



第七章 基因的核蛋白理论
每条染色体的形态都明确而恒定,由许多片段组成;每一片段又各有特定的染色质线或宽带的图式……最初,这一发现使我们有了染色体分析的定性方法,当我们在研究已知其遗传特性的染色体重排而获知某一遗传因子的正常形态时,我们就能用前所未有的精确性来确定基因在染色体上的位置和绘出染色体图。
(佩因特,1933,585)
细胞遗传学引导我们去研究细胞力学。现在细胞力学迫使我们去推论作为其基础的结构。在探索遗传和变异的机制时,我们发现了生长和生殖的分子机制。细胞学说揭示了生命过程的统一性;研究细胞是揭示这些过程的物质基础的开端。
(达林顿,1937,562)
要用研究不太重要的生物分子的方法去研究染色体并由此而解决问题,看来还得等上几十年才会有这样的方法。因此,如果我们在斯德哥尔摩的同事们能提供蛋白质和核酸在有丝分裂周期中分布变化的资料,我们将张臂欢迎,因为这些资料是亟需的。
(林奇,1935,389)
哥本哈根会议(1938)取得巨大成功。参加第一次会后,我还没有料想到会取得这样的成功。整个小组显得很融洽,我们确实是在努力工作。包括物理学家在内的每一个人都十分热心。从我们正在草拟的报告中,你马上就会看到讨论的结果。我相信下一次会议将会是令人更感兴趣和更有效率。
(埃弗鲁西,1938)
米歇尔死后很长一段时间里一直在寻找细胞核和细胞质染色特异性的化学基础,但没有成功。在这方面起到如此重要作用的米歇尔非但没有拨开迷雾,反而进一步搞乱了思想。他宣称他没有观察到巴洛维茨、施威格-西德尔和拉范里特·圣乔治等人在精子中清楚地观察到的特异性。他称细胞学家为“染匠公会”并怀疑他们的工作(米歇尔,1897,i,107~108),假如他能活着看到阿尔弗里德·费希尔批评染色特异性的化学理论(1899),以及看到威廉·哈迪爵士认为原生质结构是固定产生的膺象(1899),那他一定会深表欢迎的。因此,在引进新技术以前,细胞化学未能对了解细胞中DNA的生物学功能作出实质性贡献是不足为怪的;特异性确实有其化学基础,它必定同DNA中糖环C2位置上的氧原子有关。这是胞核反应,1914年罗帕特·孚尔根设计这种反应来抽提核酸,九年后应用于组织。1923年在多宾根鉴定这一新技术时,使富有经验的核酸化学家阿尔勃里希特·科赛尔(参阅费利克斯,1957)留下深刻印象,但当这种技术发表时,细胞学家似乎并未采纳用于化学研究(孚尔根和罗森贝克,1924)。
1914年,孚尔根断言他的细胞核反应已涉及醛基的方位,他把醛基定位在糖环上。这样,他为鉴定DNA中的糖是脱氧核糖的工作铺平了道路。1923年,他开辟了一条途径以确定DNA和RNA在细胞里的分布,他还做了许多工作。几年后,卡斯帕森成功地使孚尔根反应定量化,可在细胞生命期间弄清细胞核的DNA含量。结果证实了长期以来的看法,即在一个合成周期以后随着是DNA分解;海登哈因在1907年就已作了描述。因为DNA合成同细胞分裂和染色体复制的开始相联系,所以修改了中心法则的蛋白质版本,赋予DNA以重要作用。现在都认为在合成基因的新蛋白质分子时,必须要有DNA。阿斯特伯里用“绷架”的形象指出了DNA的作用,一个基因现有的多肽链在“绷架”上保持伸展的形式,它可作为模板在上面制成完全相同的链。1931年,他看出倘若亲链和子链的走向相反,就可以实现这种同一性。所产生的有次序的结构将具有C2[1]对称性(阿斯特伯里,1931)。后来他提出DNA是作为两条“正链”——基因的多肽链和子基因的多肽链——之间的一条“负链”(阿斯特伯里,1939b)。非常奇怪的是,后一观点并没有提出核苷酸序列与相应基因的多肽的氨基酸序列相匹配的问题。
