



二、降解型基因工程菌的构建
构建基因工程菌的目的是使重组的新菌株有更高的降解能力,更强的适应生存能力。构建基因工程菌要在总体的构建策略指导下,在认真、详细分析现有相关降解基因、降解途径、降解调控的资料的基础上,按代谢步骤最少、调控最可靠、表达的酶量最大、重组菌存活能力最强、活性最高的原则,确定最终的代谢途径和受体菌株,然后开始实际的构建,由于难降解污染物千差万别,已有的降解基因(降解质粒)、可用降解受体菌又各不相同,因此实际的构建方法也是多种多样的。
1.工程菌构建的总体策略
刘和及陈英旭通过对难降解有机物生物降解机理及妨碍降解的因素进行分析,提出了构建高效基因工程菌的几种构建策略。
(1)优化调整降解有机物的降解途径
优化难降解有机物的降解途径是把不同的降解基因组合在一起,改变或扩展原有代谢途径、构建新的更合理、更易于调控的代谢途径,并使这些功能可以在一个受体菌株中得到表达。
①重组互补代谢途径:重组互补代谢途径是将与降解某一有机物的降解基因互补组合,使代谢过程顺畅,克服歧路、短路(产生毒性很大的代谢产物)、降解效率低等问题,从而构建出高效降解菌。
②改变代谢产物流向:生物降解过程中产生进入截止式代谢途径(end-product pathways)的中间代谢产物可以阻碍生物降解的进一步进行,把这种代谢产物(或其前体物质)导入另一种代谢途径可以使有机物得到彻底降解。这种重组方法实际上是把两种途径有机重组,克服代谢短路的现象。
③扩大对基质的利用范围:多种有机污染物共存于同一环境的现象十分普遍,使某一微生物同时具有二种或多种降解能力可以大大提高生物群体的降解能力。通过质粒转移或同源重组可以把不同的降解能力整合到一种菌株中。
(2)提高有机污染物的生物可利用性
生物可利用性是生物降解中的重要因素,提高生物可利用性和促进有机物进入细胞可以明显提高降解速率,从提高生物可利用性出发构建工程菌株也是提高生物降解能力的重要方面。
①重组表面活性剂编码基因:表面活性剂能通过改善疏水性化合物的水溶性和表面张力等理化性质而提高其生物可利用性。构建产生表面活性剂的基因工程菌或把表现活性剂编码基因和特定降解基因重组得到的基因工程菌都可以用于生物处理和生物修复,提高生物降解效率。
②重组有机污染物的跨膜转运基因:许多有机污染物要跨膜进入细胞才能被细胞内降解酶所降解,而许多有机物的跨膜运输需要专一性的转运蛋白,构建携带有跨膜转运系统蛋白编码基因的基因工程菌有助于有机物进入细胞内被降解。
(3)提高降解菌的活性及存活能力
降解菌的存活和高活性是生物降解的前提条件,使它们具有较强的竞争适应能力、较高的活性、更强的存活能力才能保证它们降解功能的实现。
①提高降解菌的抗毒能力:污染环境(包括污水、废弃物等)是一种极为复杂的环境,环境中存在的和生物降解过程中产生的对降解菌有毒的物质都会对生物降解过程产生重要的影响,针对具体过程构造具有较强抗毒能力的高效降解型基因工程菌可以保证生物降解的顺利进行。
②提高降解菌对不利环境因子的耐受能力:各种不利于细菌生长、繁殖的环境因子(如放射性、各种极端条件)也会不同程度影响降解菌的生物降解过程,构建对不利因子高耐受能力降解型基因工程菌有助于使它们在不利的环境条件下表达出降解酶,实现生物降解功能。
