



第一节 生命的起源
生命为什么能在地球上发生?我们现在的地球是否还会发生类似的过程?生命起源前,地球上是如何形成生命物质的?这些都是有关生命起源的关键问题。
一、生命起源的地球条件及其演变
200亿年前宇宙形成于“大爆炸”(The big bang),同时产生了碳、氢、氧、氮、磷、硫等构成生命的基本元素。46亿年前,太阳和地球初步形成,构建好了生命产生的场所。地球形成之初,主要是由氢、氦及大爆炸产生的星际尘埃组成,其温度仅约10K。随着星际尘埃凝聚收缩形成地球的内核,产生大量的热量,使得地球表面温度逐渐升高,加上强劲的太阳风,使地球表层以氢和氦为主的初生大气烟消云散。随着原始的地壳形成,地球表面的温度也逐渐下降,但地球内部的温度依旧很高,其形成的频繁的火山喷发,以及小行星撞击地球产生的高能量使地球表面熔化,地球内部物质分解产生的大气也随之释放,形成了地球表面的第二次大气层,也称为原始大气,或次生大气。原始大气的成分主要是具还原性的二氧化碳、甲烷、氮气、水蒸气、硫化氢和氨气等,大气中不含游离氧,以氧化物形式存在。原始大气的形成为生命的起源提供了素材。
据现有资料表明,地球生命起源于海洋,而地球形成之初没有液态水,水仅以气态的水蒸气存在。所以,地球上液态水的出现是生命的化学演变之重要转折点,也是生命起源的必要准备。随着地壳的隆起与凹陷,形成了低地、山谷、洼地,当地球气温降低,空气水分增多后,水蒸气变成雨水下落,在低洼处积聚,最后汇集至全球最低的地方,形成原始的海洋。同时,一些可溶性的化合物也溶解到水中。因此,原始海洋中积累了许多化合物,包括最原始的有机化合物——甲烷。
地球已经形成了生命起源所需的原材料和场所,只要有适当的能量驱使,即可形成原始生命。一般认为,原始地球上可利用的能量主要有以下几个来源:热能、太阳能、放电、宇宙射线等。
地球形成之初,宇宙充满星际尘埃,太阳射线到达地球的量有限,化学演变所需的能量主要来自地球凝聚过程和气体逸散散发的能量,也即地球自身释放的巨大热能。太阳系内的星际尘埃逐渐消失后,含有大量紫外线、可见光、电子、质子和X射线的太阳光大量地投射到地球上,大大地促进了原始生命物质的形成。另外,火山活动过程中,高温气体喷射到空气中,从而产生高强度电流的雷电也促进了原始生命物质的形成。比如,雷电非常有利于促进原始地球的氰化物(HCN)形成,氰化物形成在生命的化学演变中起着重要的作用。
从上可知,初形成时的地球与现在的地球环境条件差别很大,只有在当时的条件下生命才会出现。生命起源的条件主要包括:
1.必需要有生命起源所需的元素材料,主要包括碳、氢、氧、氮等;
2.必需要有液态水的出现与聚集,水是地球生物不可或缺的物质;
3.一定的能量,主要包括地球热能、太阳能、闪电、宇宙射线等。
二、生命物质的形成与演变
