



第十章 病毒-基因化学
研究波长的结果表明,孢子发生的减少和突变频率的最大活性都在265纳米处。这正好是染色体内核酸最大吸收的波长范围,在这个范围内没有发现有关蛋白质的吸收最大值……要阐明胸腺核酸和真正遗传物质之间的详细关系还有待于作进一步的研究。
(纳普等,1939,304)
(TMV)蛋白质的原来结构,(在去除核酸以后)全部被保存下来。而以烟草进行的生物学试验……却发现,在核酸被分离后,生物学活性实际上全消失了。因此,在TMV繁殖过程中,核酸是必不可少的。
(施拉姆,1941,536)
施拉姆的报告尚未得到证实,该报告说,(核酸磷酸酶)酶能完全除去病毒里的核酸并产生一种无活力的无磷蛋白质,它的大小、超离心的均匀性和电泳迁移率,都与完整的病毒一样。
(库恩、斯坦利,1942,870)
……烟草花叶病毒分子辐射突变的产生,可追溯到病毒分子核酸部分的量变和质变,特别是因为较高原子序数的磷原子的积聚可用来说明最高的辐射吸收。
(普凡科赫、考施、施图贝,1940,255)
从受(芜菁黄叶镶嵌病毒)感染的植株中已分离和结晶出两种蛋白质。一种是核蛋白……另一种似是没有核酸的同一种蛋白质……感染植株的只是核蛋白,而病毒繁殖似乎一定要有核酸存在。
(马卡姆、马修斯、史密斯,1948,90)
病毒-基因化学
最早确定遗传物质是在研究流行病学过程中实现的,初听起来,一定觉得十分奇怪。当然,这只是因为用直接法实在难以确定遗传物质。高级有机体的染色体中,核酸同组蛋白结合在一起,并以复杂的形式与球状蛋白相缔合。试图除去染色体中的蛋白质或核酸成分,并看看染色体分解与否,这只能确定哪一种成分是染色体线的结构框架。在1939年的爱丁堡会议上,丹尼尔·梅齐亚和L.耶格报告了对唾腺染色体的酶学研究,他们确信存在着“由鱼精蛋白和组蛋白组成的连续蛋白质框架”,并否定核酸分子是保持染色体结构完整性所不可缺少的(梅齐亚和耶格,1939,213)。
七年之后,在实验生物学学会举行的核酸讨论会上又提出了同样的课题,斯特德曼等否定了精核几乎全由核蛋白组成的传统观点。我们可以回想一下,米歇尔已发现精子头部中81.6%是由核蛋白所组成。施米德伯格了解到米歇尔的提取方法(用稀释的盐酸)不够彻底,他算出了遗留下来的核蛋白,再加上米歇尔提取的81.6%共达96%。这一观点得到了米尔斯基和波林斯脱尔的支持,在1942年引进了克分子盐抽提技术,当他们于1946年把这种技术应用于鳟鱼精子时,得到的核蛋白含量为91%。但是,他们感到余下的9%很重要(米尔斯基、波林斯脱尔的论文《染色素》,114)。他们认为,这里面含有另一种核成分。为此,他们又写了一篇题为《染色素——细胞核的脱氧核糖核蛋白复合物》的论文。在泰勒编的《分子遗传学经典论文集》里,读者可找到他们写的第一篇论文,但没有第二篇论文。在第二篇论文里,米尔斯基和波林斯脱尔论述了非组蛋白蛋白质,它们含有构成核蛋白27%的色氨酸。他们论证说,若把他们用克分子氯化钠技术从细胞核中抽提出的物质,作为核蛋白,那显然是错了;因为抽提到的物质是核蛋白与“残留蛋白质”的复合物。由于这种复合物几乎构成了全部染色质,因此称之为“染色素”。