2.基因工程菌构建的具体方法
DNA重组技术的不断进步已经为基因工程菌的构建提供了极为繁多的方法,这里列出一些较常用的方法。
(1)转入一个或多个质粒到受体菌构建出具有新降解能力的菌株
利用降解性质粒的相容性,把能够降解不同基质的质粒结合到一个菌株中,组建成多质粒的新菌株,这样使一种微生物能够降解多种污染物或能完成降解过程的多个环节,还可以通过接合转移使不带降解性质粒的菌株带上降解性质粒,获得降解能力,这是选育高效降解菌的重要方法之一。
(2)降解性质粒DNA和染色体DNA的体外重组,构建高效基因工程菌
降解性质粒DNA和染色体DNA分子上具有能编码降解酶及有助于生物降解的基因,综合应用生物化学和微生物学的现代技术和手段,在体外对这些DNA分子进行剪切、加工,将这些与生物降解相关的基因在体外进行重组,然后转移到受体细胞中,再通过复制表达以及筛选可获得高效的基因工程菌。
(3)原生质体融合构建多种特性相结合的高效降解菌
原生质体融合技术可以利用微生物细胞共生或互生作用的机理,将多个细胞的优点集中到同一个细胞中,这种理念也被用于构建高效降解菌。PTA(purified terephthalic acid,PTA)是一种石油化工产品,广泛用于合成化纤、塑料、染料的生产,由于PTA是苯环化合物,对生物具有遗传毒性,导致PTA废水难以被土著菌快速降解,存在着处理效率低和处理费用高的问题。处理PTA废水需要高抗毒性、高降解性、高絮凝性的三高特效菌。程树培通过原生质体融合获得三高性能的特效工程菌株(Fhhh),Fhhh是由真核原核二界细胞的三个亲株菌的原生质体,跨界融合而成的基因工程菌,三株菌是黄孢原毛平革菌(霉菌)、酿酒酵母(酵母菌)、土著细菌。融合子同时含有霉菌的降解基因mnp、酵母菌的絮凝基因FLol和土著菌的DNA片段。
3.构建基因工程菌个例
(1)把甲苯生物转化成P-HBA(4-HBA)的高溶剂耐受性基因工程菌(恶臭假单胞菌)的构建
1)构建工程菌的目标。把能生物转化甲苯的二株细菌(恶臭假单胞菌DOT-TIE Pseudomonas putida DOT-TIE)和门多萨假单胞菌KRI(Pseudomonas mendocina KRI)的代谢途径和生理特性整合到一个菌株中,构建出一个对甲苯和P-HBA(P-hydroxybenzoate)有高耐受能力,能生物转化甲苯成P-HBA,并能大量积累有工业用途的P-HBA的基因工程菌。
2)构建对甲苯、P-HBA高耐受、高积累P-HBA工程菌的意义。甲苯是一种常见而严重污染环境的污染物,4-HBA是一种有附加值的化合物,可用于paraben和methyl paraben的合成,这些化合物又是合成液体玻璃和抗微生物剂的重要原料,这里把污染物变成另一种有资源价值的新化合物,这比简单降解污染物成H2O和CO2更有意义。
3)工程菌的构建过程