生命发生的最早阶段是化学进化,即从无机小分子发展到细胞诞生的阶段,也称之为生命化学阶段。这一阶段主要包括简单的生物分子的形成、生物小分子聚合成生物大分子,如氨基酸、嘌呤、嘧啶及核苷酸等的形成,以及氨基酸聚合成蛋白质、核苷酸聚合成核酸等。根据化石及岩石中碳同位素分析推测,一般认为距今37~38亿年前可能为细胞出现之前的生命化学进化,而之后为细胞继续进化的生物进化阶段。
生命的化学进化大致经历了以下四个连续的阶段:
1.从无机小分子生成有机小分子
约在47亿年前,地球还处于星云凝聚阶段时,与生命起源有关的碳、氧、氮、氢、磷和硫等元素就开始了第一次的化学反应,主要合成氨气、甲烷、二氧化碳、水等原始大气的成分。地球形成之后,开始了第二次的化学反应,主要合成一些简单的有机物,如氨基酸、嘌呤、嘧啶、有机酸等。
一旦关于生命发生的物理化学环境信息变得清晰,下一步就是实验室重演一个生命起源的早期阶段发生的过程,目前已有大量的实验室模拟了这一过程。第一个实验模拟是1953年芝加哥大学的米勒(Miller SL)进行的“米勒-尤里实验”,他利用其设计的装置(图7-1)模拟原始地球的条件下的无机小分子生成有机小分子的过程。他将整个装置内部抽成真空,加入200ml水,泵入10cm、20cm、20cm汞柱的氢气、甲烷和氨气形成的混合气体。对水持续加热沸腾产生水蒸气,并将这些气体暴露在连续放电之下。然后,将反应的水溶性产物通过冷凝器冷凝后进行检测。反应连续进行一周后,得到了多种氨基酸和有机酸,包括甘氨酸、丙氨酸、天冬氨酸等蛋白质中含量最多的几种氨基酸。这些氨基酸与1969年在澳洲发现的默奇森(Murchison)陨石上找到的氨基酸的种类和比例相当,这使得米勒-尤里模拟实验更令人信服。
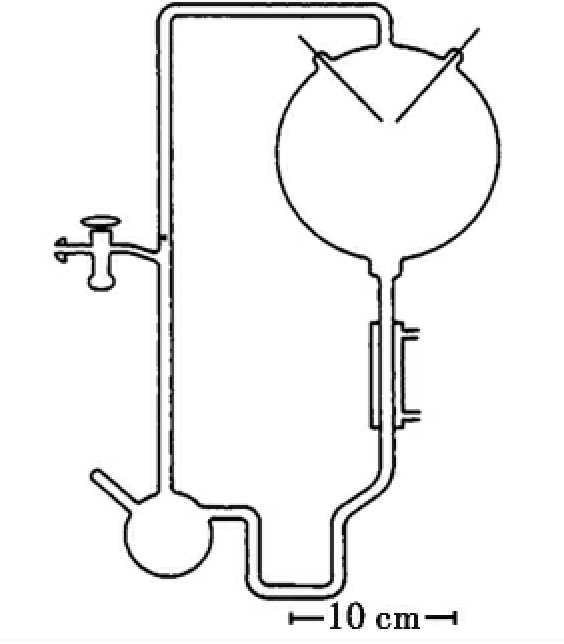
图7-1 米勒-尤里实验装置示意图
此后,大量的学者进行了类似的模拟实验,比如1960年休斯敦大学的奥罗(Oro J)利用氰化氢和氨气溶液合成了腺嘌呤;1963年又以甲醛为原料,在紫外线照射下合成了核糖和脱氧核糖。1986年北京大学教授王文清以甲烷、氮、硫化氢、氨气和水为原料,采用米勒装置进行试验,最后得到14种氨基酸。这些结果证明,所有构成生物体的氨基酸、碱基等均可在原始地球条件下经多种途径产生。
2.从有机小分子生成生物大分子
蛋白质和核酸是生命的两大基石,在原始地球条件下,这两大物质如何形成?可能实现的途径有哪些?这是生命起源的一个关键问题。
关于蛋白质和核酸的起源,目前的普遍共识就是:氨基酸、核苷酸在海水中经过长期的积累、相互作用,在适当条件下,分别通过缩合反应或聚合作用,形成了原始的蛋白质和核酸分子。关于蛋白质和核酸的起源地点和条件,目前有三大不同的学说。