这些作者能够指出,胸腺核酸成分的传统观点已遭埃德加和埃伦·斯特德曼的反对。因为他们用米歇尔的抽提方法,发现在精子头部中核蛋白只占70%。他们认为其余30%是比鱼精蛋白和组蛋白更高级的蛋白质,他们称之为“染色体朊”(chromosomin)(1943,1947,239)。他们声称,这种染色体朊是一组蛋白质,它们的氨基酸含量随物种而异。染色体朊的蛋白质不同于组蛋白,是酸性蛋白质,同卡斯帕森在唾腺染色体间带观察到的吸收光谱的蛋白质一样。卡斯帕森认为:“这种蛋白质并非只是中期核特有的合成产物,间带蛋白质显然是由一部分染色体朊的连续线所构成,这些线贯串在染色体的全长。”(斯特德曼,1947,250)现在斯特德曼等能够回答化学所不能回答的遗传多样性的微妙问题了,因为细胞核的化学组成毕竟不是简单划一的,而是多样的,取决于染色体中染色体朊成分的性质。另一方面,核鱼精蛋白确实是核液的组成成分!核鱼精蛋白一定是在核周期的某些阶段浓缩在染色体上面(例如在精子头部里),但这种状态是短暂的(同上,247)。
1947年,在冷泉港举行的核酸和核蛋白的学术讨论会上,关于细胞核的化学组成莫衷一是。会上,米尔斯基的论文谈到染色体中含残留色氨酸的蛋白质所占百分比有很大差别——鱼的红血球中为5%,而在肝染色体中则占50%。在随后的讨论中,西蒙·库恩提出可否废除“染色素”这个术语。米尔斯基回答说:
本篇论文没有用染色素这个术语,这清楚地说明我们已经废除了这个术语,因为我们对所制备的“染色素”的性质已有更好的了解……
(米尔斯基,1947b,146)
玛格丽特·麦克唐纳表示她感到迷惑不解:
米尔斯基博士显然用同样的方法制备出两种物质:第一种只含核酸和组蛋白;第二种含核酸、组蛋白和含色氨酸的蛋白质。他是否能从技术角度告诉我们,为什么在制备的第一种物质中没有发现含色氨酸的蛋白质……
米尔斯基回答说:
在我们第一项工作中,由于反复沉淀、溶解和离心除去了大部分含色氨酸的蛋白质。在刚提交的论文所谈到的工作中,由于用了完全不同的方法,所以清楚地确定了含色氨酸的蛋白质。
(米尔斯基,1947b,146)
生物学家需要了解抽提出来的染色体的化学组成和性质。米尔斯基和波林斯脱尔为生物学家提供了详尽的技术细节。这表明生物化学家必须付出巨大的精力,才能满足生物学家的这种需求,而且要评价得到的数据也是困难重重。高等生物的染色体没有提供生物学家所要求的简单答案。
但是,病毒的情况又怎样呢?从二十世纪三十年代后期起,人们已意识到病毒与基因之间的类似之处,并用结晶的病毒材料来解答基因的化学问题。由于动植物病毒的研究具有重大的经济价值,因此,病毒研究的传统是没有国界的。细菌性病毒此刻也成了热门的研究课题,因为有希望由此来控制细菌性疾病。结晶的植物病毒,烟草、西红柿和土豆的病原体,都是解决这个问题的研究对象,但最好的还是烟草花叶病毒,即TMV。