卡斯帕森和布拉歇以类似方法独自研究了RNA在细胞质蛋白质合成中的作用,再次认为在所有重要蛋白质的复制中,核酸起了“接生婆”的作用。因此,染色体和细胞质的复制基本上相同——两者都涉及多肽链的复制。关于DNA和蛋白质的作用的这一假说,我们可称之为“基因的核蛋白理论”。这种假说的力量在于DNA和蛋白质都起重要作用,遗传的特异性保持在多肽链的氨基酸序列之中。当艾弗里的工作宣布时,生物学家和化学家的思想深受这一理论的影响。本章将概述这个理论所依据的一些证据。
哈默斯坦和卡斯帕森
核酸在细胞核里周而复始地合成和分解,这一过程的最令人信服的证据来自斯德哥尔摩。在那里,哈默斯坦家族从奥洛夫·哈默斯坦那时起就同生理化学研究结下不解之缘。奥洛夫只有两篇有关核蛋白的论文,但他的侄子艾因纳·哈默斯坦在其兄——体操教员哈拉德——的帮助下,多年来致力于研究胸腺核酸的物理化学、黏度、对电解质和酸碱度的敏感性、对蛋白质的亲和性以及渗透压。在这一研究过程中,艾因纳·哈默斯坦改进了伊瓦尔·班的抽提方法,1924年时可把他得到的产物描绘成“雪白的,像强棉药一样有特殊的韧性”(哈默斯坦,1924,386)。因为在抽提时他一直使温度低于1℃,酸碱度保持中性;因为他避免使用碱和酸,所以得到了像白色纤维一样的胸腺核酸钠盐沉淀物。这项研究迥然不同于利文的工作。其实,利文说明了四核苷酸的结构,哈默斯坦揭示了降解程度较小的分子的物理性质;由此重新发现了五十年前米歇尔曾在鲑鱼核素中观察到的极度不稳定性。利文研究的是有机化学,也提出了与该学科有关的一些问题。哈默斯坦研究的是生理化学,米歇尔、科赛尔、纽曼和哈默斯坦的叔父奥尔夫等在这个领域中建立一种传统。遗憾的是艾因纳研究的也是胶体科学。因此,他没有把低渗透压归因于高分子量,而也是归因于高分子体积对低分子体积的阳离子效应,好似这些阳离子被大的分子吞入,从而隐藏起来不再对周围溶液发挥其影响。当他谈到高分子体积时,他指的是相对分子质量为1545,稍大于四核苷酸的分子量。
作为一位生理化学家,哈默斯坦想把他的发现同细胞核的生物学功能联系起来。这里他看出了核酸同水、酸碱度以及蛋白质之间的关系有重要意义。这也许可解释为什么细胞核有不同的酸度,而细胞质则是相当恒定的中性反应。他似乎属于贾克斯·洛布和威廉·哈迪爵士的“机械”学派,他们在电解质、水、胶粒中看到了揭示生命现象的关键。因此,DNA的黏度很大使他困惑不解,情况就像十九世纪时的米歇尔,他探求“乳酸在肌肉中的作用与核酸在细胞核中的作用之间的直接平行关系,两者对于水的涨落的不稳定性都有巨大影响”(哈默斯坦,1924,385)。