①出发菌株及其生理生化特征。Pseudomonas putida DOT-TIE:该菌株对溶剂有高度抗性,其辛醇/水分配系数的对数P(LogPow)高于2.5。其能利用甲苯作碳源和能源,矿化利用甲苯的代谢途径是Tod(toluene degradation)途径,甲苯先被氧化成3-甲基儿茶酚(3-methylcathecol),其后又通过间位断开途径依次转化成三羧酸循环的中间产物(图15-16途径A)。其和下述菌株一样也具有利用P-HBA(pobA基因编码酶的降解产物)的能力。
Pseudomonas mendocina KRI:该菌株也能利用甲苯作为碳源和能源,tmo(toluene monooxygenase)基因编码的T4MO(酶)(甲苯4-加单氧酶)活性把甲苯羟化成P-甲酚(P-cresol),其后pcu基因的产物又将甲酚氧化成4-HBA(4-hydroxybenzoate 4-羟基苯酸盐)。这种菌株的pob基因产物活性还可以把4-HBA羟化成3,4-二羟苯酸盐,其后再经邻位(ortho)断裂和β-Ketoadipate途径被进一步降解(图15-16途径B)。

图15-16 甲苯好氧代谢的两种降解途径。
(A)途径是恶臭假单胞菌DOT-TIE主导的由todClC2BA编码的甲苯双加氧酶和tod编码的cis-toluene dihydrodiol脱氢酶进行的。(B)途径是门多萨假单胞菌KRI的tmo、pcu(p-cresol utilization)和pob基因编码酶降解甲苯的过程。
②构建的基本思路。利用DOT-TIE的对甲苯、4-HBA高耐受性的背景,同时使todC失活(△todC),体外剔除(Knocked out)pobA基因,从而得到双突变体DOT-TIE△todCpobA。然后再把来源于P.mendocina KRI的tmo、pcu基因转到DOT-TIE△todCpobA的染色体上,在DOT-TIE背景表达tmo和pcu基因,从而得到能利用苯作为碳源和能源,并积累大量4-HBA的工程菌株。
③构建过程。构建过程按下述步骤进行:
(A)使tod基因失活,从能利用甲苯的DOT-TIE中选育出不利用甲苯的新菌株(DOTTIE(tod F'△todCl∷km∷'tod C2)。
首先使亲本菌株包括全部tod Cl基因和部分todF、todC2一个2.2kb的DNA序列缺失。带有插入缺失的质粒pMIR29(tod F'△tod Cl∷km∷'tod C2)和pMIR30(tod F'△tod Cl∷kiL Ate LAB∷'tod C2)被作为传递(delivery)质粒。通过同源重组,野生型的编码甲苯双加氧酶的基因(位于DOT-TIE带有缺失译本(deleted version)的染色体中)被取代。分离到两种不能利用甲苯作为碳源的突变体,分别定名为△tod C Km和△tod C tel。染色体中被删除的tod Cl基因可用PCR和Southern blotting来证明。两种突变株也具有良好的稳定性,在非选择条件(不含抗生素)的LB培养基传代90次,100%的细胞仍具有对卡拉霉素(km)和亚碲酸盐(tel)的抗性,同时在以甲苯为唯一碳源的M9液体培养基(没有标记的选择压力)中培养一周也未见生长,利用点种技术测出甲苯生长的回复速率低于10-9。