TMV的化学
这是一段不同寻常的历史。由于用蛋白质沉淀剂成功地沉淀了 TMV,因此提出了病毒是蛋白质的观点(穆尔瓦尼,1926)。一年之后,纽约博伊斯-汤普森植物研究所的C.G.文森宣称“在醋酸铅的沉淀产物中”,他发现有氮。继此之后,巴顿-赖特和麦克贝恩也宣布得到了无氮的TMV晶体(1933)。罗瑟阿姆斯特德试验站的病理学部马上对这项成果进行了验证。他们发现巴顿-赖特-麦克贝恩获得的晶体却是来自盐析剂的磷酸二氢钾,TMV是不纯的(考德威尔,1934)。
翌年,斯坦利在《科学》杂志上发表了一篇引人注目的论文《分离具有TMV性质的晶体蛋白质》。这位洛克菲勒研究所的研究员从感染的土耳其烟叶的叶汁中,抽提出一种物质,其活性比原来的叶汁大1000倍。斯坦利声称,他“有充分的证据证明本文提到的晶体蛋白质,或是纯的蛋白质或是一种固体的蛋白质溶液”(1985,644)。因此,TMV是“一种自体催化蛋白质,现在可认为在繁殖时需要有活细胞存在”(同上)。
在英国,病毒研究最早是针对有重要经济价值的土豆病毒X,烟草不是该国的经济作物。在贝特森的影响下,R.N.萨洛蒙从1906年到1925年,早就在他的家乡巴利开始了土豆的遗传学研究。1926年,他写了一本小册子,总结了他的研究成果,并终于影响了农业部的丹尼尔·霍尔爵士,在剑桥大学农业系设立了土豆病毒研究组。肯尼思·史密斯在那里开始了他后来称为土豆病毒X的研究工作。由于这项工作取得了进展,使生物化学系的诺曼·皮里能同罗瑟阿姆斯特德试验站的弗雷德里克·鲍登合作,开始了抽提悬浮状态的活性因子。他们表明,土豆病毒X含有大量的氮和血清活性,当他们把同样的技术用于TMV时,证实了斯坦利的大部分发现。在他们撰写的关于TMV的最早出版物中,提到了在TMV含有2.5%的碳水化合物和0.5%的磷。五个月之后,他们认为TMV是一种核蛋白(鲍登、皮里,1937)。马卡姆在1938年参加了皮里的工作,并在战后继续进行研究。他说,在TMV中总是有磷存在。这位洛克菲勒研究所的分析人员一定把核酸留在乙醇沉淀病毒后的上清液中。因此,斯坦利实际上丢失了大部分核酸。一年后,当斯坦利承认在山茱萸嵌合病毒中有磷时,他认为这是由于核酸而不是感染粒子的一部分。他声称:“在pH值为8或9的情况后透析,有可能除去核酸,得到具有病毒活性的无磷蛋白质……”(斯坦利,1937,329)
斯坦利的工作,是博伊斯·汤普森研究所在路易斯·孔克尔指导下,由文森开始的研究计划的继续。孔克尔于1931年离开该研究所前往普林斯顿的洛克菲勒研究所去领导刚成立的植物病理学部。从一开始他就任用了斯坦利。正是年轻的斯坦利把文森的醋酸铅技术同诺思罗普的蛋白质水解酶的结晶法结合起来,得到了TMV液晶制备物。斯坦利的抽提技术和广泛研究TMV的失活因子,使他指出了TMV的蛋白质性质。其他研究人员(德沃夏克,1927;珀迪,1929)也证实了TMV的血清活性,这是TMV的蛋白质性质的明证。
