



第二十章 DNA是三重螺旋
克里克在斯特兰奇韦实验室工作时,收到了下面这封信:
亲爱的克里克:
剑桥近况如何?是不是刺骨的寒风正吹遍沼泽地,使剑桥湖水碧波荡漾,墙上铁丝网萧萧作声,校门上链条锁格格作响,打扫宿舍的校工容光满面,而学生们穿过鹅卵石小路匆忙地走向学校的浴室?从斯特兰奇韦实验室门下面吹进的寒风,是不是使培养基凝冻并使两栖动物冬眠?
下次来市区时,请事先给我写个明信片,到时我再打电话给你,约个时间请你来吃饭。最近我学会了烧菜并准备去弄一桶苹果酒。
下次来时,务必先告诉我。
你的莫里斯·威尔金斯
(威尔金斯,1948)
这两位科学家之间的友情,对发现双螺旋起了重要作用。他们是如何相识的呢?虽然威尔金斯出生在新西兰,但是从八岁起他一直住在伯明翰。克里克来自北安普敦,为了上大学,他到了伦敦,而威尔金斯去了剑桥。在二战爆发前,他们就开始了研究工作。当时,克里克在伦敦,威尔金斯在伯明翰。由于战争的需要克里克在英国服役,而威尔金斯前往美国参加了曼哈顿工程。这好比是命里注定他们不会碰头。但是在不同的时候,他们都曾在哈里·马西(现在的哈里爵士)手下工作过。在他们想从事生物学研究时,他们都去征求马西的意见。先是威尔金斯,然后是克里克。马西向克里克建议,应该去见见威尔金斯。克里克回忆说,他“当时,不认识许多生物物理学家。也许我只碰到过兰德尔,但是我同威尔金斯是有私交的……”(克里克,1968/1972)。
沃森第一次碰到威尔金斯是1951年春天,在那不勒斯。当时他想说服威尔金斯一道去游览帕斯特姆的希腊神殿,但是威尔金斯没有发表意见。的确,R.D普雷斯登回忆说,在参观这些神殿时,其中有一个人利用他挡住了沃森搜索的目光(私人通信)。这很可能就是威尔金斯!
沃森与克里克的会见
沃森到剑桥工作时,正好与罗伯特·帕里什和弗朗西斯·克里克合用一间办公室。由于沃森在转到剑桥大学当研究生问题上,曾同保罗·韦斯有过争执,因此他自称是在罗伊·马卡姆手下研究蛋白质和植物病毒结构的,实际上他是在卡文迪什肯德鲁手下试图结晶肌红蛋白,作X线衍射研究。由于这方面的工作没有成功,因此他每天能有几小时的时间同克里克交谈。沃森向德布吕克谈起克里克时,把他说成“无疑是我共事过的最聪明的一个人。而且他的工作方法也是我见到过的、同波林最相似的……他总是在谈话或思考……”(沃森,1951b)。过了好长时间后,沃森说他觉得“克里克知道DNA比蛋白质更重要”(《双螺旋》,48)。但是,克里克并没有那样肯定DNA比蛋白质重要,他评论说:
你问我在我第一次碰到沃森时我对DNA和染色体知道些什么,我只能回答我确实已记不清了……当时,关于DNA我一定早有了解了。因为在斯特兰奇韦实验室工作时,有人叫我做一个报告,当时的题目(或者是我自己决定的)就是关于DNA。那时关于DNA,我知道的还不多。我记得,在报告中我指出了辐射确会以某种奇怪的方式降低黏性,这证明分子大概是长的。显然,当时我一定是很有兴趣的,但我认为,用现在的话来说,我没有相信DNA具有压倒一切的重要性,建议研究DNA而不研究蛋白质。因为,当时我认为蛋白质也是重要的,并正在做这方面的研究。我很怀疑我还能找到这次报告的笔记,从中看出在遇到吉姆·沃森前我是怎样想的。我可以相当肯定的是,这篇报告的主题是生物学的中心问题在于解释基因是怎样复制、怎样作用以及指出基因的主要作用可能是控制蛋白质的合成,特别是蛋白质中氨基酸顺序的形成。可惜,我现在没有文件来证实上述回忆。
(克里克,1968/1972;1973)
为了验证克里克在遇到沃森之前对DNA的看法,我们可看一下他对威尔金斯的研究工作所持的态度。威尔金斯的同事们回顾了当威尔金斯在研究DNA碰到问题时,克里克是如何取笑他的朋友的。一次在河堤公园里吃茶点时,克里克曾说:“你应当做的,是为自己搞到一种适当的蛋白质。”(G.布朗,1969)由于1950年10月到1951年9月,杰弗里·布朗正在斯德哥尔摩,因此我们可以推测到,克里克讲这句话是在1950年夏天,当时大家都认为,对蛋白质晶体学家来说,DNA照片是毫无希望的。
克里克坚信,他参加了在卡文迪什举行的、由佩鲁茨召集的蛋白质讨论会,但他回忆不起威尔金斯在会上是否谈到DNA。如果那是指1950年召开的那次会,那么正如威尔金斯所回忆的,当时从X射线数据中几乎没有得到关于DNA的情况(威尔金斯,1972)。我们还不应忘记,根据威尔金斯的同事——比尔·西兹的说法,在会议过程中,克里克坐在会议厅的后排同别人谈得很有劲(西兹,私人通信)。
我们也不应认为,威尔金斯于1950年6月首次拍摄到赛纳提供的纤维的X光照片时,他确信DNA是遗传物质。那年八月,威尔金斯曾向马卡姆吐露:“当然,我们真正想做的是要找出核酸在细胞中的作用。我很希望这台用于研究活细胞的新紫外线显微镜,能使我们更清楚地知道核酸是在细胞的什么地方(及什么时候)产生的。”(威尔金斯,1950c)翌年春,他信心更足了。他说:“研究活细胞的晶体核蛋白有助于解决基因结构问题。”(1951a,105)沃森在那不勒斯听到这些话之前,他对核酸的态度似乎不太明确。他说他的梦想是:“要发现基因的奥秘,但我从未想到过什么有价值的主意。”(《螺旋》,30)确实,他曾去哥本哈根学习核酸化学。DNA看上去很像是基因的成分,但DNA是不是基因复制的关键还有待于研究。眼前的问题是如何弄清遗传物质的结构。染色体不是好的材料,核蛋白又十分复杂。在那不勒斯时,威尔金斯给沃森讲解了通过研究DNA钠盐的定向纤维的结构,可以了解基因的结构。
关于沃森同克里克在1951年1月讨论的性质,我们能猜测些什么呢?我看有下面这些情况:遗传图的重要特性在于它表示了单位的线状顺序。这些单位又可一直细分到分子水平的亚单位直线顺序。对这样的顺序来说,一定还相应地有一条可能由核苷酸组成的化学顺序。但是,染色体和病毒具有的神秘的自我复制特性又是什么呢?根据卡斯帕森的工作,人们都同意染色体复制少不了DNA,但还不清楚DNA能否不需蛋白质的帮助,而复制自身。现在也没有搞清这个问题。在1951年,沃森和克里克也推论说:不可能从DNA结构中找到这种复制的性质。他们确实可能研究出DNA的结构,但只会发现,基因的复制仍笼罩着神秘的色彩。他们提出的第一个模型和1953年波林的模型就是如此,但接下来的问题是如何来解释这种结构。克里克无疑知道需要什么方法来解决这个问题。他们一定使用了波林在研究α-螺旋上所用的方法。
正如我们已知道的,波林和科里根据他们对多肽亚单位的丰富知识提出了多肽链结构。现在没有证据可证明沃森和克里克在1951年时也提出了DNA结构。1951年,他们是否看到了富尔伯格在1950年《斯堪的纳维亚化学学报》上发表的关于核苷立体化学的论文?从留下来的文件中,没有迹象表明他们曾看过这篇论文。但是,他们是了解阿斯特伯里在1947年写的著名论文的。在这篇论文里,关于多肽链连接提供了正确的信息。但是,他们不可能找到关于亚单位键角和原子间距的信息。只是在后来,才获得了这方面的信息(皮特,1948;克卢斯和科克伦,1949;布鲁姆黑德,1950)。
当然,如果说在这时候沃森和克里克还不知道科克伦和布鲁姆黑德在卡文迪什的工作以及托德关于多核苷酸链连接的工作,那是难以置信的。沃森来到以前,克里克和科克伦就在卡文迪什老大楼的顶层上,使用同间实验室。科克伦在1951年发表了关于腺嘌呤盐酸盐晶体结构的论文,他确定了接近环的那些质子的位置(见图20.1)。质子确实具有固定的位置,而且克里克早该在他的模型中反映出这种情况。显然,这是最简单的研究路线,但这一点是否得到了证实呢?克里克认为,情况并非如此,因为在DNA中的或游离状态的腺嘌呤同带一个额外氢原子的盐酸盐之间可能存在着巨大的差别。
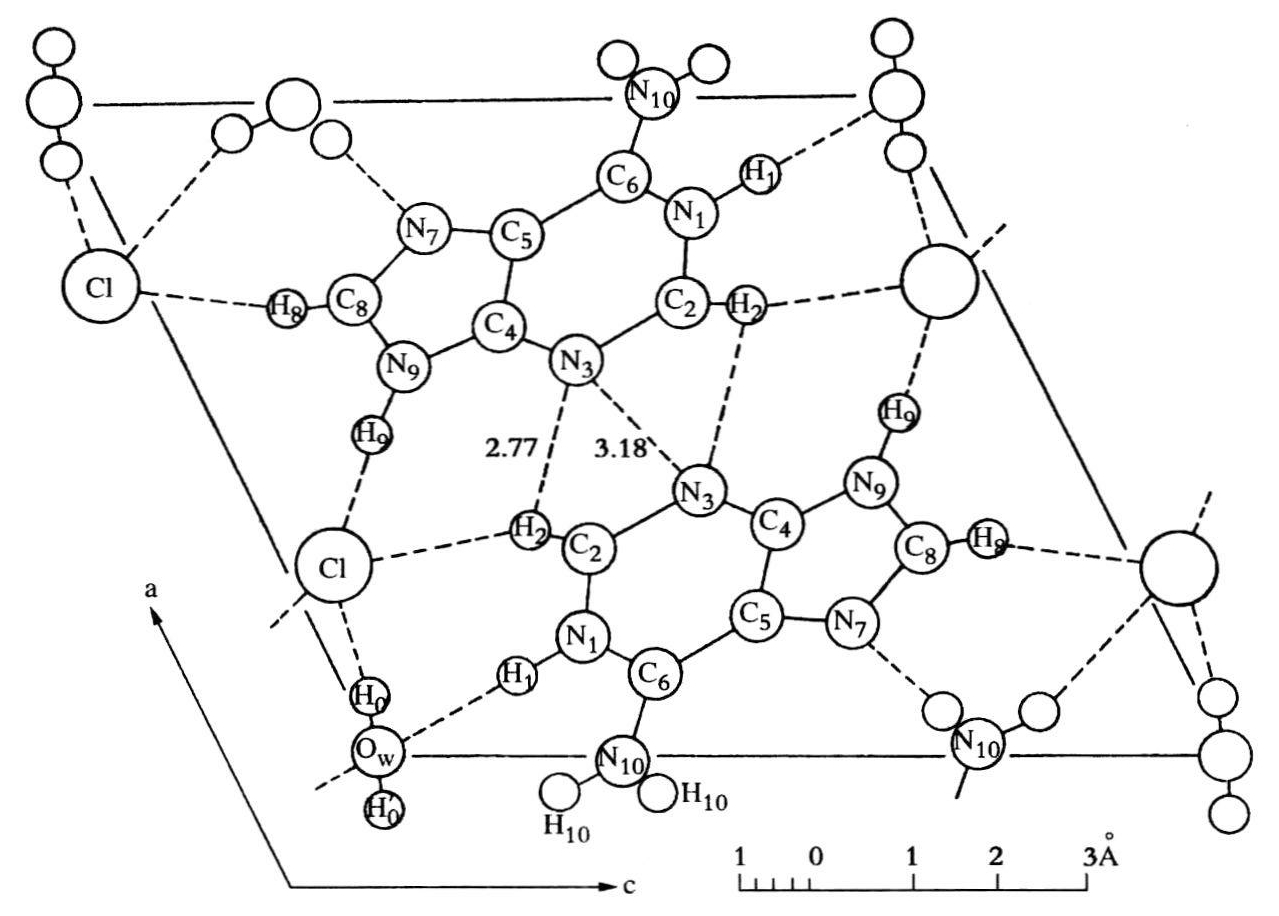
图20.1 腺嘌呤盐酸盐结构图(录自科克伦,1951,88)。
克里克宁可把一些不肯定的条条框框抛在一边,而以尽可能少的假设来建造模型。采用这种办法就可以消除许多可能性,然后利用所有可能的数据从那些最有可能的结构中作出抉择。
