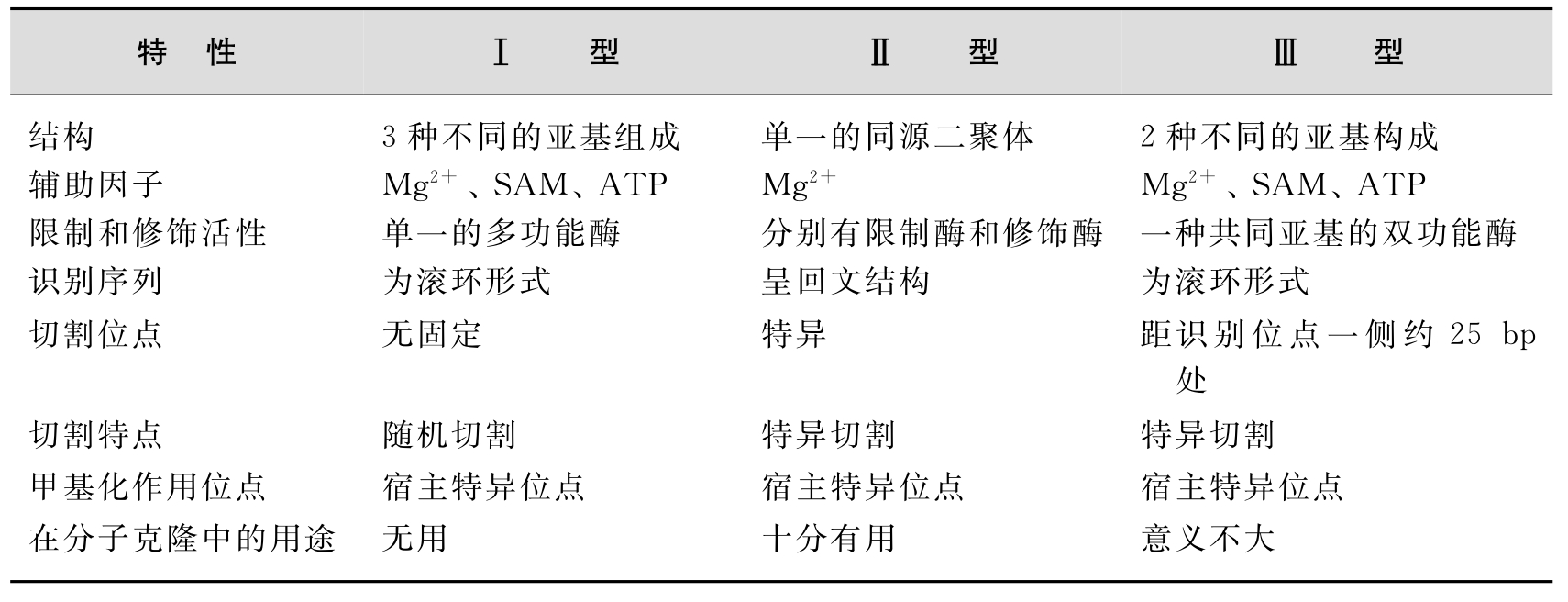
Ⅰ型限制性内切酶:是一类兼有限制性内切酶和修饰酶活性的多个亚基的蛋白复合体。既有限制性内切酶的作用,也有甲基化酶的作用。它们在离识别位点至少1000bp的地方切割DNA链。以前人们认为Ⅰ型限制性内切酶很稀有,但现在通过对基因组测序的结果发现这一类酶其实很常见。如EcoR和EcoK。
Ⅱ型限制性内切酶:由一群性状和来源都不尽相同的蛋白组成,在其识别位点之中或临近的确定位点特异地切开DNA链。大部分这类酶都以同二聚体的形式结合到DNA上,因而识别的是对称序列。在基因工程操作中非常有用,已有很多商品化的酶。
Ⅲ型限制性内切酶:也是兼有限制-修饰两种功能的酶。它们在识别位点3′端24~26个碱基对处切开DNA链。如EcoP I由两个亚基组成,识别序列不对称,酶切位点也不在识别位点上。
Ⅰ型酶和Ⅲ型酶通常兼有修饰和切割DNA的特性;作用除了需要镁离子作为催化反应辅助因子外,还需要ATP和S-腺苷酰甲硫氨酸(SAM)等辅助因子;酶切位点和识别序列不同,且切割产物无特异性,因而在基因工程中不具备实用价值。在基因工程操作中主要采用的是Ⅱ型限制性内切酶(restriction endonucleaseⅡ)。
(三)Ⅱ型限制性内切酶
1.识别序列和切割后形成的末端 Ⅱ型限制性内切酶是相对简单的酶蛋白。在基因工程中主要用的酶,如果不特别指出,一般所说的限制性内切酶就是指Ⅱ型酶。其识别序列一般为4~6个bp的核苷酸序列,少数为8个或更多,这一序列无论从正链还是负链读均相同,称为回文结构。Ⅱ型酶能识别其特异的识别序列,并在这个序列上切割产生特异的DNA片段,切割位点固定、可知。以核酸内切的方式水解DNA链中的磷酸二酯键,产生的DNA片段5′为磷酸基,3′端为羟基。根据切割位点相对于双重对称轴的位置,其酶切主要有下列两种方式。
(1)在识别序列的对称轴上将DNA双链同时切割,产生平末端,或称钝性末端。如SmaⅠ的识别序列是:5′-CCCGGG-3′,它的切点在G和C之间,在对称轴上将DNA切开,形成两个平末端(图3-1)。
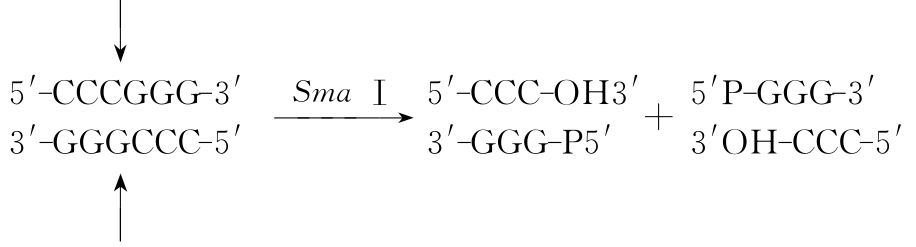
图3-1 限制性内切酶切割DNA后产生两个平末端