在1936/1937年冬,斯坦利完全有理由贬低鲍登和皮里分离的磷和碳水化合物——毕竟只有0.5%的磷。即使核酸是TMV的真正组成成分,它也不可能有重要功能,因为RNA仍被认为是四核苷酸(米尔伯克、乔帕斯,1935),而且权威的利文仍支持这种看法。不仅如此,洛克菲勒研究所普林斯顿分部的研究人员同纽约实验室的工作人员一样,都坚信中心法则的蛋白质版本。认为病毒活性和专一性与某种核蛋白有关而不只是同蛋白质有关,这种看法如果没有充分的证据,那么在利文、兰德斯坦纳和米尔斯基的洛克菲勒研究所里以及在萨默、诺思罗普和库尼茨进行酶结晶的研究机构里,都会被视为异端邪说。TMV一定是蛋白质,而且是一种有自体催化能力的蛋白质。
一年以后,斯坦利出来支持TMV具有核蛋白性质。1937年12月,在印第安纳波利斯举行的一次讨论会上,他说:“迄今分离到的病毒蛋白质已发现是核蛋白。”(斯坦利,1938,119)由于他不止一次地把病毒着作蛋白质,因此他的这个讲话是事出有因的。1937年初,他描述了不同于TMV蛋白质的山茱萸嵌合病毒的晶体蛋白质,认为这证明“两种不同的病毒株产生两种不同的蛋白质”(斯坦利,1937,340)。病毒具有蛋白质性质的想法的确造成了危害,因为,像穆勒和德布吕克这些有影响的科学家,在他们早期探索遗传物质时就持这种观点。
奇怪的是,虽然病毒与染色体的化学成分相似,斯坦利却不认为它们之间存在相似性。相反,他提出由于“核酸和高分子量蛋白质的这种组合,使我们完全可以在单个分子中实现这种结构,从而使之具有类似生命的特性”(斯坦利,1938,119)。斯坦利认为,病毒繁殖的机制就是原生质生长的机制。生长中的重大事件,不是细胞分裂而是导致细胞分裂的生长过程。“我想,我们现在可以不考虑细胞分裂从而繁殖的机制,因为我们感兴趣的是一种基本的机制,即导致细胞生长直到分裂机制起作用的那种机制。”(斯坦利,1938,120)当然,原生质的生长也是以细胞内的酶为介导的一种蛋白质合成。
我们现在回顾对蛋白质水解酶、pH、热和亚硝酸等作用所作的早期研究时,可以看出这些研究成果同现在的观点完全一致,即核酸的重要性压倒一切。是否这个证据太好了?普凡科赫探测到来自两个有关病毒株的核酸在电泳迁移率上有差别,而蛋白质之间却没有差别,这导致他提出了病毒的生物学上的差异起因于病毒的核酸部分(普凡科赫,1940)。对此我们怎么看?或者看一下5个有关的病毒株的核蛋白在电泳迁移率上也有不同。这些作者确信,产生这些病毒株的突变起因于病毒分子的核酸部分发生了量变和质变,特别是辐射的最大吸收是由于原子序数更高(磷)的原子的积聚(普凡科赫、考施、施图贝,1940)。或者,我们可以再看六年后在德皇威廉研究所做的同样实验,他们取得了所研究的每种病毒核蛋白随pH而变化的电泳迁移特性曲线(施拉姆和雷本斯伯格,1942)。但如西摩·科恩所说:“这些突变体在迁移率上的差别,并未反映突变体的RNA在迁移率上的有什么不同。也许这些差异只反映了某些磷酸盐穿过蛋白质外壳的程度,以及在多种折叠的蛋白质亚单位中羧基的暴露程度。”(科恩,1973)