沃森回忆说:“我们需要做的只是构思出一组分子模型,然后摆弄这些模型——运气好的话,得到的结构就是螺旋。”(《螺旋》,50~51)但是,如果最终得到的结果不能同DNA产生的衍射图相匹配,这种做法就将是徒劳的。阿斯特伯里在1947年发表的论文中,有半张DNA伸展膜的X光图。这张图是贝尔在二战前拍摄的,图上显示出27Å的重复,3.4Å的子午线弧及沿赤道有16.2Å的侧距。但是,衍射最大值很不清楚。若要得到更可靠的数据,他们就得求教于克里克的朋友威尔金斯,当时威尔金斯已从美国回来了。因此,在周末,威尔金斯被请到了剑桥。这样他们就获悉,自从1950年的夏季以来他没有拍摄到更清晰的DNA晶体的照片。然而,威尔金斯还是设法使他们注意到他在佩鲁茨召集的第二次蛋白质讨论会上提出的有关DNA螺旋结构的证据。而且,由于威尔金斯被排除在富兰克琳活动圈子的外面,他们也不能从威尔金斯那)儿打听到她的最新研究成果。因此,他们只能参加在皇家学院举行的那次讨论会以听取富兰克琳的研究工作报告。
沃森参加皇家学院举行的讨论会
克里克可能已另有安排,因此派沃森参加该讨论会。此时,他已在剑桥呆了六个星期。在这期间,他想方设法弄懂X线结晶学原理并向克里克学习。遗憾的是,他当时还未学到家,因此当克里克询问会议的详细情况时,沃森只能含糊其辞地回答。大约过了一个星期,沃森和克里克就得出了一个模型,克里克就如何解决这个问题写了一份摘要。
胸腺核酸钠的结构。一种可能的研究方法:摘要
引言
1951年11月21日,伦敦皇家学院的研究人员在讨论会上发表了他们的研究成果,这对我们是很大的触动,促使我们去设法找到决定DNA结构的基本原理。虽然有些实验结果已为我们提供了许多启示,但在我们的研究方法中,我们试图采用最少量的实验结果,其中包括DNA结构可能呈螺旋形、晶胞的大小、每个晶格点的残基数和水的含量。我们由此获得一种试验性的结构,我们把认为是重要的特性作了如下概括,并以此作为假设。
假设
1.所有残基都是相同的。正如波林已指出的,这种情况必然形成了螺旋结构。我们没有被这一假设完全束缚住。这个假设可在两个方面放宽:(1)每个残基的某些部分是相同的,但另一些部分可能是不同的。因为不同残基有不同的碱基环,因此严格地说这是显而易见的。(2)残基可分成几组,同一组的残基是相同的。
目前我们赞同磷链和糖环都是相同的那些结构。
2.带电原子支配着结构。除了在碱基环和水中,没有原子能够“贡献”氢键,因此在该结构中氢键不像在多肽α-螺旋中那样起主要作用。与范德瓦尔力相比静电力是如此之大,以致我们相信Na+和PO2-是决定排列的主要因素。
从上述两条假设我们推测有不那么肯定的第三条假设。
3.Na+和PO2-都非常靠近在单圆柱体的表面处。
这个结构如是严格呈螺旋状,所有的Na+离子都位于一个圆柱体的表面,而所有的PO2-离子位于另一个圆柱体的表面。我们推测,在这两个圆柱体的半径很接近时,将产生最小的局部中和,通过计算,也许能验证这种想法。(在同佩鲁茨和肯德鲁交谈中,他们指出这一条件不是最普遍的,发生中和的条件可能是Na+半径近似于两个氧原子半径的平均值。)
系统方法
根据这些原理,我们现在可系统地着手解决可能存在的结构问题。首先,必须为Na+假定圆柱体表面的配位数。因为氧的半数电荷,因此配位数可能是偶数。2作为配位数太小了,因此不可能是2,而8又太大了。这样剩下的就是4或6。迄今我们倾向于4,但是6并不是不可能的,因此也应对6作些研究。在这个问题上,简单的静电学也可助一臂之力。
在选定了配位数之后,下一个问题是系统地写出所有可能的拓扑组合。先写下能满足第一个假设,一个无限大平面表面上所有可能的连接图。然后从表面上分离出一长条并折叠成一个无限大的圆柱片层。最后系统地揭示出这样做的方式有多少种。
在获得了所有可能的连接图后,接下来就是建立它们的模型。在这一阶段,我们只需考虑Na+和PO2-是分离的,并按此建立模型。然后对模型作一系列试验,试验期间愈来愈多地应用实验数据。例如:可证明任何两个磷原子同糖不可能很好地连接,因为它们分得太开了。后来又证明,只有当正常的范德瓦尔条件被破坏时,才有可能把它们连接起来。每27Å长的螺旋中的残基数可能是错误的。在这样做的过程中,特别重要的是不能把含糊的标准作为否定的根据。例如,不能把是否符合与X线图有关的先入之见作为判定依据。如果不十分小心,很可能只是因为在后阶段的某些困难而抛弃正确的模型。
实例
为了更清楚地说明问题,下面解释一下我们至今主要遵循的方法。
我们首先取配位数4,基本网如下:
上面仅是一种拓扑图,角度不是像图示的那样。根据下列条件可精确地确定任何结构:
(a)围绕圆柱体一周所通过的钠离子数。
(b)围绕圆柱体一周后,在起点的上方或下方的钠离子数。
给定横切该结构的一条明确的规则后,就能使上述两项条件更为精确。我们所用的一条规则是:“在任何结点上取对角相反方向的路线。”因此,O、P和O在拓扑学上看来都位于一个点上。
我们已大体建成了一些2Na结构。这些结构在27Å长度上给出了大约16个残基,虽然对伸长的DNA来说,这个数值可能是恰当的,但我们暂时把这些结构搁置一边。更有意思的是3Na结构,因为这种结构在27Å长度上给出了约24个残基而且在中间有一个大小可容纳8个水分子的空洞。我们现在要系统研究这些结构。
最后,在这次研究中我们注意到了一些过去尚未发现的问题:
1.如果螺旋是由3个分离的多核苷酸链交织而成,而且X线图在27Å后重复,那么这样的螺旋在27长度中的残基数不一定是3的倍数,因为这一重复可能来自一条链到另一条链间的距离。
2.一条螺旋要适合于相邻的、取向相同的同样的螺旋,很可能在一条链上有一个凸起,而在另一条链的相对位置上有一个凹孔,因此对任何给出的螺旋图来说,我们可以粗略地预测单斜角β。
3.在核糖核酸中,额外的氧所处的位置很可能使DNA结构不能成立。也就是说,它的硬脂效应可能比它形成氢键的能力更重要。
(克里克和沃森,1951)
这是很说明问题的文献。它表明克里克对α-螺旋的看法。波林第一次提出α-螺旋模型时,就不采用α角蛋白的X射线证据。卡文迪什研究小组使用α角蛋白的数据否定了非整数螺旋。阿斯特伯里依然不承认α-角蛋白存在的螺旋折叠,因为他不明白如果链是解旋的,怎么使链之间的硫氢键保持完整。因此,克里克强调了在开始时有必要采用系统的和全面的研究方法而少用经验数据。
由于沃森、克里克和波林都提出骨架位于内部的三线模型,因此人们认为有理由按运动方式建造分子。这两位剑桥大学的科学家就是按这些思路来论证的。碱基具有不同的形状和大小。碱基沿多核苷酸链的排列顺序可能很不规则,然而DNA形成了有序的聚集体从而产生了晶体衍射图。这种有序一定是由于分子的有规则的特性,因此与碱基无关,而与糖-磷骨架有关。所以,我们在构建模型时,不必考虑碱基。
作出了上述决定后,他们很自然地认为碱基是朝外的,处在这种位置上他们就不考虑如何排列骨架结构上的原子以产生似是可能的模型。他们后来评论上述推理时指出,看来“这种结构是根据所有核苷酸共有的特性。这提示我们首先应主要考虑磷糖链构型,每个糖‘平均’连接一个碱基。换句话说,理想的多核苷酸的单体都是相同的”(克里克、沃森,1954,83)。
把骨架放在分子内部还有一个原因。有趣的是,这正是克里克本人引进的对一种化学性质的限制。这关系到碱基,碱基被描述成具有几种互变异构形式。互变异构,意味着在分子中质子从一个地方迁移到另一个地方就可以改变碱基的结构形式(见图20.2)。因此,克里克否定了碱基间的氢键促使长键结合成刚性螺旋体。克里克阐明了他论证的要点。
我们知道氢键对α-螺旋是很重要的。我假定每个碱基实际上有不止一种互变异构形式,这是完全错误的。我还认为,即使每个碱基的不太正确的互变异构形式存在的时间只占5%或10%,仍难利用碱基的氢键来构成有规则的结构。因此,我很错误地断言,在形成这种结构中,氢键很少有或者根本没有作用。
(克里克,1968/1972)
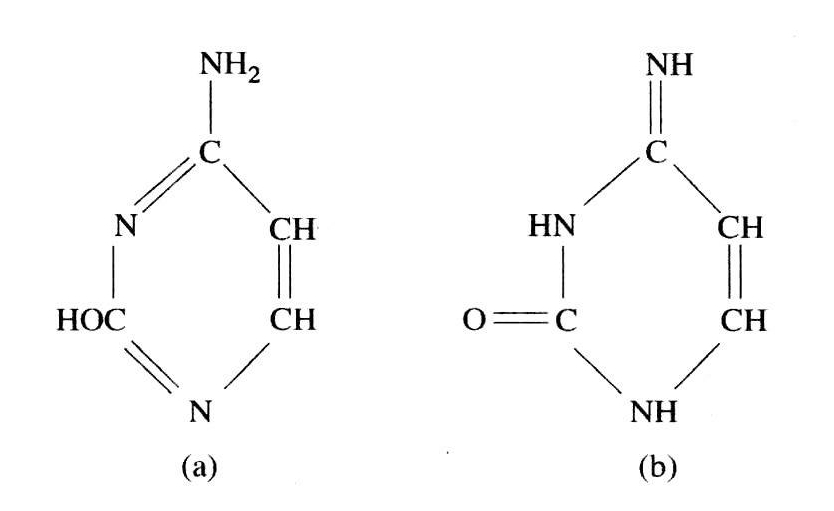
图20.2 胞嘧啶模型:(a)氢基和烯醇式;(b)亚氨基和酮式。
克里克认为,氢键与水分子也不能把这种结构结合在一起,因为根据沃森的看法,几乎不存在水。不存在水意味着环绕在局部电荷周围的水壳不能中和这些电荷。因此,带电的Na+和PO2-必须靠得很近,随之而来的结论是,DNA钠盐带不同电荷的成分之间的引力,使这种结构结合在一起。
我们曾试图建立一个平衡正负电荷的模型。这完全是错误的,因为事实上存在着许多水。由于我对化学不很精通,不知道像钠这类物质很可能是水合物。否则,我早就发觉这一错误了。这不仅是沃森犯了这个错误,我也没有注意到这个错误,同样该受到责备。因此,这完全是浪费时间。沃森作了相当精确的说明,但这个结构比他描述的更系统些。我们画了几个网,我又把问题形式化并提出了几条几何定理,但对我们都毫无用处!