当DNA在溶液的酸碱度改变时,哈默斯坦确实现察到黏度的巨大变化,他认为原因在于改变了“水合”程度;水合作用越大,核酸分子对其水环境的影响越大,因此核酸溶液的黏度也就越大。我们已经知道,1938年在卡洛琳斯卡研究所用哈默斯坦-班制备的核酸已被看出不止是四核苷酸胶束或聚集体,而是一种大分子化合物。但是,这个结论最后是由斯德哥尔摩的科学家形成的。哈默斯坦开始考虑DNA的“巨大黏度”起因于对分子移动的某种“限制作用”,例如拉长的不对称分子会产生这种作用。1927年,龙斯特罗姆根据流动双折射研究得以向哈默斯坦证实这种不对称性;六年后,佩德森用斯维德伯格的超离心机重新检验了这个问题。虽然DNA的行为像一个多分散系统,在pH4和pH10之间得到的相对分子质量大约在50万~100万之间(许多年后.这个数字减至250 000)(斯维德伯格和佩德森,1940)。可是直到1939年,哈默斯坦根据这些结果还在颗粒、胶束或分子的意义之间动摇不定(哈默斯坦,1939)
此时,卡斯帕森正试图从他的核酸溶液中除去所有的浊度,以便在孚尔根染色后测定它们的透明度。过滤作用不是保留蛋白质并让核酸留在滤液中,而是保留核酸并让低分子量的蛋白质通过滤膜。卡斯帕森以困惑的语调写道:“核酸复合物一定大于蛋白质分子……分散的蛋白质颗粒的半径和核酸分子的半径之比,一定大大不同于开始时所预料的。”(卡斯帕森,1934,162)当他写“复合物”时,他已开始想到这可能是种高聚物。此时,他用光学方法定量测定细胞核反应导致他朝着另一重要方向前进。
核酸循环
1932年前后,卡斯帕森试着用紫外线照射核酸溶液,在照相底片上记录下透射光谱。显影时在2600Å处出现一个明显的吸收峰(见图7.1和7.2)。由于不了解以前的报道,卡斯帕森很惊奇,但查阅文献以后,发现有很多论文有关核酸、核蛋白、腺嘌呤、鸟嘌呤、尿嘧啶和胞嘧啶的紫外吸收(索雷特,1883;戴雷,1906;霍利德,1930;海洛思和卢夫波洛,1931 ;以及其他作者)。因此,核酸的这种性质同孚尔根的细胞核反应结合起来,提供一种方法去定量计算细胞里的胸腺核酸和核糖核酸。
瑞典的研究组有很多理由可作出这一发现,因为他们对核酸和蛋白质之间的平衡发生改变的生理意义感兴趣。染色质也是一个恼人的问题:只有当细胞开始分裂时,染色质才表现为一条条棒状的染色体。在所谓的休止期,它们分散成一大团球状物质,很难染上颜色。博维里确定染色体通道连续的休止期仍保持个体性,他提出,在休止期染色体的永久结构失去嗜碱物质(染色质),在开始分裂时又重新获得(博维里,1888)。这种“非染色质”假说得到海克、蒙哥马利、马修斯和斯特拉斯伯格的支持。斯特拉斯伯格在谈到染色质时说:
这种物质汇集在染色粒里,也许构成了遗传单元载体的营养物,我们现在认为遗传单元包含在染色粒里。染色质本身不可能是遗传物质,因为它以后离开染色体,在发育不同阶段它在细胞核里的数量有相当大的变动。 同参与形成细胞核纺锤丝和细胞里其他过程的物质结合在一起,染色质在休止细胞核里积累起来以形成核仁。
(斯特拉斯伯格,1909,108;引自米尔斯基,1968,88)
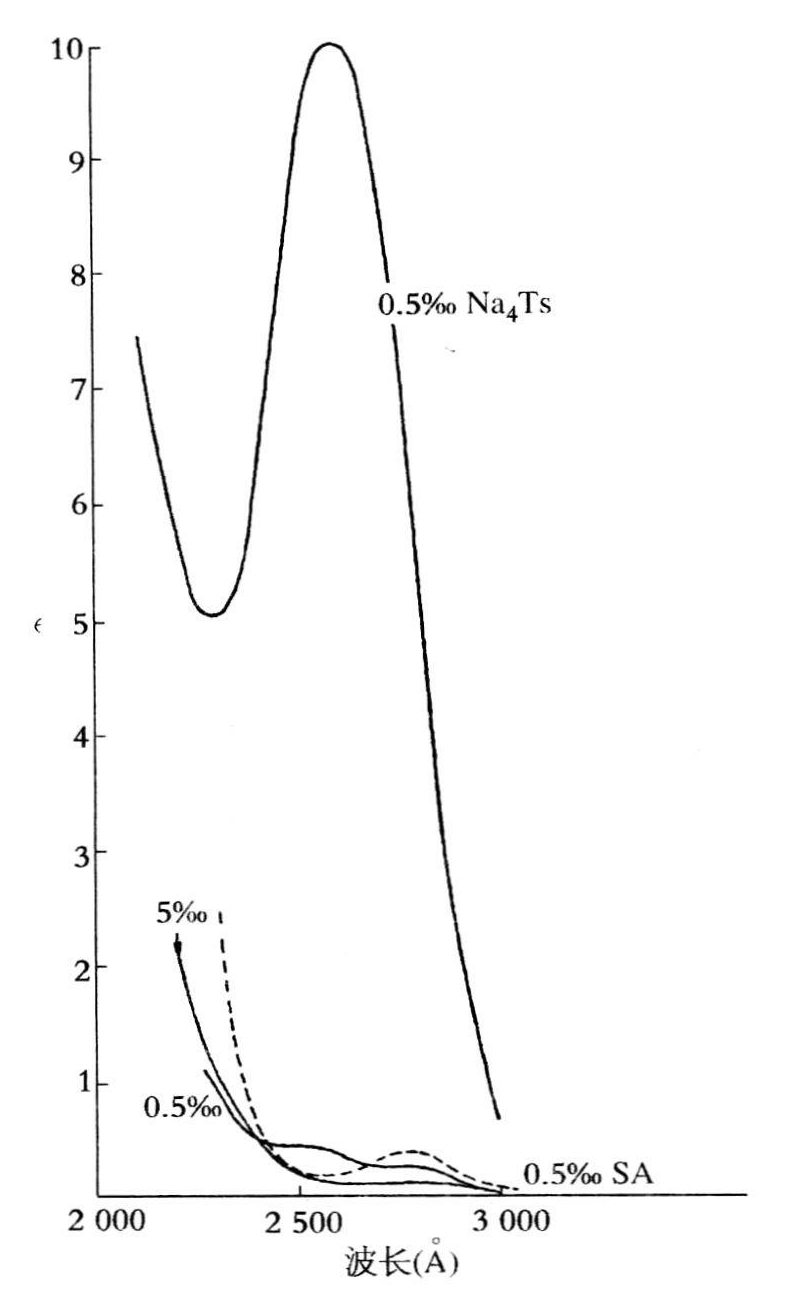