(2)在识别位点对称轴两侧相同位置分别切割DNA分子,形成具有黏性末端的DNA片段。包括5′黏性末端和3′黏性末端两种情况。
如EcoRⅠ在其识别序列的双侧末端切割DNA双链,于对称轴5′末端切割,产生5′端突出的黏性末端。而PstⅠ在其识别序列对称轴3′末端切割DNA双链,产生3′端突出的黏性末端(图3-2)。
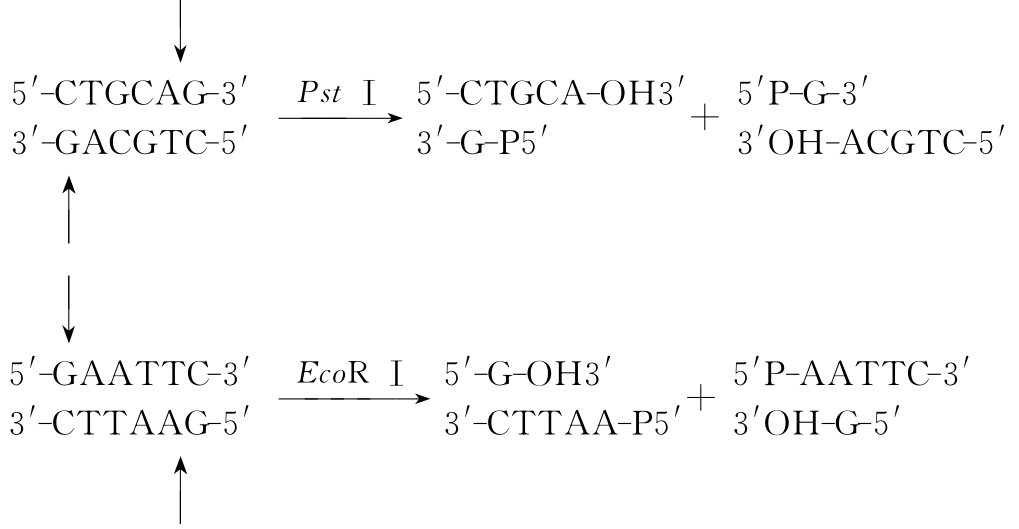
图3-2 限制性内切酶切割DNA后产生黏性末端
2.影响酶切反应的因素 一单位的限制性内切酶活性指在一定时间内(60分钟),适当温度下,在50μl反应体系中,将1μg的底物DNA完全消化所需的酶量,定义为一个活性单位。目前绝大部分的内切酶以噬菌体DNA(λDNA)作为底物测定其活性单位。活性单位依所用底物的不同而不同;使用其他底物时,应根据情况确定所用的酶量。影响酶切反应的因素主要有以下方面。
(1)DNA纯度:DNA作为内切酶的底物,其纯度的高低,很大程度上影响酶切消化反应的效率。DNA溶液中的某些物质会对限制性内切酶的活性有不同程度的影响,例如酚、蛋白质、乙醇、碘醋酸、EDTA、SDS及过高浓度的盐离子等。为了提高限制性内切酶对纯度差的DNA的反应效率,可以采用以下方法:①增加限制性内切酶的量;②延长酶切反应时间;③增加酶切反应体积,使杂质浓度稀释等。如果无法通过这些方法得到较满意的酶切效率,必须进行DNA的纯化。
(2)DNA的分子构型:不同分子构型的底物也会影响限制性内切酶的切割效率。有些酶对于超螺旋质粒DNA或病毒DNA进行酶切所需的酶量,就比酶切线性DNA高出许多倍。对于不同的酶切位点,其酶切效率也有差异。
(3)反应温度:不同的内切酶有不同的酶切反应温度。大多数内切酶的最佳反应温度为37℃,但也有些低于或高于37℃。酶切反应高于或低于最适反应温度,均会对内切酶活性产生影响。有关酶解温度的资料可以查阅销售公司的产品资料。
(4)反应缓冲液:每一种内切酶都有最适合的反应条件,不同的酶要求不同的反应缓冲液,甚至不同供应商的同一种内切酶的最佳作用缓冲液也不尽相同。内切酶的厂商都配有与该厂出售酶相配套的10倍酶切缓冲液以保证其100%的活力,其成分主要包括氯化钠或氯化钾、氯化镁、二硫苏糖醇(DTT)或β-巯基乙醇、Tris-HCl和(或)牛血清白蛋白(BSA)等。