(克里克,1968/1972)
当时沃森和克里克对于由磷酸骨架的带电基团把三股螺旋结合在一起感到非常高兴(见图20.3a)。骨架可能显得有些不顺眼,但碱基是非常明显的,立刻可同蛋白质反应。后来克里克在解释他们这种天真的想法时说:“要知道,我们从来也没有做过模型,如果我有这方面的经验,当时就会知道我们压根儿没有取得什么进展,这是我们后来才逐渐明白过来的。要是我们的同事们,例如约翰·肯德鲁不说,如果不告诉他们,我们就不能再继续干下去,我们也许不会这么快就给皇家学院的研究人员看我们的模型。那时我们应该再等一两个星期,定下心来再作些探讨。这是我们失败的部分原因。现在回想起来我们当时提出的模型一点用处也没有,我们从未很恰当地建造模型。正如沃森所说,做这样的模型是非常困难的。”(同上)
他们听从了同事们的劝告,给威尔金斯挂了电话。第二天,威尔金斯、西兹、富兰克琳和戈斯林专程到剑桥来看这个模型。沃森描述了突然陷入的尴尬场面,那是当克里克兴致勃勃地讲述了科克伦、范德和他在四星期前得出的螺旋理论的重要性后出现的。沃森和克里克还没有来得及说明他们的模型是同X线数据相符时,威尔金斯已在抱怨他的同事斯托克斯早在去年夏天就研究出了这个理论,富兰克琳在克里克讲述螺旋的整个谈话过程中一直流露出轻蔑的表情。作为X线结晶学家,她知道要弄清这些X线数据即使是讨论可能存在的结构,他们还差得远呢!分子可能是螺旋形的,但目前“毫无证据”可证明这一点(《螺旋》,94)。至于正离子的作用,它们为什么一定被水壳包围着,这意味着在一段距离上它们的电荷能被中和,因此正离子不大可能同磷紧密结合在一起。沃森把水的组成搞错了,不像他报告的每个晶格点上有4个分子而是每个核苷酸有8个分子(富兰克琳,1951b)。各种各样的结构都是可能的,因此支持他们这个模型的论据就站不住脚了。
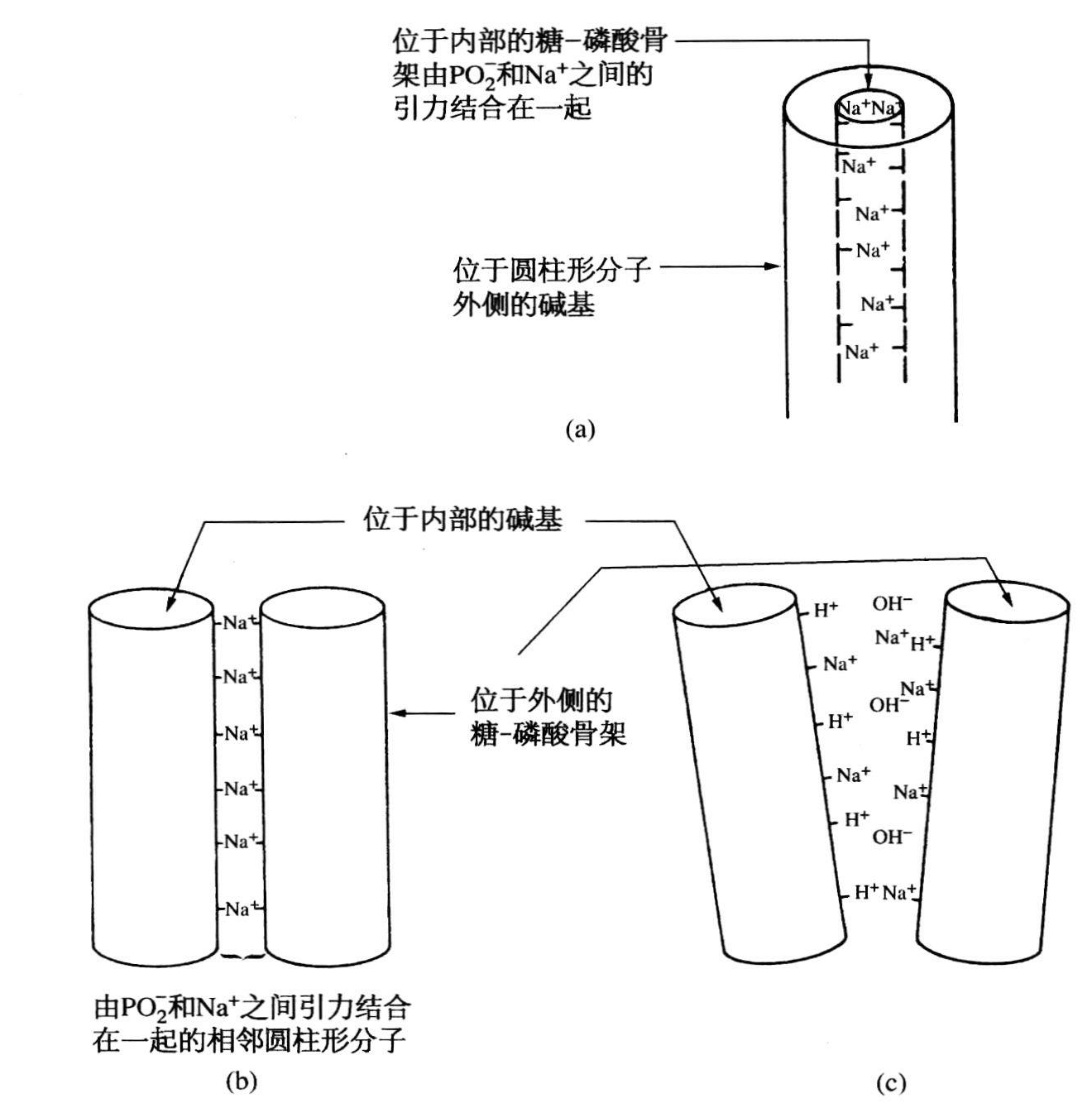
图20.3 (a)沃森和克里克在1951年提出的方案;(b、c)富兰克琳在1951年提出的方案。须注明的是,在1951年时没有画什么图,这些图是本书作者根据他们的手稿绘制的。
沃森和克里克为了打破这一僵局,建议两个研究小组今后携起手来协同工作。不难理解富兰克琳和戈斯林不愿采纳这一建议,因为他们已目睹这两个小丑所干的蠢事,为什么要浪费精力去同他们合作呢?所以,皇家学院实验室的四位科学家立即返回伦敦,让泄了气的沃森和克里克孤零零地呆在剑桥。
下一次——如果有这么一次的话——沃森和克里克就不会那么粗心大意了。今后他们再也不会听从同事们提出的必须把研究结果告诉威尔金斯的劝告了。对他们来说,这个教训是够深刻的了。然而,富兰克琳关于DNA结构与水的关系的报告,沃森显然不是很听得进的。她已指出,这么容易吸收水的结构尽管有大量的非极性基团(碱基),只有当极性基团(磷)位于分子外面时才能这样做。由于暴露的磷同介于相邻分子间的正离子非常接近,因此这种“晶体”形式是很有规则的。所以,这些长棒形分子是“背靠背”地紧密相连。干燥可强化这种有序效应;湿润则逐渐破坏这种结构(见图20.3b、c)。虽然沃森和克里克不知道这一点,但大量的物理化学证据可证实富兰克琳的推断。
电势滴定
约翰·马森·古兰德从供应部回到诺丁汉大学不久,就开始了小牛胸腺核酸的电势滴定。当时认为核酸溶液在加了酸或碱后黏度发生改变是由于共价键断裂而引起的解聚作用。后来,古兰德及其同事们根据蛋白质的情况提出另一种解释。他们论证说,引起黏度的变化类似于蛋白质的变性,即氢键的断裂。当pH值大于或等于2时,所有主要的磷基团都被离子化了。但是,对pH值大于2的核酸溶液滴定时,却发现其他的解离作用。这是发生在pH值为4~5或11~12时,中间隔了一稳定区(pH为5~11),此处没有任何基团释放出来(见图20.4)。同时存在着一种滞后现象,因此在回滴定时,这段稳定的pH范围就缩小了,这说明在有胶束碎片的环境中,核苷酸的PK值是不同的。他们论证说,pH值为酸性时,鸟嘌呤、腺嘌呤和胞嘧啶的氨基被释放出来,鸟嘌呤和胸腺嘧啶的“烯醇羟基(我们曾称它为酮基)”则在pH值为碱性时释放。在中性时,这些基团显然连接在一起,其中最可能的键就是氢键。能滴定氨基和烯醇基的pH值正是DNA黏度大大降低之时,这说明黏度的降低是由于胶束的分裂引起的,胶束本来是由氢键连接的几条链组成的。“……人们认为由于氨基和烯醇基之间键合的断裂,使胶束解聚为较小的分子单位……”(古兰德和乔丹,1947,60)

图20.4 牛胸腺脱氧戊糖核酸滴定曲线。Ⅰ是正滴定曲线,Ⅱ是回滴定曲线。数据是古兰德、乔丹和泰勒在1947年测得的(图取自乔丹,1952,70)。
1951年,D.O.乔丹在兰德尔新办的杂志《生物物理学进展》上发表了这一研究成果。皇家学院实验室的研究小组肯定也看到了这篇文章。这样,乔丹终于能提到晶体状态下碱基间形成氢键的证据。古兰德和乔丹把这些键画在同一条链上或相邻链上的核苷酸之间。乔丹显然不赞同DNA是螺旋分子的观点。他知道,在没有电解质的溶液中,伸展的DNA分子构成了极不对称的棒状体。他认为这是带负电荷磷基之间的斥力造成的。加了电解质后,这些电荷被中和,分子就卷起,像图20.5所示。因此没有迹象表明,相邻链上由氢键连接的碱基受到了它们周围螺旋状糖磷圈的保护。这正是富兰克琳于1951年11月提出的。

图20.5 加了各种试剂后脱氧戊糖核酸分子的形状可能发生的变化(录自乔丹,1952)。
对沃森、克里克和威尔金斯的责难
皇家学院实验室人员参观了沃森-克里克所做的三股螺旋模型后,在兰德尔和布拉格之间引起了一场讨论。他们一致认为DNA结构的研究留给兰德尔的小组去做。克里克集中精力完成论文《多肽链和蛋白质:X射线研究》。沃森是正式指派给植物病毒学家罗伊·马卡姆的,因此他应从事TMV的研究也是顺理成章的。沃森认为,研究TMV可能提供有关DNA结构的线索。在6月,他的确也成功地拍摄到具有螺旋特点的X光照片,但图像太复杂未能提供他需要的那种线索。
与此同时,威尔金斯的研究工作遭到了挫折。他从美国一回来,就同意富兰克琳使用仅有的赛纳的DNA。威尔金斯改进了1950年时用的单晶照相机,“使其能更好地瞄准,并于1951年12月从查格夫提取的大肠杆菌、麦芽和猪DNA的B图型中,观察到层线的赤道反射”(威尔金斯,1968)。伴随着这成功的却是两项失败。他得不到查格夫的DNA样品作A/B转换,而且他根本得不到A图型。这些点的分辨率也不及富兰克琳在1952年用赛纳的DNA所得到的那样高。因此,在DNA上难以取得进展。
幸而,威尔金斯还可以从事核蛋白结构的研究。因此,在这架性能更好的照相机的帮助下,他拍摄了乌贼精子的衍射图。1952年春,在动身到欧洲大陆度假和工作旅行前,他向克里克通报了所取得的成功。
我已拍摄到好得多的乌贼精子的X光照片,这些照片清晰地显示了一系列螺旋层线和胶束间的空隙。如果用没有取向的样品,这些点不会重叠。我打算进一步研究在试管内有活力的精子。我记得你在罗马的那位朋友曾答应给我一只X光管……