图7.1 NaDNA(用Na4Ts标记)溶液,血清白蛋白(SA),硫酸鱼精蛋白和硫酸组蛋白(底部的实线)的吸收曲线,最上面的曲线是0.05%胸腺核酸钠,虚线是0.05%血清白蛋白,实线是0.5%硫酸鱼精蛋白,底部曲线是0.05%硫酸组蛋白(录自卡斯帕森,1936,32)。
E.B.威尔逊也站在斯特拉斯伯格一边,在很有影响的《发育和遗传中的细胞》一书中,他写道:
“染色质”(即碱性染色质)的连续出现是染色体遗传连续性所必不可少的,可是,这已成为过时的见解……染色反应表明,长期保持的不是嗜碱成分(核酸),而是所谓的“非染色”物质或嗜氧物质。核酸成分在细胞活性的不同时期时有时无……
(威尔逊,1925,895,653)
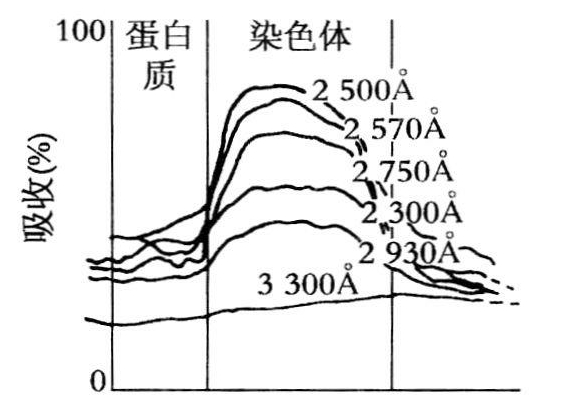
注:卡斯帕森的图上2300Å显然是2800Å之间。
图7.2 染色体显微切片在6个波长处的吸收曲线,该染色体的两侧都有蛋白质(录自卡斯帕森,1936,94)。
1934年6月,T.H.摩尔根在斯德哥尔摩发表他的诺贝尔奖获奖演说时提出了一个问题:“基因是什么?”
现在我们把基因落实在染色体上,我们有理由把它们看作是物质单位,看作是比分子更高级的化学实体吗?坦率地说,这些是遗传学家还没有很好关心的问题,而只是对假设的因子的性质不时作出猜测。关于基因是什么——是真实的还是纯属虚构的——遗传学家莫衷一词,因为在遗传学实验层面上,不管基因是一种假设单位还是物质颗粒,都是差不多的。不管是哪一种情况,单位总是同特定的染色体相联系着,因而可以通过纯粹的遗传学分析来定位。因此,如果基因是一个物质单位,那它是一段染色体;如果基因是一个虚拟单位,也一定可通过推理确定它在染色体上的位置——就在按其他假设得出的同一位置上。所以,它在任何一种观点的遗传学实际研究中都没有两样。
在遗传学家所用的性状同遗传学家的理论假设的基因之间,是胚胎发育的整个领域……
(摩尔根,1933,315)
人们必须记住,在那时,还很少有证据表明沿着染色体纵长存在化学上的分化。不过当时确已认识到染色体是一串深染的染色粒,由称为“核丝”所组成的淡染的线串在一起。可是,遗传学家对这些模糊差别的化学基础却缺少热情。当E.海茨把地钱的染色质区分成形态上完全不同的异染色质和常染色质两类时,遗传学家的态度开始发生改变(海茨,1928),常染色质在有丝分裂结束时失去着色性,在此以后很长一段时间内异染色质仍可着色。后来,他把这一发现推广到果蝇(海茨,1933)。T.S.佩因特发现果蝇唾腺染色体上的带纹是染色体的恒定的特征,这一发现更引起了遗传学家的注意[2]。
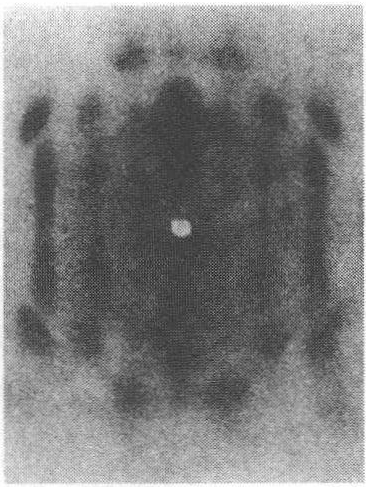
在X染色体的常染色质中(见图7.3),他画出了现在了解得已很清楚的150多条带纹,画出了断裂点位置以及染色体纵长上的基因座位(佩因特,1933)。关于这一工作,斯蒂特文特说:
这里,最后是在交换图和细胞学上可见标记之间的详细对应关系,以及一种根据可见带纹精确确定基因座位的技术。每条染色体上的标记不再只有两三个(端部和着丝粒),而是有几百个,对于整个染色体组来说很快就会有几千个标记。
(斯蒂特文特,1965,76)