(B)使pobA基因失活,从DOT-TIE△tod C km中选育出△tod C pobA双缺失突变株。
以质粒pMIR31作为基因置换的传递系统。DOT-TIE△tod C km细胞与pMIR31作电转化,失活拷贝的野生型pob等位基因被同源重组,筛选到不能利用4-HBA作为碳源带有pipr标记(来自质粒pMIR31)的克隆。△tod C pobA双缺失突变株中野生型pobA基因失活拷贝的成功等位交换用Southern blotting证实。失活的pobA基因稳定性好,菌株在存在hydroxylated carboxylic acid条件下培养一星期后,仍然不会回复出利用4-HBA生长的能力。
(C)mini-Tn5Tc tmo/pcu稳定转移到DOT-TIE△todc pobA的染色体,使菌株获得转化甲苯积累4-HBA的能力。
来源于P.mendocina KRI的tmo和pcu基因被亚克隆到mini-Tn5 Tc,最后构建出pMIR44。
DOT-TIE△tod C pobA突变体与E.coli CC 118λpir(pMIR44)(AprTcr,15kb)在存在辅助有机体(helper organism)E.coli(pRK600)的条件下进行融合(mated),DOT-TIE的接合后体(exconjugants)在补充抗性标记抗生素(卡拉霉素、链霉素和四环素)的LB培养基上选择,受体的接合后体比率为5×10-8,分离出6个在加入葡萄糖的无机培养基和母体菌株生长得一样快的克隆,所有这些接合后体由于pobA突变而不能以甲苯或4-HBA为唯一碳源生长。菌株标记的稳定性被测试,6个菌株在不加抗生素的LB培养基上传10代,而后再稀释涂布在带有抗生素的培养基上,结果证明全部细胞仍然带有所有的抗生素抗性标记。通过Southern blotting方法可以检测分离细菌中转座子的不同插入位置。
所得菌株异化转化(catabolic turnover)甲苯产生4-HBA的能力被测定。一种方法使用休止细胞(resting cells)在提供葡萄糖的M9无机培养基中进行,甲苯以气态形式提供,细胞密度保持恒定(1010CFU/ml),试验结果表明所有菌株产生积累4-HBA能力都存在一个从3h到24h的延迟期,然后4-HBA浓度迅速提高,其中最优良的菌株(clon-24)在其产生4-HBA线性期内产率达到0.4mmol/(L·h),悬浮液中4-HBA的最高积累量达到12mmol/L。另一种方法是低密度细胞生长在添加葡萄糖的M9无机培养基中(甲苯以气态方式提供),结果表明检测到一定数量4-HBA所需时间和休止细胞是一样的,而且clon-24菌株积累4-HBA的最高浓度也可达到12mmol/L。
此外P.mendocina KRI tmo启动子(tmo promoter)的表达需要一个两成分信号转导系统(two-component signal transduction system)(tmoS/tmoT),而在DOT-TIE中正好也有与之(tmoS/tmoT)同源的todS/todT转导系统,其也能激活启动tmo基因的表达。