在实验中,经常遇到需要在同一缓冲体系中同时进行2~3种内切酶反应的情况,此时选择对两个酶均合适的缓冲液很重要。各个厂商都会提供有关各种酶在不同缓冲液中活性发挥程度的百分率资料,可供寻找几种酶都能发挥很好活性的共同的缓冲液。如果找不到同时能满足两种酶降解作用的适宜的缓冲液,双酶切反应可以分两步进行:首先,用需要低盐浓度的限制性内切酶降解底物,然后用少量体积的高盐溶液把体系盐浓度调到第2种酶所需要的浓度,然后再加入第2种酶并完成第2个酶切反应。如果这两种酶作用的最佳条件不仅仅是盐离子浓度的区别,那么就只好分别进行酶切作用,即先用一种酶切割后回收底物,再进行第2次酶切反应。
3.同工异源酶(isoschizomer)和同尾酶(isocaudarner) 同工异源酶又称同裂酶,是指来源不同的一组限制性内切酶,它们名称不同,但具有相同的识别序列和切割位点。如HindⅢ、HsuⅠ、BhrⅠ和ChuⅠ互为同裂酶(图3-3)。
![]()
图3-3 同工异源酶具有相同的识别序列和切割位点
同尾酶是指一组限制性内切酶虽然名称不同,识别序列也不同,但切割后能产生相同的黏性末端。由同尾酶产生的DNA片段,可通过相同的黏性末端之间的互补作用而达到连接。但是连接后的酶切位点有可能消失。如MboⅠ,BamHⅠ和BglⅡ就是同尾酶(图3-4)。
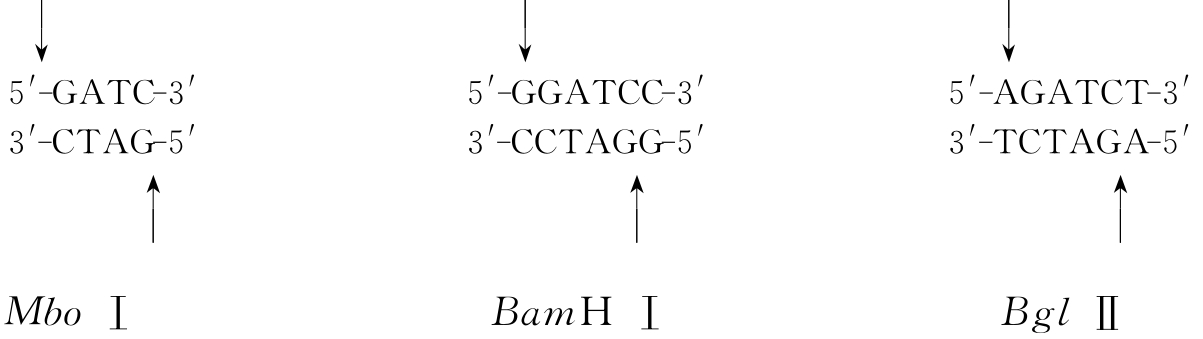
图3-4 同尾酶切割DNA产生相同的末端
二、DNA连接酶
DNA连接酶是基因工程操作中另一类很重要的酶。当进行DNA片段重组时,如果两个DNA片段具有相同的黏性末端,可以依靠互补碱基之间的氢键相互黏连在一起,但因并未形成共价键,这样的分子连接不够稳定,必须使用连接酶使DNA的3′端羟基和5′端磷酸基之间形成3′,5′-磷酸二酯键才能稳定。连接酶有两种:T4噬菌体DNA连接酶和大肠埃希菌DNA连接酶,在重组DNA技术中常用前者。
1.T4DNA连接酶 是从T4噬菌体感染的大肠埃希菌中分离纯化而来,它是T4噬菌体基因编码的产物,为一种单链多肽酶,催化活性需要Mg2+作为辅助因子和ATP提供能量。它不但能把两个具有相同黏性末端的DNA分子连接在一起,而且也能把两个平末端的DNA分子连接起来。连接两个DNA分子的先决条件是它们的末端必须靠拢在一起,所以连接黏性末端的分子比连接平末端分子的效率要高得多。