我研究了钠胶束变换,这些胶束凸出在与轴线成直角的一个平面上,更多地研究了层线强度,同时我也重复了奥斯特和赖利在湿胶上定向的长距离的研究。我已发现你提出的许多设想是很有价值的,但我很相信有许多理由说明磷一定是位于外表面。现在照片上只有一个部分仍使我迷惑不解。但是,如果螺旋是正确的,那么我们一定能很快作出解释。我本人确已专注于这项工作,但在模型制作和化学上还乏善可陈,因为我认为我们还没有认识到这种X光照片的关键作用。当我们完全搞清楚这些照片后,模型问题就可直接解决了。现在,我自己拍摄X光照片并做了几架新的照相机(因而获得了更佳的精子照片)。但是,同你一样,我也认为解决核蛋白的关键在于晶体照片。确切的结晶性的重要性在于能明确地告诉我们一般的胶束排列情况。虽然富兰克琳常常非难我的观点,但她未能真正打倒我。由于重新安排了时间,我得以集中精力研究这个问题。这样她再也难不倒我了。上次我碰到你时,当时我的日子可真不好过啊!
圣诞节前我见到了吉姆,他说根据你们的办法是做不出模型的。如果真是这样,事情就简单了。总之,我相当肯定磷是位于外表面。我不愿重提我们之间在核酸问题上的“争执”,期待着今后能同你们再次讨论我们的最新看法和研究成果。下次来时让我们共进午餐吧!
(威尔金斯,1952)
接着,他又概述了从活细胞核或整个细胞得到衍射图的计划。他这样做,是为了同生物化学家从分离出来的无定向核蛋白获得的衍射图作一比较。
这样我们也许能证明,螺旋观点既适用于精子那样含水量极少的细胞,也适用于一般的细胞。但是,即使我不出去一两天,这项工作还是需要一段时间的。你是否听说我已收到里奥的邀请。这妙极了!从某些方面来说,我觉得我是个幸运儿。我希望收集到含有分开的染色体的精子束,分别拍摄到细胞中螺旋状和非螺旋状两种染色体的X光照片。这样做也许是多余的,但我觉得在一两年后也许是很有用的,可以把这些X光照片同真实的染色体更多地联系起来。但是,愿我这次旅行不会一无所获,从工作观点来说,我认为这次旅行还是很值得的。
(同上)
从这封信中我们可看出,自从11月份威尔金斯到剑桥后,他同沃森和克里克的唯一接触就是圣诞节前沃森一次短暂来访。威尔金斯当时还不想做模型,因为他确信,“我们还没有认识到这种X光照片具有的关键作用。当我们完全搞清楚这些照片后,模型问题几乎就可直接解决了”。信中还清楚地表达了威尔金斯乐意同克里克分享他的研究成果。
那一年的晚些时候,威尔金斯从法兰克福的A.费利克斯那儿搞到了已去除尾巴的鲑鱼精子头部。这些精子是未经固定和未经干燥的,因此威尔金斯预料“这些精子的结构同活着时的结构是相似的”。因为,从光学上来看,这些精子是各向同性的,因此它们更接近于普通的细胞。
这张照片同乌贼精子的照片是相似的,不同之处是每个弧已成为一个完整的环。这说明,无论是各向同性还是各向异性的精子头部都是由类似的长链核蛋白分子的晶体所组成。
(威尔金斯和兰德尔,1953,192)
虽然人们承认提取的DNA和原位DNA显示出相同结构特性所具有的重要性,但仍想知道威尔金斯所说的精子图型近似于哪一种结构:“湿的”结构还是“晶体”结构。1952年,威尔金斯给克里克的信以及他同兰德尔合写的报告(《生物化学及生物物理学报》于1952年11月27日收到)都没有提到这个问题。最近,克卢格提到下述情况:“……没有明确确定周期性,也没有报道对这两种已知的尚未发表的形式的解释。”(克卢格,1963,844)但是,我们必须记住威尔金斯在1951—1952年期间不能得到唯一能产生A型的赛纳的DNA,因此他未能产生A型。从威尔金斯给克里克信中所画的那张图上可看出,在层数上由强度组成的交叉线,威尔金斯分别标上1,2,3。尽管我们都承认还有一些缺点,但老实说,谁能说这种乌贼图型看上去既不像A图型也不像B图型呢?好多年后,威尔金斯对此总结如下:
到1951年12月,所有图形上的赤道反射都粗略地给出了DNA螺旋的外径和确定了A和B结构的螺旋间距。1952年2月,也从精子头部观察到螺旋间隔。我想大约在1951年和1952年我常常到克里克他们那儿去,因此我能把DNA分子的一般图像、链的近似数目、螺旋的螺距和直径以及碱基堆砌等告诉克里克和沃森……
(威尔金斯,1968)
1952年5月,沃森写给德布吕克的长信中曾部分地提到了这些情况。关于DNA,他是这样写的:
DNA的结构:最近伦敦皇家学院的莫里斯·威尔金斯已拍摄到取向的Na+胸腺核酸的极好的X光衍射照片。这些点极为清晰,表明在三维上极好的取向并充分提供了细节,可对该图形作出严密的解释。在下列形状(图20.6)的单位晶胞的基础上,已标定了该图形。根据密度的论据,这个晶胞可能含有14~17个核苷酸。皇家学院的研究人员确信,这些核苷酸排列成螺旋状,但迄今为止,获得的X光证据远远不及合成多肽的证据那样令人信服。
图20.6 沃森给德布吕克信中的单位晶胞。
很明显,需做大量的工作来阐明这种结构。但是,皇家学院的那些人却卷入了内部的争斗,因此目前他们还没有真正作出努力来解决结构问题。我们已试图使他们对波林的模型制作方法发生兴趣。事实上,今年冬天我们确实花了几个星期,试图建造似是可能的模型。然而,我们最近出于政治原因,暂时停止了这项工作,我们一个要好朋友没有参与这项研究工作。但是,如果皇家学院的研究人员仍一事无成,我们要再次碰碰运气了。
虽然皇家学院的研究小组在主要问题上没有取得进展,但他们已搞清了两个问题:
1.牛胸腺、鲱鱼精子和大肠杆菌纯化DNA的Na+盐的X光照片完全相同,即沿轴线每隔27Å有一重复——因此看来有一个基本的DNA结构图(至少Na+盐是如此)。
2.定向的完整的乌贼精子的X光照片显示出与提纯DNA相同的DNA图型!!!这就表明精子里的DNA是以一定方向排列的。定向乌贼精子的“粉末”照相给出了有环的粉末图,环对应于定向DNA照片上很强的点子。这样就可能表明,在不能定向的样品里有定向的DNA。从这个观点出发,我同莫里斯现在已开始研究T2噬菌体。
(沃森,1952)
关于这方面的情况,威尔金斯写道:
在那时,光学测量结果是唯一能证明化学家从细胞中提取的DNA的结构,同完整的染色体结构是一样的。X线研究工作提供了直接证据,证明两者在许多细节上都相同。研究晶体DNA的X线结晶学家没有研究化学家的人工制品,而是在研究同基因性质直接相关的结构。
(威尔金斯,1972)
但是,沃森信中的结晶学数据是什么呢?根据沃森在图20.6所标出的尺寸,看来他涉及的不是单位晶胞而是原始晶胞。即使如此,他提到14~17个核苷酸仍是奇特的。确实,在早些时威尔金斯已提出在晶胞中可能存在螺旋结构(见第十九章)。看来沃森是根据威尔金斯以前的论点行事的。另一点值得注意的是,没有提及DNA存在两种形式,也没有提到在湿度高时得到的照片上有34Å重复的情况。在这些问题上沃森是否也听从威尔金斯?看来,这也是很可能的,因为富兰克琳无论在1951年11月讨论会上还是在她第一篇的研究员报告中都没有提到这些数据。因为这封信可表明沃森、克里克和威尔金斯对富兰克琳和戈斯林最新获得的数据是一无所知。因而沃森认为,威尔金斯新拍摄到的鲱鱼精子和大肠杆菌的DNA是属于晶体A型。显然A、B型的强度数据的这种混淆不清,不仅被波林而且被沃森和克里克在1952年继承下来了。这一结论可突出说明当富兰克琳和戈斯林根据极为清晰的X光照片确定了不同子午线弧的位置时,他们作出的重大贡献。