如果认为德国研究人员是正确的,而美国研究人员是错误的,那就大错特错了。1948年,梅尔彻斯回忆说,施拉姆在病毒核蛋白的蛋白质部分中,而不是在核酸部分中去寻找病毒株差异的化学基础(梅尔彻斯,1948,116)。在美国,只用普通的和主脉TMV株重复了电泳迁移率研究,结果只观察到一带条(奈特、劳弗,1942)。施拉姆和穆勒试验用乙酰或苯脲基酸基团“覆盖”TMV的氨基酸基团,虽然完全覆盖了氨基酸基团,但病毒的感染性一点也不低于未经处理的。在美国,也进行了同样的实验,结果发现在完全覆盖之前,病毒的感染性有所减弱(米勒、斯坦利,1941);加上德国的研究工作表明磷酸酶能使TMV失活(普凡科赫、考兹,1938;1939)也没有被洛克菲勒研究所的洛林或科恩所证实。因此,在英国和美国,人们对德国人的工作失去信任,也不足为奇了。然而,尽管发生了战争,美国普林斯顿的斯坦利研究小组继施拉姆及其同事们之后进一步作病毒的生物物理学研究,其兴趣不亚于罗瑟阿姆斯特德试验站的工作。美国人和德国人都能得到对方的所有论文。科恩回忆说:“罗瑟阿姆斯特德试验站与普林斯顿实验室各执己见。他们之间意见分歧的程度,至少达到了他们与德国实验室之间不一致的程度。我从未听到过由于政治原因而批评施拉姆的工作。为了弄确实,我们重复别人做过的每一件工作。”(科恩,1973)
1943年,施拉姆观察到pH为9时,TMV断裂成大小一样的两种亚单位。一种含核酸,另一种没有核酸,但它们的粒子的相对分子质量都是360000,接近于从单位晶胞得到的相对分子质量(370000)(据贝尔纳、范库亨,1941)。但令施拉姆奇怪的是,当他把这些亚单位回到pH 5时,在超速离心机里再也找不到它们了。他发现的却是像原来TMV那样的特征形态,也没有把蛋白质亚单位结合在一起。无论是亚单位还是重组的无核酸粒子,都没有感染性。战后,他把这项研究工作推进到一个新阶段。那时,他在去除了蛋白质后,完整地保存了TMV的核酸。但没有证明这种RNA核心有感染性(施拉姆,1948)。
又经历了漫长的八年之后,才证实TMV的RNA核心的感染性(吉雷尔、施拉姆;1956;弗伦克尔·康拉特,1956)。即使在那时,也没有技术可防止核糖核酸酶使RNA失去感染性的解聚作用。除去蛋白质外壳就能使RNA核心受到酶作用的看法,“一点也不明显”(梅尔彻斯,1965,120),病毒亚单位的重组,可恢复感染性,被认为是“生命基本系统的‘再合成’”(斯坦利,1955),当然蛋白质在其中起决定性作用。这些病毒学家似乎陷入了库恩的框框。克里克回想说,他“曾打算说服(剑桥大学的)马卡姆和史密斯做重组试验——后来弗伦克尔-康拉特做了——但是,他们不愿认真地采纳我的建议”(克里克,1973)。加利福尼亚的病毒学家,也不见得比他们的剑桥同行来得开明。早在1942年,他们曾试图证明去除苏氨酸的病毒是一种实验室突变体。结果表明,去除了7%苏氨酸含量的病毒是能感染的,而且产生了正常的含苏氨酸的病毒颗粒。但是,他们却仍无动于衷(哈里斯、奈特,1952)。