图7.3 果蝇X染色体的照片简图。唾腺经醋酸洋红处理后,由佩因特拍摄(录自佩因特,1933,586)。
如果确定了基因在染色体上的特定位置,下一步就将是分析这些染色体区段的化学组成。把遗传学家的抽象单位同化学家的有机化合物联系起来,现在看来是值得一试的。这不止是另一个发现。这是把遗传学家和细胞学家的发现结合起来的一个里程碑。而且,这是开辟了从化学上探究染色体精细结构的道路。
摩尔根在佩因特的工作发表之前,在1934年发表的诺贝尔奖获奖演说中指出,他在著名的果蝇唾腺染色体的切片中证实了经强醋酸处理的物质出现了横纹。人们不难想象哈默斯坦和卡斯帕森对摩尔根的切片是何等地感兴趣。斯德哥尔摩的研究组搞出了一种试剂可消化染色体的蛋白质,把核酸固定成一种镧盐而保留下来。他们用这种酶-镧试剂去研究“细胞核里特别是染色体里的蛋白质结构”(卡斯帕森、哈默斯坦和哈默斯坦,1935,367)。他们计划用紫外显微分光光度计来显示该试剂处理前后紫外吸收图式的变化。不出现任何明显的变化,使他们怀疑到在他们所用材料中,蛋白质像是一块块地填在核酸条纹之间,但他们预期在摩尔根的材料中将会证实这种排列(同上,369)。他们认为,强醋酸掩盖了染色体纵长上蛋白质浓度的“不着色”,深色带纹代表的是低蛋白质含量和高核酸含量。
当哈默斯坦和卡斯帕森于1934年向伦敦法拉第学会提出报告时,他们的工作还处于早期阶段。他们用的紫外显微镜很不完美,以镁在280mµ处的双线为光源。后来,卡斯帕森大大改进了这些显微分光光度计的质量,采用镉光源,他通过一个单色光器产生257mµ——紫外光谱中核酸吸收最强的部分。蔡司紫外光显微镜的设计师奥古斯特·科勒,于1904年已鼓吹采用镉,当时他发表了用这种光源得到的照片,这些照片中最令人惊奇的是蝾螈幼虫鳃芽细胞的照片。他把紫外显微光度测定技术,看作是传统染色技术所得结果的对照,并作为开辟了一个新的研究领域(科勒,1904,302)。卡斯帕森也许是在1933年把切片交给耶拿的科勒,他已看出可用那里的蔡司仪器设备完成那些工作。
科勒的玩意终于证明是有用的。希望在275mµ处使分辨力提高一倍,未能得到实现。早年试图用显微镜去研究细菌细胞核未获成功(施罗特,1906),卡尔·蔡司用这种仪器作过一次示范(1905)。但当卡斯帕森用科勒改进的单色光器研究唾腺染色体时,却揭示出那么多的细节!瑞典科学家在细胞化学领域开辟了一个新天地。