2.大肠埃希菌DNA连接酶 大肠埃希菌DNA连接酶是相对分子质量为74000的单一多肽链,催化DNA片段连接反应的机制与T4DNA连接酶相同。两者的不同在于大肠埃希菌DNA连接酶的催化反应需要烟酰胺腺嘌呤二核苷酸(辅酶Ⅰ,即NAD)的参加,而T4DNA连接酶需要的是ATP。大肠埃希菌DNA连接酶在催化各种底物的连接反应时有一定限制,不能对平末端DNA分子之间的连接起作用。因此,大肠埃希菌DNA连接酶在分子克隆中的应用不如T4DNA连接酶广泛。
三、DNA聚合酶
DNA聚合酶是一类能够将脱氧核糖核苷酸连续加至双链DNA分子引物链的3′-OH末端,催化核苷酸聚合反应的酶。在基因工程中应用较多的DNA聚合酶主要有大肠埃希菌DNA聚合酶Ⅰ、大肠埃希菌DNA酶Ⅰ的大片段(Klenow酶)、T4DNA聚合酶、T7DNA聚合酶及反转录酶等。
1.大肠埃希菌DNA聚合酶Ⅰ 是一种单链多肽,有3种酶活性:5′→3′聚合酶活性、5′→3′核酸外切酶活性和3′→5′核酸外切酶活性。主要用于DNA缺口平移制备核酸分子杂交探针及DNA3′黏端的末端标记。
2.大肠埃希菌DNA聚合酶Ⅰ大片段 是大肠埃希菌DNA聚合酶Ⅰ经枯草杆菌蛋白酶或胰蛋白酶水解后得到的大片段,保留了完整的5′→3′聚合酶活性和3′→5′核酸外切酶活性,但失去了5′→3′核酸外切酶活性。主要用于:①随机引物标记探针;②补平DNA5′黏端或进行3′端标记;③合成cDNA的第2条链和进行DNA序列测定。
3.Taq DNA聚合酶 是从耐热菌T.aquaticus中分离获得的,具有5′→3′聚合酶活性和依赖于聚合作用的核酸外切酶活性。其特点是在DNA变性温度下仍具有很高活性,因此能以高温变性的DNA为模板合成DNA新链,最适反应温度为75~80℃,主要用于聚合酶链反应(PCR)在体外扩增DNA。
4.反转录酶 催化以RNA为模板合成DNA的反应,产物为cDNA。目前有两种:一种来源于禽成髓细胞瘤病毒;另一种则分离自克隆有Moloney鼠白血病病毒反转录酶基因的大肠埃希菌。两者均具有5′→3′聚合酶活性,能以RNA或DNA为模板合成DNA;具有RNA酶H活性,即核糖核酸外切酶活性,以5′→3′方向或3′→5′方向特异降解RNA-DNA杂交分子中的RNA链。