(1)陆相起源说。该学说认为,大陆火山附近的水池中生成了大量的氨基酸、核苷酸,火山喷发使水池被高温蒸干,氨基酸、核苷酸在高温下发生热聚合反应,脱水形成高聚物,然后经雨水冲洗进入原始海洋,高聚物进行自我装配形成蛋白质或核酸。其核心观点是大陆无氧干燥环境是脱水的理想场所。1955年,美国学者福克斯(Fox SW)等将多种氨基酸氨比例混合,在无水条件下加热到160~200℃,经过0.5~3小时,合成出一种具水解、脱羧、胺化和脱氨基等类似酶活性的高聚物——类蛋白质。这一模拟实验及类似的实验结果支持了这一假说。
(2)海相起源说。该学说认为,相对分子量小的氨基酸和核苷酸能够被吸附在粘土、蒙脱石一类物质的活性表面,在适当的缩合剂的存在下,氨基酸、核苷酸可以发生脱水,缩合成大分子量的高聚物,高聚物积累后可进行自我装配形成蛋白质和核酸。持这一假说的霍洛维茨(Horowitz MP)在1967年进行了模拟实验:他在甘氨酸和ATP的水溶液中加入沸石,缩合生成了甘氨酰腺嘌呤核苷酸-磷酸,这种高能键化合物在弱碱性条件下被吸附到蒙脱石的活性表面,经聚合产生类蛋白物质——多聚甘氨酸。
(3)深海热泉起源说。该学说认为,海水与洋嵴下的岩浆体可以进行物质和能量交换,与海底烟囱喷出各种气体、金属和非金属,如甲烷、氢气、氦、硫化氢、铁、锰、锌、铜、硅等,沿着烟囱从岩浆到烟囱口形成了从1000℃至350℃的温度梯度变化,烟囱口与周围海水经热交换又形成了从350℃至0℃的温度梯度变化。沿着这两个温度变化梯度,形成了一个连续的化学反应渐变区,提供了满足各类化学反应的条件。岩浆中喷出的氢气、甲烷、氨气、硫化氢等经高温合成氨基酸、核苷酸,继而形成含磷、硫的复杂化合物,进一步形成多肽、多核苷酸链,最后形成类似蛋白质、核酸的化合物。该学说由华盛顿大学的巴罗斯(Baross JA)及其同事在1985年提出,并且得到加利福尼亚大学洛杉矶分校的莱克(Lake J)及北京大学张昀、李江海等人的支持,也说明这一假说有一定合理性。
以上三种说法各有其合理之处,同时也各自有实验证据给予支持。因此,也有人试图综合这三种学说提出“冰火反应器”的学说,认为原始地球上存在可以保持冰水混合物存在,又邻近火山及海中热液口的潮汐池塘,蛋白质和核酸在这里进行组装。在没有找到合理假说完美解释蛋白质和核酸的合成与装配之前,这些假说都认为是合理可信的。同时也说明,只要有适合化学进化进行的条件存在,生命的出现就是可能的。
3.由生物大分子组成多分子体系
单个生物大分子单独存在时不表现生命现象,不是原始生命,只有在它们形成多分子体系时才能显示出生命现象。生物大分子如何组装成多分子体系?它们需要在什么条件下才能组装?这是生物大分子合成后,紧接着要解答的问题。目前有两个经典的实验模型可给人们提供一些思路。
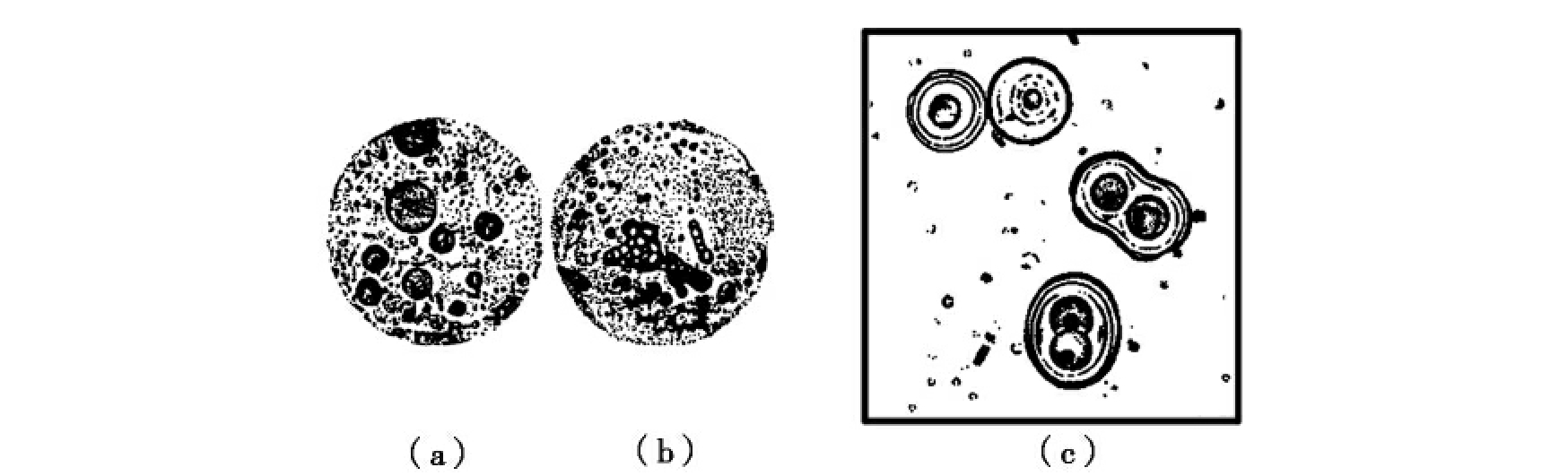
图7-2 两个多分子体系模型图
(a)团聚体模型;(b)团聚体模型的生长与分裂;(c)微球体模型的生长与分裂。