2.石蜡的衍射图(录自弗里德里希,1913)。

3.氟化锂粉末的衍射图(图a和图b),无定形硫的衍射图(图c)(德贝和谢勒,1916)。

4.纤维素纤维的衍射图(录自波拉尼,1921,339)。
5.纤维素(大麻)纤维的衍射图(录自阿斯特伯里,1940,26)。

6.丝的纤维的衍射图(录自阿斯特伯里,1940,22;阿斯特伯里,1933,79)。

7.阿斯特伯里——英国X射线晶体学的“约翰牛”(左),右面是H.J.伍兹。

8.α和β角蛋白的X射线图(录自阿斯特伯里,1930,图1a,1b)。

9.阿斯特伯里提出的α和β角蛋白中分子链的弹子模型(录自阿斯特伯里,1930,图2a,2b)。

10.胸腺核酸的X射线图——乳胶片比原来长度伸长1.67倍(大部分图式是A型所特有,但在3.34Å处的子午线反射表明B型的存在)(录自贝尔,1939,46)。

11.NaDNA和鲱精蛋白溶液界面上形成的干而卷的膜的X射线图(可以清楚地看到B型十字线特性和子午线反射,1947年复制了这种图式的一半,沃森和克里克作了研究)(录自贝尔,1939,66)。

12.紫外线吸收照相揭示的摇蚊染色体的带纹(录自卡斯帕森,1936,图92)。
13.多聚苯甲基谷氨酸伸展薄膜的X射线图型(录自班福德等,1951,图3,图7)。

14.J.D.沃森同特雷西·索恩本在一起。

15.X射线图,上面一半是NaDNA,下面一半是鲱精蛋白胸腺核酸盐(沃森和克里克特别注意后者)(阿斯特伯里,1947,图1,图2b)。

16.胸腺DNA的X射线图(查加夫提供,1951年5月,贝顿摄影)。薄膜比原来的长度伸展了3倍,R.H.=98%。这是A型和B型的混合物。

17.同一样品在98%R.H.放松几天并再伸展后的X射线图。这是非常好的B图式,以前未发表过,因此沃森、克里克和波林都不知道。

18.伦敦国王学院制备的小牛胸腺DNA的“片层”图式(材料由M.J.弗雷泽制备,戈斯林摄影),它显示了“十字线”,在3.4Å上的子午线和在这个角度里沿着子午线的虚假强度。