四、末端修饰酶
一大类DNA末端修饰酶也是基因工程中的重要的工具酶,主要包括以下几种类型。
1.末端脱氧核苷酸转移酶(terminal deoxynucleotidyl transferase,TdT) 简称末端转移酶,是一种无需引物即能将若干碱基加在双链或单链DNA3′端的酶,利用这一点可以进行DNA3′端加尾和标记。该酶是一种非特异性酶,4种dNTP中任何一种均可作为其前体物。反应底物可以是带有3′-OH的单链DNA,也可以是3′突出的双链DNA,反应时需Mg2+存在。对于平末端或5′端伸出的双链DNA片段,也可以通过这种酶催化反应,在它们的3′端加脱氧核糖核苷酸,但是必须用Co2+。在基因工程中,末端转移酶可以用于给载体和外源DNA片段末端加上互补的同聚尾,使外源DNA与载体分子重组起来及DNA片段的3′端标记。
2.T4多核苷酸激酶 是从T4噬菌体感染的大肠埃希菌中分离获得的,该酶催化γ-磷酸基从ATP转移到DNA、RNA、寡核苷酸或核苷3′-磷酸等分子的5′-羟基端。可用于DNA的5′-羟基标记;使缺失5′-P端的DNA或寡核苷酸磷酸化。
3.碱性磷酸酶 主要有两种:一种来自大肠埃希菌,称为细菌碱性磷酸酶(bacterial alkaline phosphatase,BAP);另一种分离自小牛肠,称为小牛肠碱性磷酸酶(calf alkalinephosphatase,CIP)。是在碱性pH时具有活性的磷酸单酯酶,可特异地切去DNA或RNA5′磷酸基团,从而使DNA或RNA片段末端的5′-P转变为5′-OH。这就是所谓的核酸分子脱磷酸化作用。具有5′-OH末端的产物在T4多核苷酸激酶和ATP(γ-32 P)作用下,可被带上放射性标记物。在DNA体外重组操作中,载体DNA经过碱性磷酸酶处理后失去5′磷酸基,不能自己连接为二聚体,从而防止了载体的自身环化,提高克隆效率。
4.核酸酶S1 可以降解单链DNA和RNA,产生5′磷酸化单核苷酸或寡聚核苷酸,又称单链特异的核酸酶。在基因工程中,可以利用它把黏性末端的DNA切成平末端。在反转录反应合成cDNA的过程中,可利用它将所形成的发夹切除,形成无发夹结构的cDNA。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