(2)降解2-氯甲苯工程菌(假单胞菌)的构建
1)背景。
①发展用于生物降解异生物源物质新生物体是我们环境保护中的一项极具挑战性的任务。对异生物源化合物代谢遗传和生化基础的理解使重组微生物更具有潜力,重组微生物可以用于城市与工业活动中所造成的环境污染的生物修复。
②氯化芳香烃化合物的降解是一个重要方面,至今没有能降解单取代氯甲苯的自然菌株被特征化,但能把甲苯生物转化成苯酸盐的P.putida mt-2质粒pwwo的上游TOL途径的某些酶相关的底物范围打开了代谢工程的有意义的可能性。
③TOL上游基因导入到3-氯苯酸盐降解菌(Pseudomonas.sp.B13)能降解2-氯甲苯和4-氯甲苯、3-氯甲苯。但未能产生对2-氯甲苯有活性的菌株。TOL质粒的xylAM基因的二甲苯单加氧酶复合酶能转化3-氯甲苯和4-氯甲苯成为相应氯苯酸盐。TOL二甲苯单加氧酶不能转化2-氯甲苯。
然而研究又发现,2-氯甲苯的甲基经单氧化(monooxydation)成2-chlorobenzylalcohol,但其后的氧化分解特别缓慢。2-chlorobenzylalcohol可以被广泛基质范围的xylb和xylc编码的TOL脱氢酶催化,产物是2-氯苯酸盐,从上可见使2-氯甲苯降解的关键活性是甲苯氧化成相应的醇。
最近的研究说明P.putida F1的环-双加氧酶(ring-dioxygenase)可以把2-氯甲苯转化成2-chlorobenzylalcohol。氯苯降解菌Burkholderia sp.ps12的酶活性成功攻击2-氯甲苯得到相应的benzylalcohol,其效率高于70%,但后面的代谢却是低速的。在两种情况下,催化甲苯/苯生物降解第一步的双加氧酶起到如单加氧酶的作用,能把2-氯甲苯转化成2-氯苯乙醇;这些观察提供了构建能完全降解难降解2-氯甲苯工程菌株的基础。
④构建降解2-氯甲苯工程菌构想:结合两种降解片段(从P.putida的TOL和TOD途径获得),得到一个杂合体(负责上游途径把2-氯甲苯转化成2-氯苯酸盐),遗传决定子重组到2-氯苯酸盐降解菌的染色体(后者提供下游途径),结合得到的重组菌有全部对2-氯甲苯和下游中间代谢产物所需的遗传信息和酶促活性。
对重组子的分析,可以同时分析降解动力学、酶促瓶颈、过程的能量平衡、副产物的产生和代谢途径的调控多方面的问题。研究结论也适合于许多难降解的氯代化合物。
2)构建代谢2-氯甲苯杂合途径的基本原理。
①选择和整合2-氯甲苯降解途径策略:选择降解2-氯甲苯降解途径的总体策略如图15-17所示。完全降解包括上游途径和下游途径。
上游途径,集合TOD和TOL两种降解的降解能力,把2-氯甲苯氧化成2-氯苯酸盐,todC1C2BA基因(源于P.putida F1)编码的广基质范围的甲苯双加氧酶把2-氯甲苯的甲基单氧化得到2-氯苯乙醇。xylBC基因(源于P.putida mt-2)编码的苯基醇脱氢酶和苯醛脱氢酶把2-氯苯醇进一步氧化成2-氯苯酸盐。