一个是前苏联生物化学家奥巴林(Oparin AV)提出的团聚体模型(coacervate theory),该模型认为蛋白质溶液与核酸溶液混合时,可形成团聚体小滴,从而构成多分子体系,这种多分子体系的小滴具有一定的生命迹象。该模式是由奥巴林在1953年进行实验后提出,他将透明的白明胶(蛋白质)水溶液与阿拉伯胶(糖)水溶液混合后发现,溶液变混浊,在显微镜下可观察到溶液中出现了小滴,即团聚体,团聚体与水之间有明显的界限。随后,他分别混合蛋白质、核酸、多糖、磷脂及多肽等溶液也会形成类似的团聚体,并且他还能使团聚体小滴具有原始的增殖与代谢特性,并使之稳定存在几个小时到几周。
另一个是福克斯(Fox SW)提出的微球体模型(microsphere theory),他将干的氨基酸或合成的类蛋白质加热浓缩,可形成相当于细菌大小的微球体,这种微球体在溶液中表现稳定。同时,微球体还表现出了一定的生物学特性,如表面有双层膜、在盐溶液中会缩小、能吸收溶液中的类蛋白质而生长和繁殖、在吸收ATP后表现出类似细胞质流的活动等。福克斯认为类蛋白微球体是一种比较理想的多分子体系,并认为这是生物细胞的前体。
不管是团聚体还是微球体模型都有各自的合理之处及缺陷,如团聚体模型所采用的实验原材料为现代生物细胞的产物,而类蛋白微球体的组成中没有核酸。然而,这些模型都说明生物大分子是自动聚集从而形成独立的多分子体系的。
4.由多分子系统形成原始生命活动
多分子体系虽然显示出些许生命现象,但是没有严格意义上的生命活动,只能算是生命的萌芽。由多分子体系发展到原始生命,是生命起源过程中最复杂和最具决定意义的阶段,这一过程目前还不能进行实验室模拟。一般认为,这一过程的发生,还有以下三个重要问题需要解决:细胞膜的形成、开放系统的形成和遗传密码的形成。
(1)细胞膜的起源
生命是一个与环境很不相同的独立体系,细胞与周围环境有清晰、明显的界限,将其内部与外部环境隔开。将细胞内外相隔的结构是细胞膜,所以生命起源首要解决的一个问题就是细胞膜的形成。细胞膜是具有选择通透功能的薄膜,是一种嵌有蛋白质的类脂双层膜结构。目前关于细胞膜起源的问题还不是很清楚。最早有人认为是随着生物大分子浓缩成多分子体系的过程中而产生的,如团聚体模型、类蛋白微球体与周围环境形成的界限。戈尔达克尔(Goldacre RJ)提出的类脂蛋白质囊假说认为原始海洋中存在类脂和蛋白质,并且它们之间相互作用形成囊状,把富含有机物的海水包裹,使之形成独立于外界环境的原始生命体,如磷脂分子由亲水的磷脂酰碱基头部、两条疏水的甘油基团尾部组成,在水中易形成微团、双分子层(如图7-3),蛋白质分子也很容易嵌入形成细胞膜。但也有人认为,磷脂分子结构太复杂,在前生物条件下不能大量地合成,脂肪酸才是前生物条件下大量存在、可利用且能够自我装配成囊泡的有机物质。在不同pH条件下,脂肪酸可自我装配成囊泡或微团(如图7-4)。高pH时可聚集成微团,低pH时分离成油滴,中等pH时(通常为7.5~9.5)形成双分子层的囊泡。不管细胞膜怎么形成,原始膜的形成在生命起源中起着奠基作用,标志着原始细胞的诞生。