那么,从TMV研究中得到什么结果呢?如果说延误了一些时间,但终究确证了携带病毒的遗传特异性的不是病毒蛋白质,而是病毒的RNA。但在二十世纪三十年代后期,他们就表明了研究精子头部未能揭示的内容,即感染粒子完全是由核蛋白所组成。
病毒的失活和突变
要进一步分析——确定遗传特异性的载体和感染、复制的因子是核酸还是蛋白质——需要讨论化学和物理诱变剂对病毒的作用。斯坦利在这方面碰到了困难,因为亚硝酸似乎并不破坏山茱萸嵌合病毒的蛋白质,但却能使病毒失活。由于他深信蛋白质是起决定作用的物质,因此,他认为上述结果是“奇怪的”(斯坦利,1938,331)。另一方面,TMV经蛋白质水解酶作用后即丧失感染力,可能是核糖核酸酶污染的结果。但是,紫外线的作用怎样呢?早在1928年(见表10.1),就把紫外线的杀菌作用同核酸成分的最大吸收联系了起来(盖茨,1928)。在1941年冷泉港会议上,斯塔德勒、霍兰德和埃蒙斯已清楚地知道紫外线诱变与核酸最大吸收之间的关系。但是,斯塔德勒倾向于认为观察到遗传标记的丢失,是由于破坏了作为染色体结构物质的DNA引起了染色体断裂(斯塔德勒,1939)。他当时正在寻找一种诱变剂,以引起基因的突变,而不仅是使遗传物质沿着染色体重排。他的困难在于如何来区分这两个过程。霍兰德和埃蒙斯在详述证明核酸与紫外线突变有关的有力证据后,又告诫与会者说:
在研究辐射对活细胞的作用时,过分强调核酸的重要性可能有点危险。在辐射引起的突变中,核酸很可能只是“吸收”剂,然后把吸收的能量传递给与它紧密相连的蛋白质。
(霍兰德、埃蒙斯,1941,185)
表10.1 紫外线引起的失活和突变
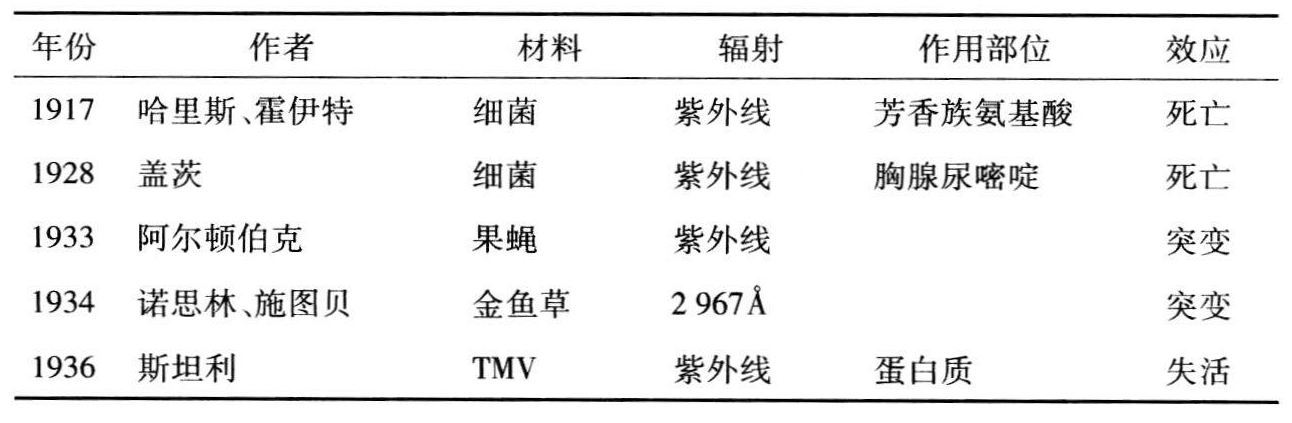
续表
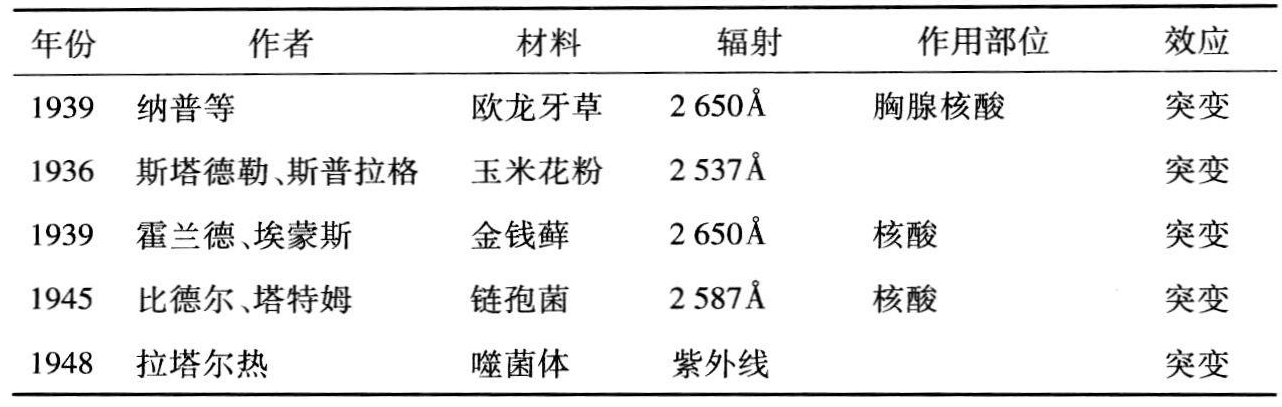

图10.1 虚线:以2600Å处作为吸收100%,胸腺核酸钠的相对吸收光谱;实线:在真菌中产生突变的相对效率;点画线:以2650Å处的效率作为100%,欧龙牙草孢子产生突变的相对效率(录自霍兰德、埃蒙斯,1941,183)。
他们所阐明的能量传递假说使他们得以推翻早先关于紫外线数据的框框,即“活细胞可能含有在2650Å处高度敏感浓度极低的重要化合物,或者细胞的形态结构在短波长处能保护其基本特征”(霍兰德、埃蒙斯,1939,400)。比德尔把能量传递假说,看作是另一种更简单的假说:“核酸是基因的一种成分……核酸可能是基因以外的,把产生突变的能量传递给基因。”(比德尔,1946,51)紫外线使噬菌体失活的发现也无助于问题的解决,因为不久发现了光复活现象(凯尔纳,1949;迪尔贝科,1949)。这不仅使所有研究噬菌体的人大吃一惊——用德布吕克的话来说,这是一次“惊人”的发现(德布吕克,1948)——而且使紫外线诱变作用变得复杂化了。