下游途径可以通过形成儿荼酚,然后进一步降解完成全部降解过程,这里可以有二种不同的降解方式,其一是2-氯苯酸盐被转化成了3-氯儿荼酚(3-chlorocatechol)(由P.aeruginosa JB2菌株进行),其后再被导入到一种修饰的邻位断裂途径被完全降解。其二是苯酸盐双加氧酶攻击2-氯苯酸盐的1和2位点(P.aeruginosa PA142),结果hydrodiol被转化成儿茶酚,再经housekeeping ortho途径进一步降解。
②上游降解基因的调控设想:设想构建出一个对tol和tod基因具有单一调控能力的Pu启动子和XylR蛋白(与TOL质粒同源)系统来协调上游降解基因的调控。

图15-17 设想的降解2-氯甲苯途径
当细菌暴露于甲苯或二甲苯以及各种结构类似物(包括2-氯甲苯)时,XylR能激活来自Pu(对tol)的转录。但来自P.putida F1的tod基因则受制于二种调控成分,主要的效应物不够清楚,为了克服这种不确切性,我们设置由XylR和2-氯甲苯单一控制tod和xyl基因簇转录的系统(图15-18),实现这一设想的方法是把tol的基因簇和它的自然启动子工程化成一个单一的NotⅠDNA片段,插入到一个带有Telr的Tn5转座子上,同样tod基因簇被集合作为一个NotⅠ片段,所有部分构成一个mini-Tn5[TOD2]并插入载体,并在Pu启动子和同源的反应甲苯(toluene-responsive)调控物(regulater)XylR调控下表达。
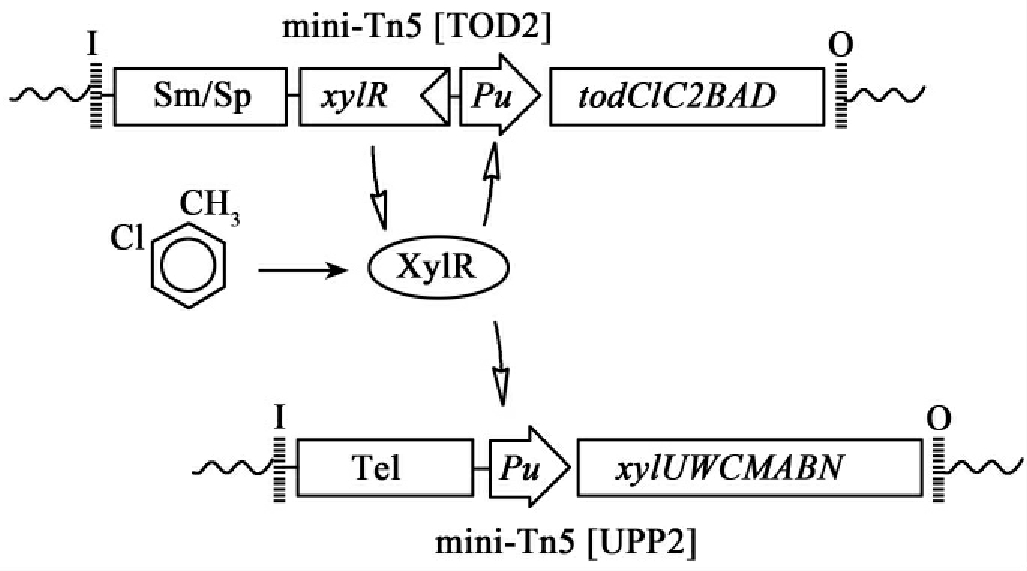
15-18 Xyl R和2-氯甲苯控制的tod和xyl基因簇转录的系统
3)携带TOL和TOD(途径)基因的铜绿假单胞菌(P.aeruginosa)的构建及对氯代烃底物的利用能力。
带有上游TOL和TOD(途径)基因的两个可传递质粒按上述所介绍的方法构建出来。质粒pUPP2带有杂交转座子mini-Tn5(UPP2)(其带有上游TOL(途径)基因,基因由Pu启动子驱动),Pu启动子邻近选择亚碲酸盐(tellurite)抗性标记。质粒ptod2带有mini-Tn5(其带有上游TOD(途径)基因,基因由Pu启动子驱动),启动子邻近xyⅠR基因(图15-19)。这二种质粒通过三亲本融合方法被插入到2-氯苯酸盐降解菌株铜绿假单胞菌PA142和JB2的染色体,从而构建出带有TOL和TOD(途径)基因的铜绿假单胞菌。带有所希望插入的接合后体(exconjugants)在以2-氯苯酸盐为唯一碳源、含有Sm、Sp、Tel的无机培养基平板上选择,具有所希望表型的菌落(生长在2-氯苯酸盐、telr、Smr/Spr)然后被检查对pip(piperacilline)的敏感性。带有mini-Tn5(TOD2)和mini-Tn5(Upp2)的菌株PA142被命名为P.aeruginosa AH001,而带有这二个DNA片段的JB2则被命名为AH002。每个可转移片段的转化频率为10-6~10-7。这两个菌株都不能生长在以2-氯甲苯为唯一碳源的平板或液体培养基中,但当甲苯以气态方式提供时两个菌株均可生长,这说明新导入的TOL和TOD(途径)基因已被激活。不能生长在氯代基质可能意味着三种单个的代谢板块(blocks)活性部分或全部缺失,或不能有效耦合它们的表现。