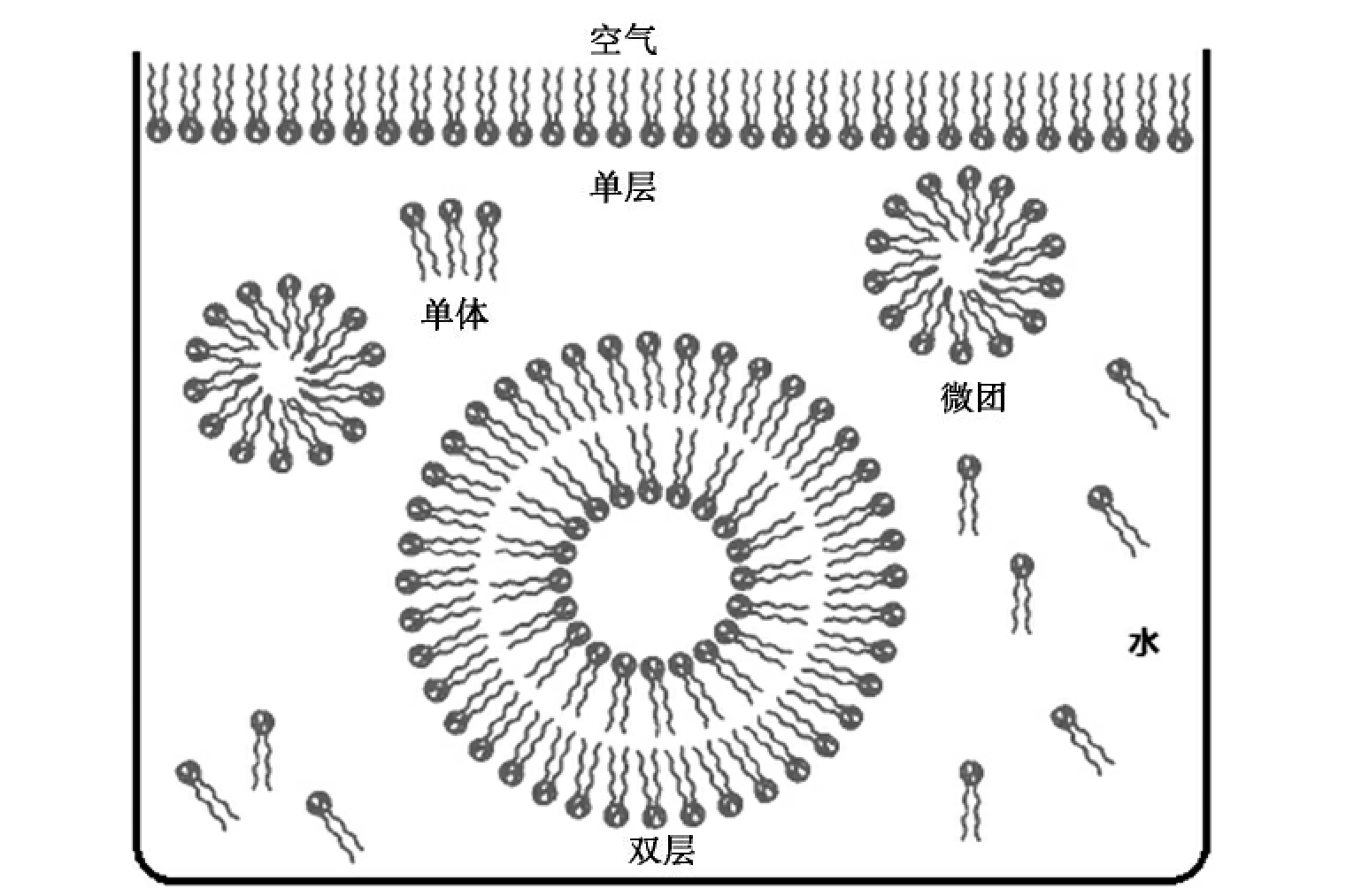
图7-3 磷脂分子在水溶液中的结构
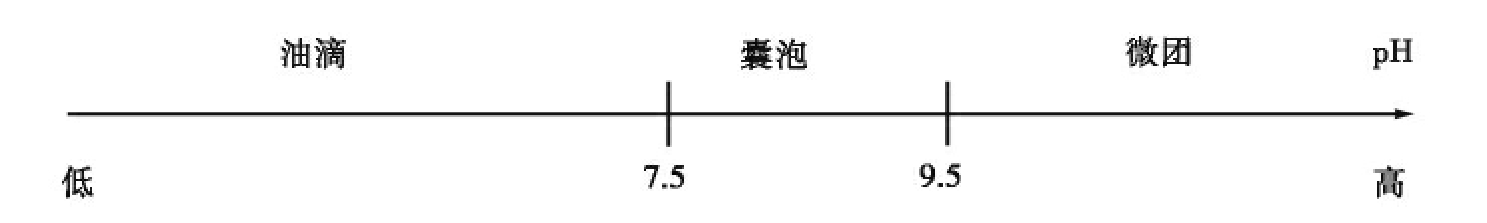
图7-4 脂肪酸的自我装配过程
(2)开放系统的建立
生命现象的本质特征在于不断与环境进行物质和能量的交换,因此,原始生命必然是一个开放系统。奥巴林曾经利用组蛋白和核苷酸构建的团聚体进行研究,他将可以催化葡萄糖-1-磷酸盐合成淀粉的葡萄糖磷酸转化酶和催化淀粉水解为麦芽糖的β-淀粉酶加入到团聚体内,并在团聚体周围加入葡萄糖-1-磷酸盐,结果在团聚体外测到麦芽糖,说明此时的团聚体与周围环境进行了物质和能量的交换(如图7-5)。
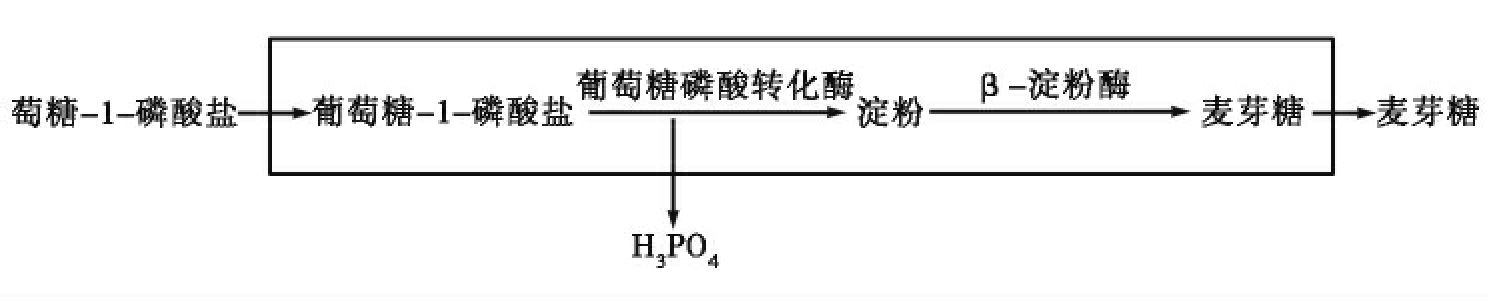
图7-5 奥巴林的团聚体实验示意图
(3)遗传密码的起源
生物体含有非常大的信息量,这些信息主要由DNA中的多聚序列携带,可通过转录和翻译被转化成RNA和蛋白质。核酸与蛋白质之间的信息传递是生命现象的另一个重要特征,也是生命起源的另一关键问题。
关于核酸与蛋白质的信息传递系统如何形成,也即遗传密码如何起源,目前有两类盛行的假说。一个是克里克(Crick FHC)在1968年提出的偶然冻结理论(The acsident frozen theory),认为三联体密码子与相对应的氨基酸的密码关系完全是偶然的,而这种关系一旦建立就立即冻结保持不变。另一个是韦斯(Woese CR)在1966年提出的立体化学理论(stereochemical theory),认为三联体密码子与相对应的氨基酸的密码关系起源于它们之间特殊的立体化学相互作用,RNA与氨基酸之间的化学作用及其物理和化学互补性促使遗传密码子的形成。但是这两个假说都未得到很好的实验验证,无法令人信服。有关遗传密码起源问题的研究还在继续。
遗传密码最初如何建立?过去也有许多的设想,有人认为遗传密码一开始是单体,后来变为二体密码,最后进化成三联体密码。也有人认为一开始是二体密码,继而进化成三体(联)密码。这两种设想都面临一个问题:当密码子从单体向二体进化,或从二体向三体进化时,原有的密码系统都完全改变,这种改变不利于遗传信息的积累,也不符合生物进化的规律。所以,目前广泛接受的就是,遗传密码子一开始就是三联体密码。
三、原始细胞的进化和原核细胞的出现
地球上第一个原始细胞的形成标志着生命的化学进化的完成和生物进化的开始。据化石记录推测,37~38亿年前就已经形成了原始细胞,而且从最早细胞生命的出现到原核细胞形成的生物进化,经历了太古宙和元古宙两个地质时期。