再者,TMV紫外线失活光谱(见图10.2),更是使人误解。因为最大吸收大大低于核酸的最大吸收,这好像支持了斯坦利关于病毒遗传物质是蛋白质的看法。研究TMV的突变也存在特殊的实验困难,这是由于竞争效应同突变效应混淆起来了。后来,梅尔彻斯正确地强调了蒙德里和吉雷尔工作的重要性。他们的工作表明,用诱变剂稀释接种的TMV可使之部分失活,因而减弱了突变体和正常感染粒子之间的竞争。因此,要证明诱变剂提高了TMV的突变率,而不只是让自发突变体感染寄主,就必须用水稀释作为对照。这不是“在‘蛋糕上裱花’,而是必不可少的条件”(梅尔彻斯,1965,124)。在普通的烟叶上,病毒只有几百个进入点,但在典型的接种液里有几百万个病毒粒子。亚硝酸对TMV的诱变,为掌握遗传密码提供了关键的线索(1958),但这是在克服了研究TMV时遇到的各种艰难险阻,经历了数不清的失望之后,才得到最后的证明。
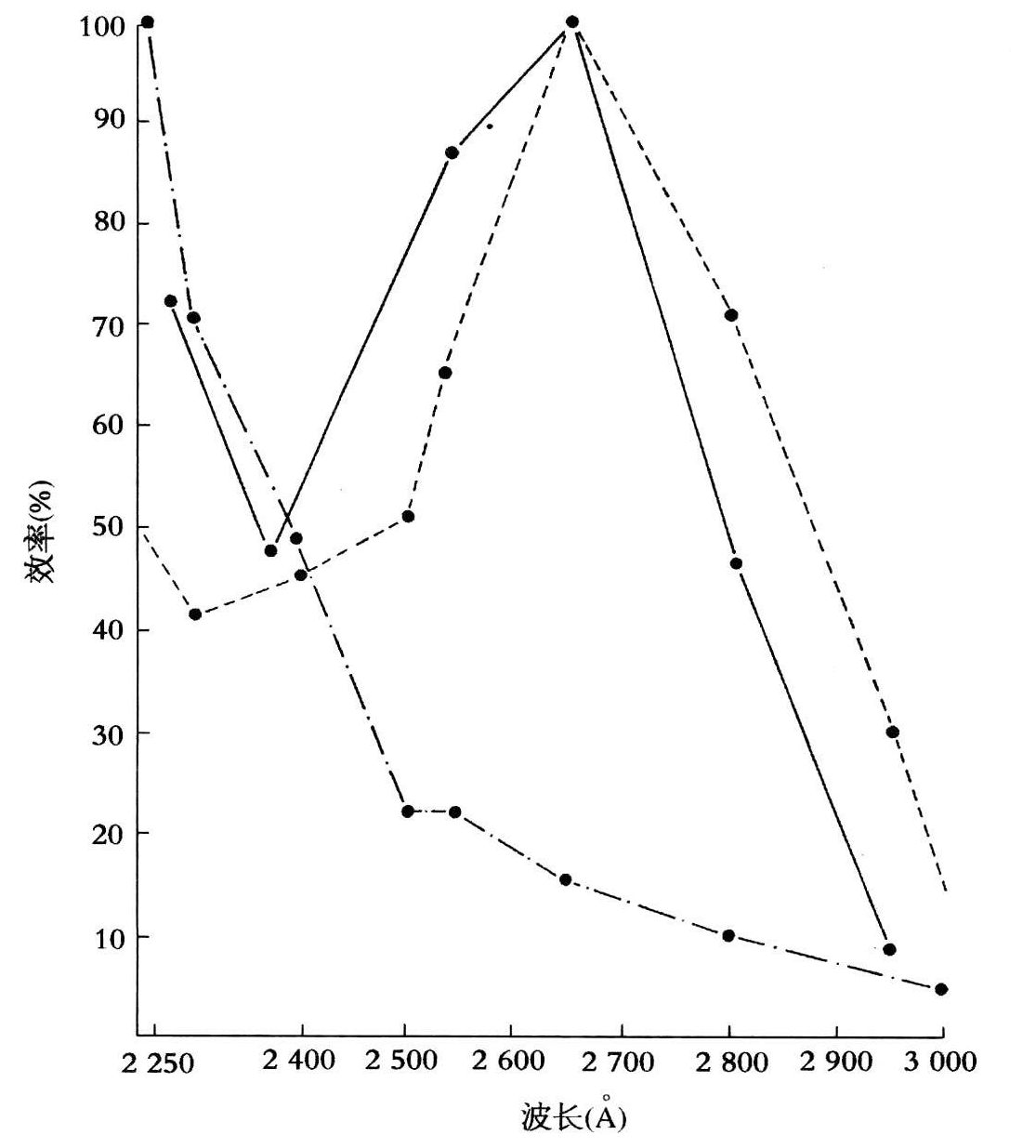
图10.2 杀真菌作用(实线),杀细菌作用(点画线)及烟草花叶病毒失活(虚线)的相对有效曲线(录自霍兰德、埃蒙斯,1939,400)。
病毒结构
这里讲的是研究病毒的诱变、化学和结构的国际活动,旨在弄清病毒复制的性质。这里有:洛克菲勒研究所普林斯顿分部的路易斯·孔克尔学派;与剑桥大学皮里合作的罗瑟阿姆斯特德试验站的鲍登学派;先在柏林后在土宾根的梅尔彻学派及剑桥大学莫尔顿诺研究所的马卡姆学派。还有些不那么出名的研究人员,如先在德国弗赖堡后在柏林生化研究所的普凡科赫,莱比锡卫生学研究所的欧文·威奈克和日本的高桥;当然,还有在剑桥大学及后来在伦敦伯克贝克学院工作的贝尔纳和范库亨,以及利兹大学的阿斯特伯里和贝尔这些在植物病毒学家影响下从事病毒研究的物理学家。阿斯特伯里在一篇稍稍有名的论文中说道:“毫无疑问,对分子生物学家来说,本世纪最振奋人心的发现是了解烟草花叶病毒的性质——它只是一种核蛋白。”(阿斯特伯里,1939b,125)
在刚开始研究病毒的日子里,认为活性粒子是球状的(贝希霍尔德,1902;贝希霍尔德、施勒辛格尔,1933;K.M.史密斯,1935,55~57)。我们在第一章里已提到,在研究双折射时,首次发现这一看法不适用于某些病毒(高桥、罗林斯,1932,1933;鲍登等,1936),而且根据X射线的数据估算TMV粒子的长宽比,也说明这一看法是错误的(鲍登等,1936,1052)。与此同时在瑞典的乌普萨拉用沉淀速度和电泳得到的相对分子质量为15000~20000的粒子(埃里克森-克文泽尔、斯维德伯格,1936)。斯坦利实验室的马克斯·劳弗继续进行这一研究工作。根据黏度测定,他计算出长度和宽度分别为480纳米和12.3纳米(劳弗,1938)。这样不对称的TMV粒子的相对分子质量,不可能在20000的范围里,而是要大得多——斯维德伯格错误地认为TMV是普通蛋白质的不对称性。劳弗计算的相对分子质量为42.5×106。根据TMV的物理化学特性推断出粒子呈细长形。一年之后,用电子显微镜拍摄到了这种细长形的TMV粒子(考兹、普凡科赫、吕斯卡,1939)。因此,病毒的实际情况,同生物物理学家的推断完全一致,这同称为染色体的难弄的东西成鲜明的对照。下一章中,我们将讨论植物病毒对物理学家的巨大影响。看来是大有希望的病毒又是多么捉摸不定。