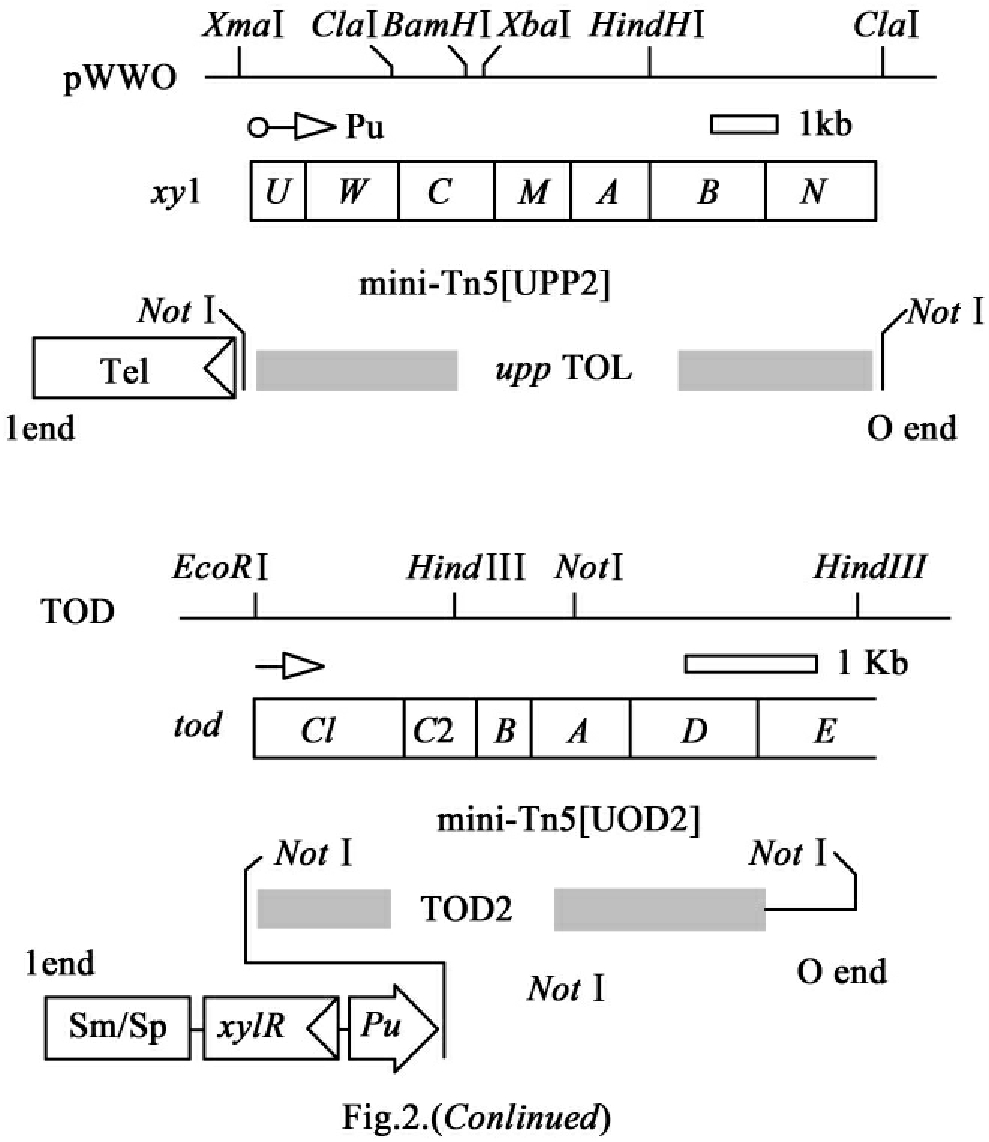
图15-19 编码TOL和TOD途径活性的DNA片段的设计
4)AH001和AH002在存在柠檬酸盐条件下利用2-氯甲苯的能力。工程菌株(AH001和AH002)不能利用2-氯甲苯作为唯一碳源生长。可检验它们在存在柠檬酸盐条件下利用2-氯甲苯的能力。菌株在含有0.2%柠檬酸盐无机培养基上培养2天,培养物的吸光率(absorbance)从0.05上升至0.5,并达到稳定期早期。此后培养物再稀释并暴露在饱和2-氯甲苯条件下又培养4天,但未见培养物吸光率提高,此后再对培养物酸化抽提作GC-MS分析。结果表明重组进入菌株的TOD和TOL(途径)基因得到表达,中间代谢产物中包括氯苯醇和2-氯苯酸盐,但产生的2-氯苯酸盐显然不能支持菌株生长。而非重组的母体菌株不具有这种能力。
5)2-氯甲苯降解工程菌构建研究的启示。按照可信的理论依据构建出的降解2-氯甲苯的遗传工程菌不能利用这种基质作为唯一的碳源、能源生长。而当细胞生长在另一种碳源(如柠檬酸盐)上时,它们明显具有转化2-氯甲苯经2-氯苯醇、2-氯苯醛途径成2-氯苯酸盐的能力,这确证了杂交子上游途径的表达。这些结果说明,它们对2-氯甲苯上游的降解转化能力还不能与下游的2-氯苯酸盐利用能力相耦合匹配,以充分供应碳源、能源使菌株成功生长增殖。
产生上述结果的原因可能的因素:
①2-氯甲苯降解转化中的中间代谢产物可能成为TOD、TOL途径中的非特异性基质,从而产生截止式的羟基化产物(dead-end hydroxylated product),这些产物反过来又损害降解途径中的酶。
②菌株对氯代芳香烃(此处为2-氯甲苯)的降解是酶促的,但生理上是不利的,就是说在能量上这种功能上降解2-氯甲苯消耗和代谢回报(return)是不平衡的。而工程化一种酶或一个酶系列仅能提供生物催化而不能解决降解中这种不平衡的瓶颈。因此用另外一种能源来补偿降解过程缺乏代谢回报(metabolic return)是必需的。这种概念是新的植物修复和根际修复战略的基础。在这些修复过程中,细菌催化者主要依赖植物根排泄物生活,而表达一个或更多基因编码的生物降解活性。
③特定效应的启动子常常在一般生理信号的支配控制之下,导入基因的表达必须和细胞的一般生理学相耦合。强力的生理信号要有利于特定基因的表达和打开某些碳源的代谢。这样实际上我们可以用恒化器(chemostats)和非同源生长培养基来选择菌株的变异性,也可以用来选择释放到环境和生物修复的菌株。
(3)同源重组法构建多功能农药降解基因工程菌
农业生产中农药的大量使用造成污染环境的复杂多样,降解单一农药的微生物菌株已经不能满足生物修复的需求,通过生物学手段构建多功能的农药降解型基因工程菌有重要理论研究及应用价值。