原始的原核细胞是地球上最早的生命实体,但原始细胞与原核细胞之间还有一段漫长的细胞进化过程。这一过程主要包括细胞质的进化、原始染色质形成、细胞质内不同酶系统集中到一定的区域、细胞的代谢趋于有序化,并且随着DNA大分子的进化,原始的染色体也逐渐形成。
关于原核细胞起源的大致历程的论述,有很多学者提出了各种各样的假说,以下介绍两种最具代表性的假说。
1.超循环组织模式
上世纪40年代奥地利生物学家贝塔朗菲(Beretalanffy LV)提出生命是具有整体性、动态性和开放性的有序系统,开创了以系统论的方法研究生命的新思路,改变了过去分割、孤立式的传统研究方法。
随着以自我组织模式的系统论方法在生命起源研究中的运用,德国学者(Eigen M)认为生命起源的化学进化和生物进化之间存在一个分子自我组织阶段,通过生物大分子的自我组织建立起超循环组织并过渡到原始的细胞结构生命,即原核细胞起源的超循环组织模式(hypercyclic organization)。这个超循环组织是由自催化或自我复制的单元组织起来的超级循环系统,这个超级循环系统能自我复制而保持和积累遗传信息,并且复制中可能出错而产生变异,从而将这个超循环组织系统纳入达尔文的进化模式。这个超循环组织系统也称为分子达尔文系统。
2.阶梯式过渡模式
奥地利学者休斯特(Schuster P)等人根据超循环组织模式,发展出了一个综合的过渡理论,即阶段式过渡模式(steps translation),这是一个包括6个阶段式步骤的由小分子进化到细胞生命的理论(图7-6)。

图7-6 从无机小分子到生物进化的阶段式过渡模式图
第一步是小分子物质通过聚合作用形成异聚体,这一步主要面临着组织化危机,即假如分散的、无组织的小分子不能初步自我组织,将不能进入下一阶段的进化。第二步是从异聚体通过选择作用形成多核苷酸,这一步主要面临的是复杂化危机。第三步是多核苷酸和多肽等物质进一步自我组织形成分子准种(quasispecies),这一步主要是通过突变与选择进行优化组合,面临着适应危机。第四步是分子准种形成超级循环组织,主要通过功能性组织翻译进行,这一步面临的是信息危机,即最早的基因和遗传密码的产生问题。第五步是超级循环组织分化成分隔结构,主要是原始细胞内的分区,这一阶段面临的是基因型与表现型的分歧。第六步是分隔结构进一步进化成原核生物,在一系列多核苷酸基因系统控制下的代谢反应提供了多核苷酸复制与蛋白质合成等生命活动所需的能量。
四、真核细胞的起源
真核细胞的出现是继原核细胞形成之后的又一重大进化事件,它的出现对以后的生物进化极为重要。根据化石记录,大量的真核生物化石出现在元古宙晚期,即大约8亿~10亿年前。蓝细菌的大量繁衍,其光合作用使得大气圈中氧气积累、二氧化碳含量下降,以及地球表面的物理、化学性质变化,为真核生命的出现创造了条件。
1.三分支进化与真核细胞的始祖