为什么这样说呢?威尔金斯说道:“当然,早就可以测定这种位置,因为在富兰克琳的第一张B图上,这些层线条纹是明显的。我认为正是由于没有对B图作精确测定,所以A、B周期都在30Å左右。”(威尔金斯,1972)这可能是因为没有要求他们区分这两种形式,因为可用其他标准来区分:(1)A、B型赤道上的亮点,分别位于16.5Å和~22Å;(2)显著的子午线弧在B型的3.4Å处,而A型是没有的;(3)B型的结晶度较小。大约在1951年12月和1952年1月,威尔金斯得到一幅更好的乌贼精子图型,头四条层线上的弧线清晰地标记出来。1952年春,他写信给克里克,谈到了这幅图。虽然,威尔金斯没有告诉克里克,但这张图型显然是B型,而且可由此计算出34Å的重复。但是,威尔金斯“在当时没有作精确的测量”(威尔金斯,1972)。因此,1953年1月沃森第一次看到富兰克琳1952年得到的B图和知道重复距离为34Å时,他感到大吃一惊。
富兰克琳的新数据
1952年春,富兰克琳和戈斯林忙于用菲利浦微型照相机在相对湿度为75%情况下,对DNA作试验性快速曝光摄片。他们也试验了特殊的倾斜式镜台,这是专为沿纤维轴寻找广角重复而设计的。
4月18日,富兰克琳在记录本里记录了晶体型单纤维照片的详细资料,从照片上显示出某种双向现象。她在双向两字下面划了一条粗线。关于曝光时间,她未作任何记载。因为在复活节期间,她去度假而中断了这方面的试验。但看来她还是相当自信的,她认为这张照片有重要意义。皇家学会在5月1日召开的为期一天的蛋白质讨论会上,克里克和沃森获悉了富兰克琳“坚持她获得的数据告诉她,DNA不是螺旋”(《螺旋》,122)。令人啼笑皆非的是,我们知道就在那天和第二天晚上,富兰克琳在拍摄单纤维的X光照片,这种单纤维一旦给出了结晶图型后变化是不可逆的,即使在相对湿度75%时也产生“湿”的或B型图。五月初,在相对湿度为75%时进一步曝光得到的不是一张“优良的‘湿’照片”,而是一张“极好的‘湿’照片”。看来正是这张惹人注目的B型照片,在大约九个月后使沃森目瞪口呆。当时另一张很好的B型照片使她能把大约在22Å处的很强的赤道反射分解成由相距2.5Å两个点组成的一个双体。这个双体相当于一个分子厚的一层水。她写道:“这表明有两个相共存着,这两个相只由一个水分子层把链单元分开。”(富兰克琳,1952a)威尔金斯认为观察到这一情况“实属偶然”。在后来广泛而深入地研究DNA时,再也没有观察到这一现象,因此他把这一现象归因于两种不同的膨胀状态(威尔金斯,1972)。
富兰克琳也算出了第一层线的间距介于33~34Å。由此她得出结论,在子午线3.4Å处,很显著的弧线一定是在第十层线上。在那个月的笔记本上,她这样写道:
清晰的赤道点及在层线上的3.4弧,表明了即使在“湿”状态下,螺旋(如果存在的话)每一圈都存在着整数的(或一位小数)残基。
从“结晶”向“湿”变化过程中,主要的赤道间距约增一倍,纤维轴线周期延长了25%(27~34Å)。
富兰克琳反对螺旋的数据
如果DNA纤维是由与纤维轴平行的圆柱形螺旋组合成的,那么在该图左右两方的点子的指标应是径向对称的。但是,当富兰克琳和戈斯林开始标定A图时,发现不是这么回事,因此促使他们采取了反对DNA是螺旋的态度。4月,发现了一起双向事例,他们以此来检验所有指数的可靠性。双向本身与螺旋结构并非是不可共存的,但标定指数后获得的结果似同所认为的强度经向对称相抵触。对左(L)和右(R)象限内相同强度的点进行了指数标定。“这时发现,所有标为L的反射的指数均为hkī,所有R反射则均为hkl。”(富兰克琳。1953,7)8月,她试图重新获得双向,但失败了。因此,她的结论是这可能是偶然的巧合,或是接近表面晶体的优先取向。威尔金斯回忆说:“我们实验室的关于DNA的一次会议上,富兰克琳描述的DNA不是螺旋的事例其依据不是双向现象,而是认为hkī反射比hkl反射明显地要强得多。就她提供的那些数据来说,是与螺旋结构不相容的。”(威尔金斯,1972)
正如克里克向我解释的那样,平行排列的螺旋分子纤维图能够呈现出径向非对称的现象,这是因为分子是聚合在单斜晶胞内,因此从不同方向看到的晶格也是不同的。取样点改变时,这种变换可发生得很快,结果使图中不同部位的重叠强度图也不同。当时,无论是富兰克琳还是戈斯林都没有把这种可能性放在心上,因此有讽刺意义的是,他们最初反对螺旋的态度是依赖于这些指数标定。
为了解决指数标定上模棱两可的问题,富兰克琳和戈斯林继续计算A型圆柱的平均帕特森函数,结果导出了可靠的晶胞参数。这项细致的计算工作是在1952年夏季完成的。1952年9月初,他们为接待医学研究委员会一个小组委员会的来访所写的一份报告里,提供了很重要的信息:
根据所获得的帕特森函数,可以方便地确定晶格变换。把晶胞的a、b、c分别写为22.0Å、39.8Å和28.1Å,β为96.5°,对观察到的66个不同的反射都可以标定指数,且误差小于1%。
(兰德尔,1952)
异乎寻常的是,富兰克琳和戈斯林在这篇报告里一点也未提及她对螺旋所持的反对态度。威尔金斯报道了这些观点,并以一种不太愿意接受这些观点的口气作了如下描述。
应用两维数据,最合理的是用螺旋结构来解释,而且关于螺旋的实验证据,比从任何蛋白质得到的证据都要清楚得多。晶体材料(A型)的X光照片具有许多可以用螺旋观点解释的简明要素,但三维数据清楚地表明:该照片的简明性的基本的物理学解释,在于一些颇为不同和不太可能的结构特点。20Å的单元,虽然其截面基本呈圆形,但看来具有很不对称的内部结构。
(兰德尔,1952)
这里应为威尔金斯剖白一点,他是过了好久才看到富兰克琳反对螺旋的数据。威尔金斯只是讲了富兰克琳对从A型晶体得到的证据的口头意见。就富兰克琳来说,她从来没有以书面形式发表过直接反对螺旋的观点。1953年,她在《结晶学报》上发表的关于圆柱体帕德森函数时(见图20.7),只是写道:
可观察到c-面心的单斜晶胞在投影上近似六角形。但是,有证据表明:晶体本身绝不是呈圆柱对称。虽然造成纤维晶体双向的巧合原因还是一个谜,但看来,如果各个晶体同纤维轴具有高度对称的话,根本不可能发生这种双向现象。
(富兰克琳和戈斯林,1953e,684)
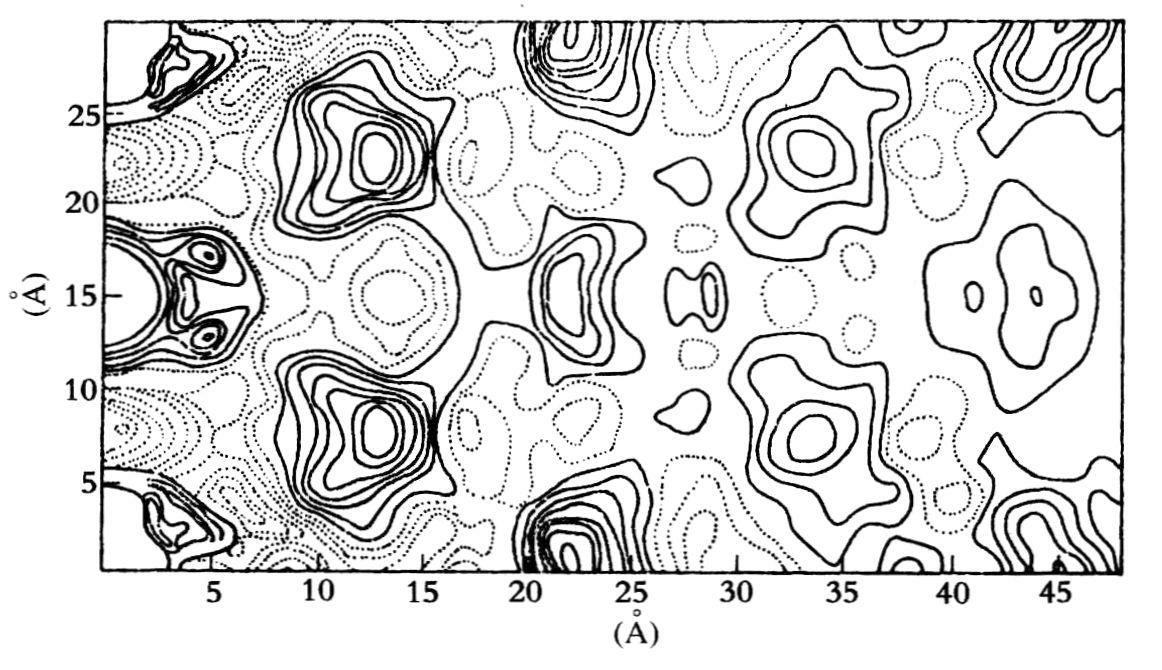
图20.7 在作者得知沃森-克里克的模型前绘制的NaDNA圆柱帕德森函数(录自富兰克琳和戈斯林,1953e,682)。
在这篇论文的打字底稿上,我们却看到了有一个词做了改动。“有证据表明结构单元……”这里的结构单元,她是指在晶胞中形成结构单元的分子或分子群。如果看一下原来的手稿,我们可看到这样一句话:“无论是结构单元本身或是这些单元组合的方式都是极不对称的。”但是,1952年7月的笔记本里,她又怎样写的呢?首先她画了一张一般形式的帕德森函数图(见图20.8),然后写道:
没有迹象表明存在直径为11Å的螺旋。中间香蕉形的尖端同每个晶胞有两圈直径为13.5Å螺旋的计算曲线相符。如果是螺旋,也只能是单股螺旋(双股螺旋将形成8字型)。
螺旋不能解释短的垂直矢量~4Å(这是叠加在尖端原点上,因此表明真正距离不是3.4Å而是>4Å)。但是,如果在该结构中存在平面状的香蕉形单元,而且香蕉轴同纤维轴平行,那么这一矢量就可作如下解释……
而且,这一晶胞的大小说明不大可能是螺旋结构——如果螺旋直径为13.5Å,那么如何填满这一长周期中的剩余空间呢?