我们应称赞阿斯特伯里朴实的研究热情而责备贝尔纳和范库亨的谨慎吗?过去的简单资料,不足以回答这个问题。贝尔纳及其合作者从X光衍射照片上早就发现了TMV粒子的亚单位结构。到1941年,他们估算,大的亚单位的体积为44Å×44Å×22Å,小的亚单位则为11Å×11Å×11Å。阿斯特伯里却赞同他们较早提出的估算——亚单位体积为22Å×20Å×20Å(贝尔纳、范库亨,1937,924),因为这一数值相当于一个核苷酸和54个或21×33个氨基酸(阿斯特伯里,1939,126)。但是,贝尔纳和范库亨并没有以这个角度去考虑问题。此外他们还觉得,认为亚单位结构在生物学上有重大意义还为时过早。
我们研究的这些粒子的晶体性质,在生物学上没有实质性意义,它不能解决这些粒子是否为感染因子的问题,也不能回答是否应把这些粒子视作生命有机体这一更玄奥的问题。
(贝尔纳、范库亨,1941,164)
他们虽然注意到了“晶体与纤维蛋白质在结构上的相似性”,但他们认为,病毒结构“不属于迄今研究过的已知的任何结构类型。有迹象表明,病毒的内部结构比晶体蛋白质分子的更简单”(贝尔纳、范库亨,1941,164)。
我们知道,阿斯特伯里曾以他历来的热情同贝尔纳讨论了TMV研究工作的重要性。因此,贝尔纳论文中提出的某些告诫,表示了他对阿斯特伯里作出的反应。然而遗憾的是,贝尔纳和范库亨在《普通生理学杂志》(1941)发表的54页的文章中,丝毫没有提到基因-病毒相似性问题。事实上,像美国斯坦利的研究小组一样,罗瑟阿姆斯特德试验站的研究小组对这样简单的相似性缺乏热情。毫无疑问,他们的这种态度也影响了贝尔纳。贝尔纳和范库亨对此漠不关心,却热心于研究测量小角度间距的方法,以揭示介于分子本身和光学显微镜所能观察到的结构之间的组织结构(见第十六章)。
生物物理学家和生物化学家认为大有希望的TMV,却被证明是捉摸不定的。TMV终究没有显示出自体繁殖单位的化学同一性。它使生物物理学家和生物化学家大失所望,部分原因是实验上的困难。另外,是没有人试图用TMV的无蛋白质RNA来感染烟草。西摩·科恩认为,他在斯坦利实验室用病毒热变性制备的RNA(科恩、斯坦利,1942)可能有感染能力(科恩,1973)。什么阻碍科恩取得进展呢?不必说,就是中心法则的蛋白质版本。对于马卡姆和施拉姆来说,情况也是如此。正是植物病毒学家的范例使他们无法揭示出含有基因和病毒遗传信息的化合物。这一任务反而由“局外的”研究小组——医学研究小组来完成。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