近年的细胞生物学和分子生物学研究表明,原核生物并不是统一的一大类,原始的原核细胞在极早的时候可能已经分化为两大类:真细菌和古细菌。真细菌包括绝大部分我们所认识的原核生物,如细菌、蓝藻、放线菌、螺旋体、衣原体和支原体等。而古细菌主要包括一些生长在地球特殊环境中的微生物,包括产甲烷细菌类、盐细菌、热原质体、硫氧化菌等,这是一类与人类生活无直接关系的原核生物。美国伊利诺斯大学的微生物学家沃斯(Woese CR)根据原核细胞与真核细胞的分子结构与序列提出了早期生物的三分支进化的观点,认为古细菌、真细菌和真核生物都起源于一个共同祖先(图7-7)。
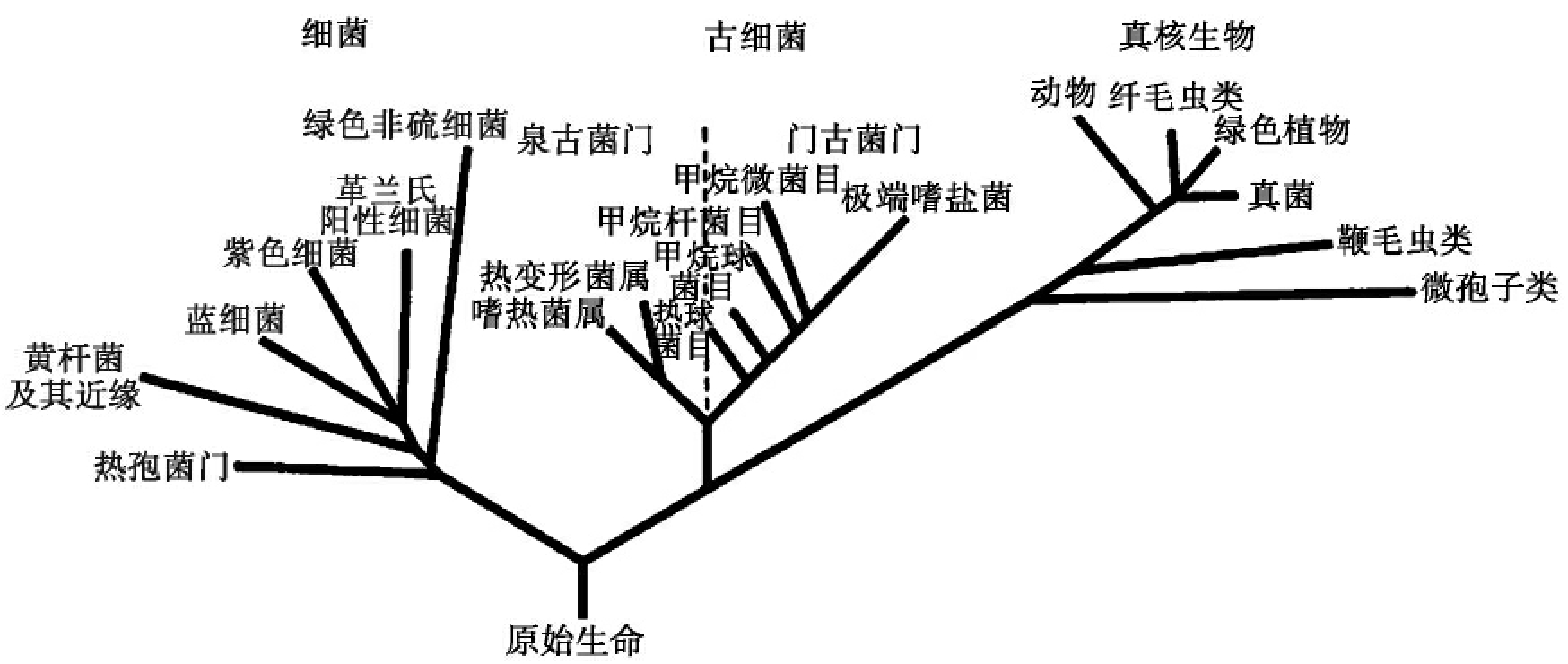
图7-7 Woese的三分支学说示意图
2.真核细胞起源的途径
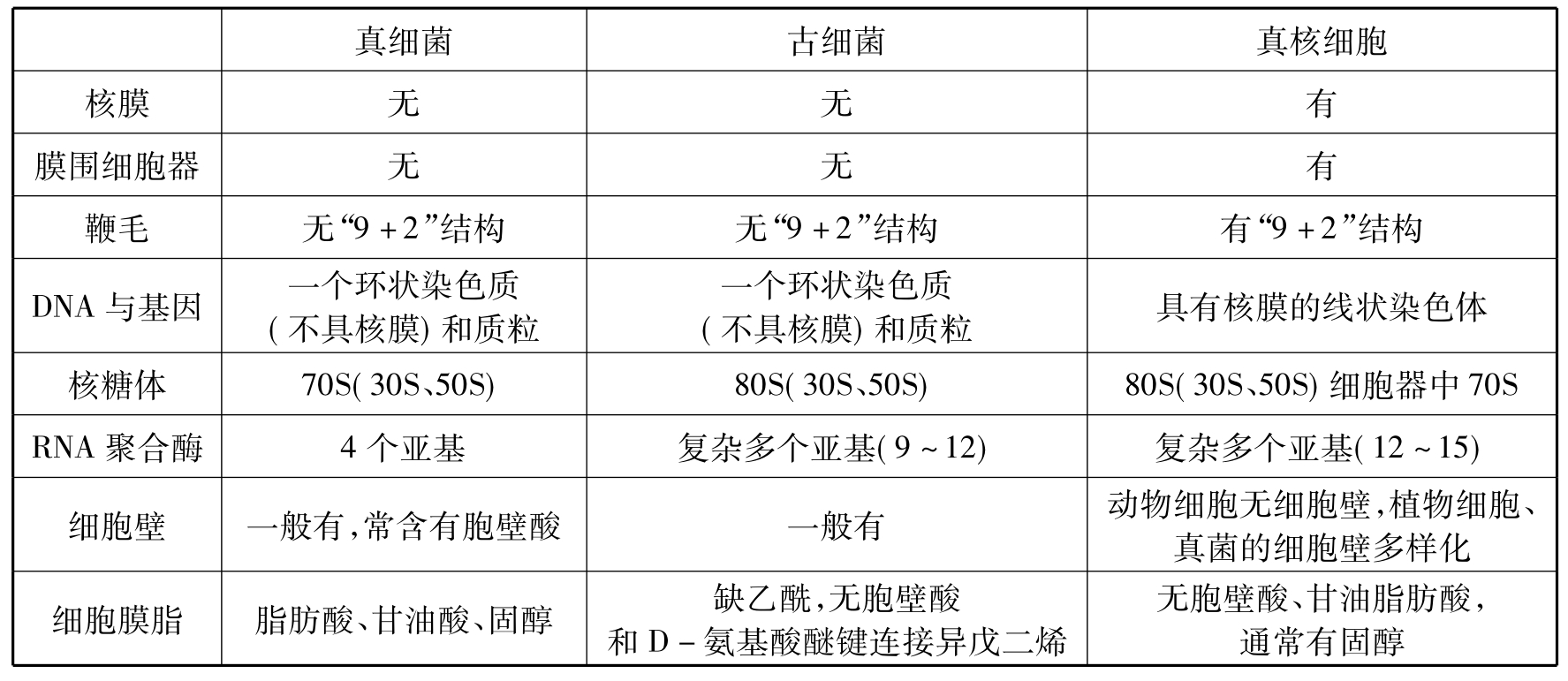
原核细胞与真核细胞在结构上差别巨大。从表7-1中可以看出,这些差别主要表现在细胞器的有无和遗传物质的存在形式与结构。真核细胞的起源,主要是细胞器的起源与细胞核的形成。目前有两种相对立的学说:内共生起源说和渐进式进化学说,推测真核细胞的起源。
表7-1 真核细胞、真细菌和古细菌的生物特征比较
(1)内共生起源说
内共生起源说(endosymbiont hypothesis)由美国波士顿大学生物学家玛格利斯(Margulis L)提出,认为真核细胞是一个复合体,原核细胞才是最小的细胞单位。真核细胞的线粒体和叶绿体来源于共生的真细菌,线粒体可能来源于紫细菌,叶绿体可能来源于蓝细菌,运动器(包括鞭毛和胞内微管系统)来自共生的螺旋体类真细菌。这一假说有许多方面的证据支持:
①线粒体膜的形态结构与细菌相似,叶绿体的膜形态结构与蓝细菌的相似。线粒体和叶绿体内外膜的化学成分与物理特性不一致,外膜与真核细胞的质膜相似,而内膜与原核细胞的质膜相似。
②线粒体和叶绿体都是半自主性细胞器,可以通过直接分裂的方式繁殖,这说明它们曾经是独立生存的,且它们裸露的环状DNA分子也与原核生物相似。
③线粒体和叶绿体的核糖体在大小和对蛋白质合成抑制剂的反应性质上与原核生物相似。
④对16s rRNA序列分析比较,红藻的叶绿体来源于蓝细菌,对tRNA序列的分析也得到类似结果。
⑤同工酶与代谢途径研究的证据也支持这一假说。
⑥现在的真核细胞中,有些种类依然存在内共生现象,如有些蓝细菌是变形虫、硅藻、鞭毛原生生物和无叶绿体的绿藻的内共生体。
(2)渐进式演化学说
渐进式说由尤泽尔(Uzzell T)和斯波尔斯基(Spolsky C)在1974年提出,用吞噬作用、内泡形成和细胞内间隔作用的渐进发展来揭示细胞核与细胞器的起源,认为真核细胞的细胞器和细胞核的形成是由原始的原核细胞通过一系列DNA复制及质膜的内陷形成。其主张从原核细胞到真核细胞是直接的、渐进的进化过程。而后,根据这一学说,进一步提出膜进化理论(membrane evolution theory),即用膜分化导致代谢分隔来解释细胞器和细胞核的起源。这一假说也得到一些证据支持:
①原核细胞与真核细胞之间存在一些中间过渡类型,如原绿藻没有细胞核但色素组成与绿藻相似,蓝细菌细胞内存在囊泡系统,色素成分与某些植物类似,可以进行释放氧气的光合作用。
②从代谢的生理和生化特征比较看来,真核细胞的需氧呼吸代谢更可能是通过原核细胞发酵代谢途径的重复、改造而来。
③某些进行光合作用的原核生物具有复杂的胞内膜结构。
这两个学说各有其缺陷:内共生起源学说不能很好地解释细胞核的起源,而渐进式演化学说则对线粒体和叶绿体内外膜的化学成分和物理特性不一的事实解释不清。虽然对这两个学说仍有争论,但是越来越多的分子系统证据倾向于支持真核细胞内共生起源学说。
比利时学者德迪韦(de Duve CR)综合真核细胞起源的研究成果,对真核细胞起源的大致历程提出了一个看法:a.大约在20亿年前,大气中氧气含量增加,某些原核生物除去了细胞壁而成具有双层质膜的细胞,且其内膜附有核糖体;b.质膜不断内陷,扩大了细胞的表面积; c.质膜的进一步内陷而与细胞膜脱离,进入细胞质内形成“胞内小泡”,且外膜附有核糖体;d.部分“胞内小泡”围绕DNA形成的“拟核”排列,而后发展成有孔的核膜,“拟核”从而进化成为细胞核;e.通过吞噬作用和内共生作用形成过氧化物酶体、溶酶体、线粒体、叶绿体等细胞器,从而消除了氧气对细胞的毒害,反而能利用氧产能供细胞的生命活动,与此同时,“胞内小泡”进一步进化成高尔基体和内质网。
3.真核细胞起源的意义
真核细胞的形成是继原核细胞起源之后的又一重大进化事件,它的出现对地球生物进化起着极其重要的作用,其主要意义在于:
(1)为生物的性分化和有性生殖打下基础。有性生殖是真核生物与原核生物的主要区别之一,也是推进生物进化的主要动力。生物进化的30亿年中,无性生物时代占近三分之二的时间,但生命长期处于单细胞阶段,进化极为缓慢。有性生殖出现后,生物进化的速度才明显加快。由两性个体组成种群行有性生殖,个体不再是原核生物的一成不变延续亲本的基因型。在有性生殖下,个体具有两套基因,使基因重组产生新的遗传变异成为可能,从而提高了物种的变异性,并增大了变异量,从而加快了进化的速度。
(2)真核细胞的出现推动了生物向多细胞化方向发展。尽管部分原核生物已有多细胞化倾向,如蓝藻,但原核细胞的遗传系统还不足以实现复杂的细胞间遗传调控,其并没有进化到组织、器官的结构形式,还仅仅是停留在单个细胞或细胞集群的等级上。只有真核细胞才有细胞分化的可能,并最终发展出具有分化的组织和行使特殊功能的器官的复杂多细胞生物,使生物向高级发展。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