很可能是一种双片层结构
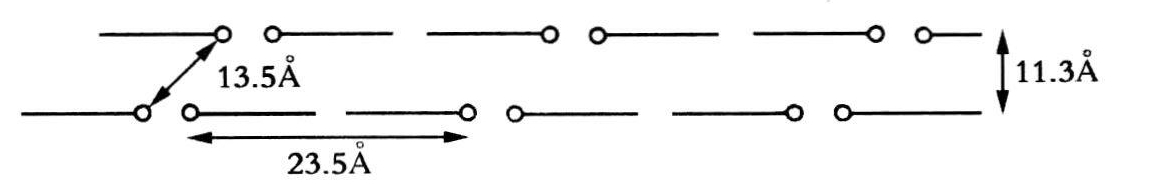
即使是这样的结构也必须用图45来解释双向现象。双向现象的原因,可能不是束中所有的纤维(虽然几乎所有的纤维都是在束中),而是同纤维表面成直角的这些片层的优先取向。

图20.8 1952年7月富兰克琳在圆柱帕德森图上的注解(录自她的研究记录本)。
克里克和克卢格认为,这种不寻常的解释表明,富兰克琳正在考虑并排存在三股直径为11Å的螺旋,三股一组的螺旋的中心距为20Å左右。每股螺旋的帕德森都应在11Å处显示磷-磷矢量,但是富兰克琳却发现在 Å处。因此,她否定了螺旋的存在。出于某种原因——可能是反对螺旋的偏见或没有想到这一点——她没有去探索其他螺旋结构的可能性。例如,她提到了两根同轴链(双螺旋)的可能性,但现在看来难以置信的是,她竟没有认识到这样的结构可根据重复距离28的一半来说明c=14Å处的峰。中间的香蕉状的峰起因于两股螺旋的交叉。5Å处的峰是一股螺旋上相邻磷之间的第一个磷矢量,而在x=6.7Å处低电子密度的谷只是当一根螺旋从原点移出而形成的下降。当然这并不表示强度为零,从图20.7中虚线表示的恒值线就可看出。如果任意选用实线表示的密度,加强了不连续性的印象。
Å处。因此,她否定了螺旋的存在。出于某种原因——可能是反对螺旋的偏见或没有想到这一点——她没有去探索其他螺旋结构的可能性。例如,她提到了两根同轴链(双螺旋)的可能性,但现在看来难以置信的是,她竟没有认识到这样的结构可根据重复距离28的一半来说明c=14Å处的峰。中间的香蕉状的峰起因于两股螺旋的交叉。5Å处的峰是一股螺旋上相邻磷之间的第一个磷矢量,而在x=6.7Å处低电子密度的谷只是当一根螺旋从原点移出而形成的下降。当然这并不表示强度为零,从图20.7中虚线表示的恒值线就可看出。如果任意选用实线表示的密度,加强了不连续性的印象。
富兰克琳对螺旋的态度
1952年2月富兰克琳写道:“这些结果表明了一种螺旋结构(一定是非常紧密地堆砌的),其中的每个螺旋单元可能有2根、3根或4根同轴核酸链。而且,靠近外侧有磷基团。”(富兰克琳,1952,4)但是令人费解的是,5月份她一反常态改变了原来看法,开始采取反对螺旋的态度。关于这个问题已作过讨论(克卢格,1968)。因此,这里我们只需列举一下她改变态度的可能原因。
富兰克琳是位专业的结构晶体学家,她不相信直观的揣测,而是想用直接的方法来解决结构问题,也就是说,不是先假设存在某种结构。她并不反对螺旋结构本身,而是在她认为证据不足的情况下,反对假设存在这种结构。这种态度是出于她作为一个研究人员的本意和她对贝尔纳的崇拜,因为她参加了1951年举行的国际结晶学会议,会上听取了贝尔纳作的题为《蛋白质结构现状》的讲演。从她的笔记中可看出,贝尔纳的话对她很有作用。贝尔纳讲到了波林的研究方法,即用化学知识做结构模型,然后用X线数据去验证。这是推测和演绎。在开始阶段这种做法是无可非议的,但富兰克琳写道:“现在要综述这些证据和假设了——我们是否找到了解决办法或一种办法。”贝尔纳把波林的演绎法,同根据帕德森断面推论链类型的经验归纳法进行了对比。“真正的验证要求在晶体图上散射分布很广,以及帕德森断面(<2Å)的最后结构。”一年之后,她无疑是遵循了这一条路线,先研究了圆柱形的帕德森分析,然后是三维的帕德森分析。DNA这样的结构看来采用这种方法是合适的,因为它的磷原子的散射能力比其他原子强得多。当然用螺旋构象来解释帕德森图是太简单了(见图20.9),但即使在那时已证明是误解了,因为如同威尔金斯回忆道:“帕德森研究得出的想法是PO4链相距c/2,而实际上相距~c/3。把帕特森峰作为标明了PO4的位置,那是过于简单化了。”(威尔金斯,1972)克卢格的论文《罗莎琳德·富兰克琳及DNA结构的发现》发表后,就这个主题讨论的结果使他作了如下说明:
……虽然富兰克琳和戈斯林计算的圆柱帕特森函数雄辩地证明了存在A型双螺旋,但是图的分辨率只有5Å左右,所以该模型的细节也只是近似的。皇家学院研究小组(富勒、威尔金斯、威尔逊和汉密尔顿,1965)后来的研究工作表明:这两条链绝非是等距的,至于三维帕特森图的分辨率能否明确无误地推论出晶胞中螺旋分子的取向,还是令人怀疑的。富勒他们认为,所用的X线数据不足以解决这个问题,即使他们的分辨率为3Å的更充分的数据,“同样也不足以解决这个问题”。
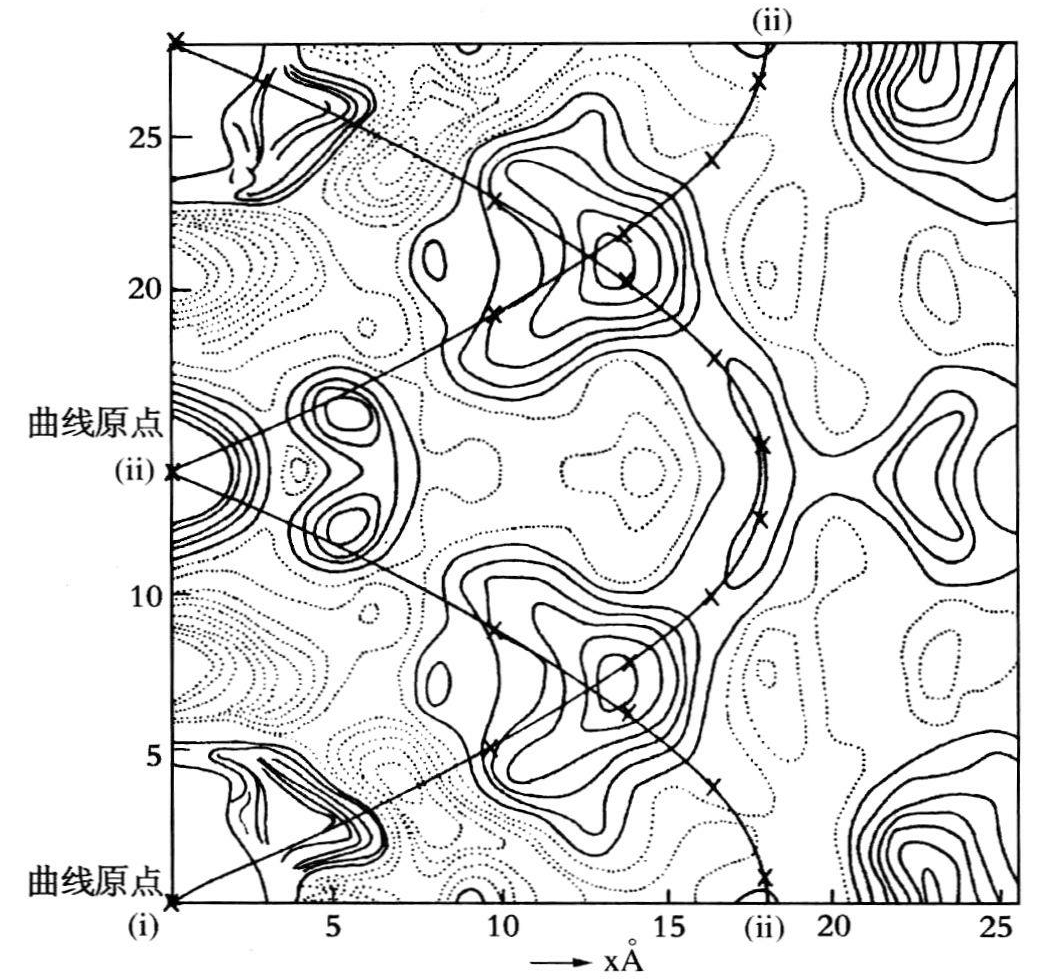
图20.9 富兰克琳和戈斯林在知道沃森-克里克模型后所绘制的圆柱帕特森函数图,其中包含了两根同轴螺旋(富兰克琳和戈斯林,1953d,156)。
(克卢格,1968,880)
富兰克琳和戈斯林当然非常关心他们的圆柱帕特森图的解释。他们为此拜访了伯克贝克学院的C.H.卡莱尔,问他:“你认为这种帕特森图会同磷酸基位于螺旋外部相一致吗?”(卡莱尔,1968)虽然卡莱尔作了肯定回答,他们还是拒绝接受这种解释。
富兰克琳因为想要利用帕特森分析,因此在1953年2月以前,她集中研究A型而没有理会在1952年5月初得到的那张极好的B型照片。从她的记录本里我们可看出,在2月10日星期二那天,她开始研究B型,几天以前沃森刚带着波林关于三股螺旋模型的手稿来到皇家学院。富兰克琳决定重新研究B型是因为沃森的访问及波林关于DNA是螺旋结构的观点呢,还是她当时已对A型作了详尽无遗的研究了呢?克里克和克卢格倾向于后一说法。1952—1953年的那个冬天,富兰克琳一直在试图解释三维帕特森图,在研究了由成对的、背靠背的反向平行棒状体组成的片层结构后,她继续研究能模拟螺旋衍射图的呈对角线堆砌的棒状结构系统。1953年1月,为了探索糖-磷骨架的可能取向和大小,她开始制作模型。当时她仍然否定螺旋结构,研究了一种8字型结构。在这种结构中,单链形成了作8字型循环的阵列。她这样做是为了说明晶胞在纤维方向上的二等分和加水能产生B结构的某种紧密堆砌的结构。
当然,富兰克琳被帕特森图引入歧途还有一个特殊原因。在得到帕特森图之前,她已拍摄到显示双向的X光照片。她第一次试图解释帕特森图后,她又拍摄到许多X光照片以期重复双向现象(1952年8月)。阿伦·克卢格暗示说,富兰克琳一贯认为B型是一条螺旋,但在1952年,她认为A型是非螺旋结构(克卢格,1968)。威尔金斯这样说:
我仍认为,她原来的强度数据表明相当大的不对称性,同螺旋结构不相一致。但是,严格检查她的测量结果后发现,强度和取向指数不够精确,因此不能证实真正不对称性及与螺旋结构不相一致的结论。
你(克卢格)认为罗莎琳德一贯把B型看作是螺旋结构,你的看法可能是正确的。但是,如果她真是抱这种观点,她也没有大肆张扬。我认为,如果A是非螺旋的,那么把B看作是螺旋结构是很不合理的。
(威尔金斯,1969)
我甚至认为,从1952年5月到1953年1月沃森来访之前这段时间里,富兰克琳不仅认为A型而且认为B型都不是螺旋形DNA。这就可以说明1952年他们在皇家学院开玩笑地举行了一次螺旋的葬礼,也可说明他们在戈斯林关于DNA一定是螺旋的结论(手稿无具体日期,可能是1953年2月)旁边画上一个骷髅,以及可说明富兰克琳给她的同事和克里克留下她是反螺旋的印象。1952年夏,克里克曾向她保证这张双向照片会把人引入歧途,但她充耳不闻。当听说波林提出螺旋模型时,她是否回到1952年2月时的立场呢?有可能是如此。
波林的三股螺旋
当问起波林怎么会对DNA发生兴趣时,他说,那是因为他认识艾弗里、麦克劳德和麦卡蒂并知道他们研究肺炎球菌的工作。
大战期间,我正在研究肺炎球菌的多糖及其抗体,也知道关于DNA是遗传物质的争论。但是,当时我未接受这种观点,因为我对蛋白质颇有好感,因此我认为遗传物质可能是蛋白质而不是核酸。当然,核酸是发挥了一定的作用,而且凡谈到核酸时,我总要提核蛋白,因此在蛋白质和核酸之间,我更多地想到蛋白质。
(波林,1968)
波林接着又说,他研究了多糖之类的许多种多聚物的结构,但都没有发表。像DNA结构一样,这些多聚物的结构也引起了他的浓厚兴趣,因此,虽然还不相信DNA是遗传物质,他还是“认为对阿斯特伯里拍摄的DNA照片作番研究是值得的”。当彼得·波林被问到同一个问题时,他说他父亲对“核酸感兴趣,因为核酸是种化合物,而不是因为核酸是遗传物质……”(彼得·波林,1967)。
那么,为什么正值1952年皇家学院的研究工作取得进展时,波林开始研究DNA呢?我相信,波林研究DNA与皇家学院的工作毫无关系,有关系的却是1951年11月爱德华·罗恩温发表在《美国化学会会志》上的一篇不寻常的论文《核酸的磷-3-酐基》。罗恩温论证说,仔细研究了核酸的结构和性质的数据后,他得出的结论是每个核苷酸只有一个磷-糖酯键。这一结论无视了莱文所作的权威性研究。因此,他提出的是磷酸盐骨架而不是糖-磷酸盐骨架。他提出的分子核心由磷-酐链组成,每个磷原子同5个氧原子连接(见图20.10)。他承认P2O5“与水剧烈反应”,但多聚形式的P2O5的反应却不是那么剧烈。托德向剑桥的马卡姆作了下述评论:如果把DNA放入水中会发出嘶嘶声,那么罗恩温的结构才可能是正确的!
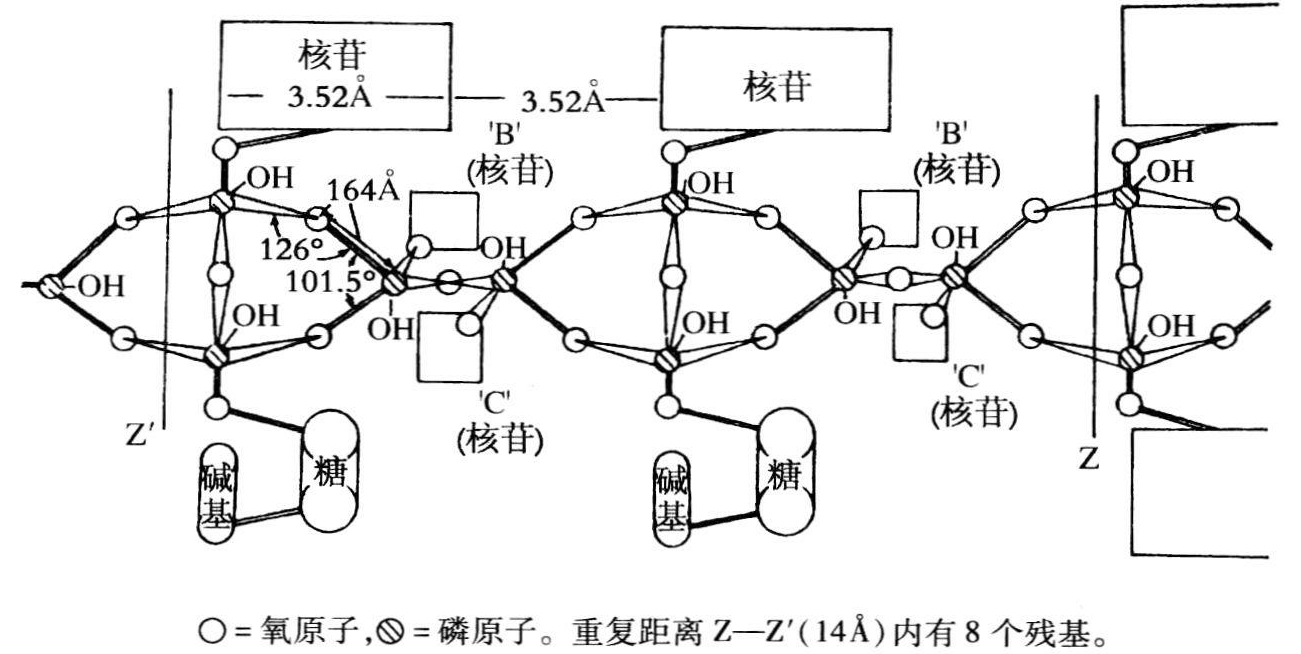
图20.10 核酸多聚物链的单位晶胞的顶视图(录自罗恩温,1951,5142)。
波林没有取笑这种结构,但“被这种胡说八道惹怒了”(舍马克,1968),因此他拉了舍马克一同投书该杂志,驳斥了这篇文章。
为核酸提出这种离奇结构的人,没有提出支持该论点的任何有意义的证据。每个磷原子与5个氧原子连接,从结构上来说是很靠不住的,因此为核酸提出的这种磷-3-酐式是不值得考虑的。
(波林和舍马克,1952a)
那年春天,罗恩温写信给波林,提请他注意化学家安许茨合成的四种化合物,安许茨也认为这些化合物的结构是磷原子与5个氧原子连接(安许茨,1927)。1952年6月,波林和舍马克不得不撤回了从未有过这种结构的说法。但是,他们提醒人们注意,安许茨合成的化合物对水是极为敏感的,其敏感程度已达到了“只能非常粗略地测定它们的熔点和其他物理性质”(波林和舍马克,1952b,3713)。
随着时间的流逝,1952年波林对DNA的兴趣也与日俱增。6月,他看到了沃森写给德布吕克的一封长信,信中谈到了在英国进行的DNA和TMV方面的研究工作。这封信也表明沃森希望在剑桥而不是在加州理工学院度过作为国立小儿麻痹症基金会研究员的第一年生活。德布吕克在给该基金会的信中写道:
波林博士和我本人已仔细研究了沃森博士的信件。我俩都同意他的计划和他继续在剑桥工作一年的理由。就沃森博士想要解决的那个问题来说,剑桥的技术设备确实要高级得多,至少在即将到来的一年里是这样。波林博士希望,到下一年我们这里将具有同样优良的设施。
(德布吕克,1952b)
因此,波林知道威尔金斯已获得“极好的X线衍射照片”。如果那年夏天美国政府没有吊销他的护照,使他能前往伦敦参加皇家学会召开的为期一天的蛋白质的讨论会,他很可能会看到皇家学院研究小组拍摄的那些照片。他的同事罗伯特·科里参加了那次会议,罗莎琳德·富兰克琳给他看了DNA照片。科里告诉波林皇家学院研究小组已拍摄了一些很清晰的照片。但是,当波林写信给威尔金斯要求提供详细情况时,威尔金斯的回复是他的研究工作还没有达到公布细节的阶段。
那年,波林在加州理工学院虽然也拍摄了一些清晰的DNA照片,但“及不上阿斯特伯里和贝尔的照片”(波林和科里,1953,85)。只是在波林发表了他的结构后,加州理工学院的博士后研究人员亚历山大·里奇拍摄了真正好的照片。1951年,利兹大学阿斯特伯里系里的E.贝登已得到了很好的B图型,波林对此几乎是一无所知,在1951年之前,还没有人对B图型作过什么研究。
阿斯特伯里已要贝登从事小牛胸腺DNA的X线分析,这些DNA是1951年阿斯特伯里向查格夫讨来的。贝登用了系里那台大功率的旋转阳极X射线管,曝光时间为15~20分钟。他使DNA材料拉长了300%,在拉长过程中,同时把水滴在DNA材料上,从而获得了很好的现在所说的B图型。放在房间湿度下的同样的样品使他得到了威尔金斯已发现的晶体图型。说来也怪,阿斯特伯里对这些结果深感失望;而且很明显,他和贝登都认为1938—1939年间贝尔拍摄的照片代表了纯净型,而贝登的B型则是混合型——当然X射线的点子要少些!