1)工程菌的构建。蒋建东等通过同源重组法将甲基对硫磷水解酶基因(mpd)整合到呋喃丹降解菌鞘氨醇单胞菌CDS-1(Sphingomonas sp.CDS-1)的染色体上,构建出能同时降解甲基对硫磷和呋喃丹又不带抗性标记的重组菌株。构建的具体过程是先构建同源重组载体,继而进行二亲结合,筛选阳性同源重组双交换子(基因工程菌)。具体过程如下:
①以PDT3(含甲基对硫磷水解酶基因mpd)为模板,以引物F1和R1扩增出含有自身启动子的甲基对硫磷(MP)水解酶mpd基因,两端分别引入Nhe I酶切位点和终止密码子。
②用通用引物F27和R1492从染色体DNA扩增出1.5kb左右的16S rDNA与pMD18-T载体连接,挑取得到含有一个NheⅠ单酶切位点的阳性克隆,命名为pMDR。
③把上述①的mpd基因片段插入到pMDR的NheⅠ单酶切位点,获得分别有1个mpd基因和2个mpd基因顺向串联插入的阳性转化子,命名为pMRP-1和pMRP-T。
④用PstⅠ和XhoⅠ双酶切pMRP-I,将2.6kb左右的16S rDNA和mpd片段酶连到同样酶切的pBluescriptⅡsk(++-)载体上,获得阳性转化子pBRP,再用xhoⅠ和Bam HⅠ双酶切,最终将16S rDNA和mpd序列连到pWM91上,转化E.coli DH5αλpir获得pWSM。
⑤用BamHⅠ和NdeⅠ酶切pIB279(Kmr,with Km-sacB fragment),回收3.8kb的Km-sacB片段,酶连到pWSM的BamHI位点,获得pWSMK-1。pWSMK-T的构建类似于pWSMK-1。但其NheⅠ位点间含有2个mpd基因。
⑥将得到的同源重组载体(PWSMK-1)转入E.coli SM10λpir作为供体菌用于二亲结合。
⑦二亲结合筛选阳性同源重组双交换子,将供、受体菌(Sphingomonas sp.CDS-1)培养到对数期后期,收集菌株,并以无菌水洗涤,以不含抗生素的LB培养基培养,再收集洗涤菌体后直接涂布于抗性(Ampr、Kmr、Strr)的选择培养基上,在三抗平板上生长出来能水解MP,产生黄色水解圈的即为同源重组单交换子。单交换子再涂布于添加Str和MP的SLB平板(添加5%蔗糖的LB培养基)上,挑取能产生黄色水解圈的单菌落即为同源重组双交换子。双交换子是仅有mpd基因不含抗性和sacB整合到染色体的同源重组菌。CDS-1染色体rDNA位点插入1个和2个mpd基因的双交换子(工程菌)分别命名为CDS-mpd和CDS-2mpd。
2)工程菌的生长及对MP和呋喃丹降解能力。野生受体菌和工程菌CDS-mpd和CDS-2mpd的比较研究结果:①受体菌和工程菌在LB培养基具有同样的生长能力,这说明mpd基因的插入对受体菌的生长未产生影响。②受体菌保持对呋喃丹的降解能力,但不能降解MP,几乎检测不到降解代谢产物PNP,而工程菌CDS-mpd和CDS-2mpd能快速(1~2h内)水解MP成PNP,而呋喃丹前期降解较慢,但随着工程菌的生长,呋喃丹在30h时被完全降解。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