所以,在帕萨迪纳的波林只能用阿斯特伯里和贝尔在战前拍摄的照片,来检验他用随机法构成的结构的正确性。
波林的研究方案
波林从研究核苷酸的立体化学开始。托德由于发现了环化作用,刚刚搞清了糖磷键合问题(布朗、托德,1952)。1951年环核苷的形成为糖苷键的β构型提供了证据(克拉克、托德、楚斯曼,1951)。波林也知道富尔伯格研究胞苷的工作,得到了糖对碱基的正确取向。因此,他构成了反映所有这些特点的多核苷酸链。
接着,他论证像多肽一样,在多核苷酸中为了把不对称的亚单元(核苷酸)转变成对称结构的一般做法,是产生螺旋的旋转变换。
接着遇到的难题就是密度问题,我们已看到阿斯特伯里和贝尔没有很好地作出分析。波林也做了阿斯特伯里等所做的那种计算,得出的结论是沿分子长度方向,每3.4Å一定有三个核苷酸,因此是三条链。他从干的DNA密度(阿斯特伯里和贝尔给出的)算出了核苷酸的体积:
![]()
每个残基的横截面积为303Å2。因此,每个残基的长度为
![]()
或者说每3.4Å中有三个残基。“于是,反射将归因于由三个残基组成的一个单元。”(波林和科里,1953,86)因此,一定有三条链。
我现在还未搞清楚波林是如何得出每个核苷酸的横截面积的。他得到了罗布林·威廉斯的电子显微镜照片的证据——DNA分子纤丝的直径介于15~20Å之间(威廉斯,1952)。阿斯特伯里-贝尔照片的赤道点给出的最小侧距为16.2Å,根据波林的推算,这一数值相当于18.7Å的分子直径。如果我们用这个数值来计算该分子的横截面积,得到的值将是235Å2。于是,高3.4Å的碟状核苷酸的体积为935Å3。因此,该碟状体中的核苷酸数目为935/338=2.8。如果取贝尔论文提供的一般湿度下的DNA密度,我们算出核苷酸的数目为2.57。上述情况表明,要是波林当初看到了1939年利兹大学获得的全部数据,他也许会继续研究双股模型了!
正如波林记录本所表明的,他已开始研究单位体积里的双链结构,但他发现“根据计算的密度,还有很多多余的空间,在这个单位体积中可容纳三条链,双链只占去整个空间的三分之二……”(波林,1968)。因此,波林被迫放弃了双螺旋而提出三螺旋结构。十五年后,他说:“我当时倾向于双链结构,因为几年前我就断定基因包含两个互补分子,每个分子都可成为合成另一个分子复制品的模板。”(波林,1967)
然后,他又提出一个问题,该分子的核心的性质是什么?因为在这里它应是一条稳定的螺旋。原子堆砌在核心比堆砌在四周是更难解决的问题。他从研究多肽得到的教训是该结构中的“原子以相当紧密方式堆砌在轴的四周”,这种结构很可能是正确的(α-螺旋是正确的,γ-螺旋是错误的)。
经试验发现,由于嘌呤-嘧啶基团会发生变化,因此它们不可能沿螺旋轴堆砌,使得在糖残基与磷酸基团之间能形成合适的键;这样就否定了这种结构。同样,糖基团也不可能构成分子核心;核糖呋喃糖原基团和脱氧核糖呋喃糖基团的形状决定了这些基团难以沿螺旋轴紧密堆砌,此外,迄今还没有找到使这些基团堆砌的满意方法。多糖淀粉是说明很难获得紧密堆砌的一个例子。这种淀粉沿轴形成的螺旋有一个洞穴,其大小可容纳一些碘分子。我们的结论是分子核可能是由磷酸盐基团构成的。
(波林和科里,1953,89)
什么使结构结合在一起
关于这个问题,波林的态度是如果结构聚合得很好,其部分原因是满足了范得瓦尔距离,以及存在着比共价键弱的价力。不像沃森和克里克在1951年提出的结构,也不像阿斯特伯里的一堆硬币模型。波林没有理会碱基之间的静电引力,而是在磷酸基团之间引进了氢键。不管在什么层次上,三个核苷酸的磷酸基团由氢键相互连接,键的大小相当于磷酸二氢钾中的氢键。这种结构必需的质子将取决于未离子化的每个磷酸盐的主要磷酰基团。波林认为,虽然抽提的DNA就是阿斯特伯里和贝尔用来拍摄照片的钠盐,但是Na+离子十之八九不同主要磷酸基团毗邻。后来,他回忆说,他“本人已分析了所报道的一些滴定曲线,这样做是为了想确定核酸离子化的程度。我得出的结论是,在生理pH值时,大量的磷酸基团不会电离”(波林,1968)。这种说法是出人意料的,因为1953年的论文没有讨论过这个课题,以及所谈到的DNA的滴定曲线不是磷酸基团的解离而是氨基和酮基的解离。根据现有情况,唯一的结论是波林把DNA结构主要看作是磷酸化学的一个问题。扼要地说,在波林心目中问题的关键是把磷酸四面体聚合起来,从而忽视了其他问题,例如,碱基之间的堆垛力。尽管他没有按照那年富尔伯格提议的方法去做,他还是成功地用了糖环。因此,他很高兴地把碱基放在结构的外面(见图20.11)。三条链中的每一条都转了七圈(包含24个残基)后,回到相当于原来的位置上。因为螺旋中有三条链,经过8个残基后,沿螺旋27Å的地方出现结晶重复。1953年的那篇论文只字未提钠离子。
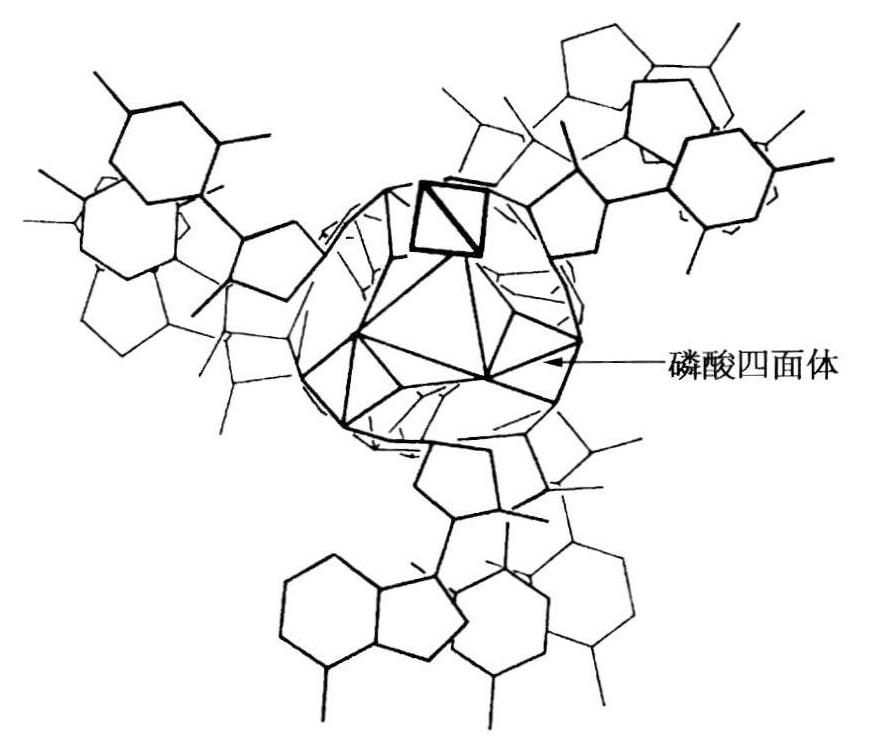
图20.11 波林的DNA结构图(俯视螺旋)。磷酸四面体由穿过中心的洞穴的氢键彼此连接(录自波林和科里1953,92)。
波林最关心的是这个结构在立体化学上是否站得住脚,同阿斯特伯里和贝尔拍摄的照片是否基本相符,同他们发表在《生物学图》(1939,95)上的、观察到的反射是否一致。根据科克伦、克里克和范德的螺旋理论作出的预测,同层线强度符合一致。波林以非常乐观的口气开始了他的沦文。这是“很有希望的结构……不是含糊不清而是可精确预测的,给出了主要原子的原子坐标……这是研究人员有史以来第一次精确阐明了核酸结构”(波林和科里,1953,84)。他以对生物功能的评论作为结尾:
有趣的是,位于分子外圈的嘌呤和嘧啶基团所占的位置,正好使它们的氢键形成的基团呈辐射状排列,从而使核酸分子能同其他分子剧烈反应。此外,每个氮碱基区都有宽裕的空间可任意选择任何基团;立体阻碍也不会干扰残基的随意排列,因此,所提议的结构允许构成最大数量的核酸,提供了高度特异性的可能性。正如阿斯特伯里已指出的,表明沿分子轴上相等距离的3.4Å线反射,正是伸展的多肽链上每个残基的近似长度;就这种大小来说,核酸同蛋白质中氨基酸残基的排列十分相称。氨基酸残基的位置很可能位于平行四边形的中央,而4个氮碱基占据了平行四边形的四角。256种不同的平行四边形(忽略不计每个氮基可能存在两种不同的取向)为每个位置的选定提供了很大的可能性。
(波林和科里,1953,96)
1952年12月底,波林寄走了他的手稿,但到了1953年2月,显然需要对该结构作修改。2月4日波林写信给儿子:
……我们已发现,对我们提出的核酸结构的原子坐标需略加修改。有一个范德瓦尔接触点太接近——介于碳原子5′同连接的磷酸基团的一个内部氮原子之间。我认为当时在作最后计算时,我忽视了这个接触点——在初始的结构里,有几个很小的范德瓦尔接触点,因此为了使它们达到能被接受的最起码的大小,我想方设法探求能提高所有这些接触点的参数。在最后阶段,我显然忘了把这个接触点包括进去。对这些参数重新作一检查将花费几个星期,但我预计这些数据将证明是正确的——至少我希望是如此。
(波林,1953a)
两星期后他写道:“我正在检查核酸结构,想使这些参数更精确些。我想,原来这些参数不是完全正确的。毫无疑问,这个结构几乎使全部原子受到紧紧的挤压。”(波林,1953b)
对这个结构的检查工作很快就完成了。正如舍马克所说的:“波林这个人的特点就是善于思考和坚韧不拔,因此总是能取得突破。接着他就迅速干起来,并准备了很周到的讨论会,把一切都告诉别人……”(舍马克,1968)在不可避免地要修改这个结构时,波林去征求舍马克的意见,因为他时常到舍马克那里去讨论DNA。很清楚,正是由于这些拜访,使DNA结构对波林来说变得“如此重要、如此奇妙。我想我对DNA结构有所了解……他已被DNA结构迷住了”(舍马克,1968)。
大约在发现了范德瓦尔距离上的错误以后三个星期,舍马克提出了一条改进意见。他建议使磷酸基团沿垂直轴旋转45°。在记有2月27日日期的笔记上,关于这个更改,波林写道:“OⅢ-OⅡ’= 4.26Å。很好。”早先对这个结构持怀疑态度的舍马克,现在也信服了。这个结构在各方面配合如此之好,因此一定是正确的。
弗雷泽螺旋
为建立三股螺旋又做过一次尝试,这就是红外线专家弗雷泽所进行的。他在1951年曾做成了改进的中央骨架单股富尔伯格模型。1952年,他做了一个三股模型,碱基之间由氢键连接,里面是螺旋。由于克里克反对把显然是错误的结构同他们的结构同时刊出,因此,在1953年没有发表这一模型,现在也不知道这个模型的具体情况。所以,即使在皇家学院——当今成果的发源地——1952年时的意见是支持三股模型。尽管这个模型不能说明纤维的对称性,也没有吸收查格夫比率,但在1952年冬,这个模型最接近正确的结构。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