19.1950年6月戈斯林拍摄的“晶体”DNA的X射线图。这是单相图的第一个例子,显示A型产生的大量数据。

20.乌贼精子的X射线图,表明B图式的特性(威尔金斯摄于1951/1952冬)。

21.大肠杆菌DNA的X射线图,威尔金斯在同第20号照片差不多的时间拍摄的一幅很好的B图式。

22.富兰克琳和戈斯林的第一张B图式(R.H.=92%,几根细纤丝曝光6小时)。

23.富兰克琳著名的1952年5月的B图式(R.H.=75%,单根纤丝直径=50微米,曝光62小时)。

24.表示“双向”的图式(R.H.= 75%,单根纤丝直径=40微米,曝光100小时)。

25.A图式,显示在第十一层线上的子午圈角(用倾斜照相机拍摄)。

26.沃森和克里克在凯文迪什实验室他们制作的模型旁(1953年5月)。
卡斯帕森并不是三十年代里第一个或唯一的使用紫外显微镜的。拉尔夫·威科夫在洛克菲勒以及穆勒和普罗科菲叶娃在俄国都用过这种显微镜,但卡斯帕森第一个看出了染色体吸收紫外线和核酸吸收紫外线之间的全部重要联系。所以,用紫外摄影奇妙地观察到活细胞中的固定细胞中的染色体,出现了佩因特特别指出的带型。这是一种表现为遗传图的化学形态学吗?可能是如此。
谈了这一切以后,强调了遗传图同核酸分布之间联系的重要性以后,对于哈默斯坦和卡斯帕森继续维护基因一定是蛋白质的传统假设,我们难以苟同。他们在提到蛋白质和核酸为“已知物质”时写道:“只有蛋白质可加以考虑,因为蛋白质是有个体特异性的唯一已知物质。根据那个假设,染色体中的蛋白质结构令人最感兴趣。”(卡斯帕森、哈默斯坦和哈默斯坦,1935,369)因此,他们后来虽发现染色体复制时核酸合成旺盛却置之不顾,以维持中心法则的蛋白质版本。这就产生了我们可称之为“基因的核蛋白理论”。
细胞分裂开始时DNA的合成
卡斯帕森观察到减数分裂期间紫外吸收明显增加,他毫不怀疑地认为:“在早细线期,细胞核里很难检出有什么吸收物质;后来一定进行了核酸合成,最后发育出核酸含量丰富的完整四分体。”(卡斯帕森,1936,120)间期和接下去的前期,都同吸收物质的失去和补充有关。吸收物质是核酸;因此,卡斯帕森可能认为这一物质对染色体复制、从而对基因复制都是必要的。
当杰克·舒尔茨在斯德哥尔摩同卡斯帕森一起工作时,在果蝇唾腺染色体某些横纹的核酸含量同穆勒发现的、并认为起因于“染色体替代”(chromosomal displacements)的各种眼色突变种之间,发现了一种相关(穆勒,1930,299)。斯德哥尔摩的紫外测定表明“某一横纹越靠近异染色质区,它的核酸含量增加越多”(卡斯帕森和舒尔茨,1938,295)。当研究XX雌体的卵改为研究XXY雌体的卵时,注意到核酸也有相应增加。卡斯帕森和舒尔茨根据对唾腺染色体的研究正确地得出结论:异染色质并非是不活动区。他们以为染色体上的这些区所完成的:
……也就是别的基因完成的功能——即合成核酸。这些区同眼色多样化的关系,连同核酸出现在染色体的一定部位,提示了核酸合成同基因复制密切相关。胸腺核酸形成结构的性质,以及它形成高分子量聚合物的能力,再加上它的X射线衍射图同蛋白质的衍射图符合一致,都表明了这种功能的基础。
对于其他一些自复制的分子——病毒和噬菌体——这些理由特别令人感兴趣,因为这些分子都含有核酸。 另外,用紫外线灭活噬菌体时,不同波长的效率曲线同噬菌体的吸收曲线并不完全一致,但同核酸的吸收光谱则是符合的。因此,通过连续的聚合和解聚作用来调节活性和自复制的唯一结构,也许取决于该分子的核酸部分。蛋白质自我复制的性质,可能在于它合成核酸的能力。
(卡斯帕森和舒尔茨,1938,295)
当我们读到这一段时,我们感到很振奋。他们将作出结论:基因就是核酸!可是不,他们却背道而驰了,在最后一句话中我们看出了他们的内心想法。在两年时间里,卡斯帕森在广泛了解细胞里蛋白质代谢的基础上,确实已足以得出这一结论。在这里,细胞质里的蛋白质合成等于细胞核里的染色体复制。两者都涉及蛋白质分子的自复制要有核酸存在。卡斯帕森并未到此为止。他概述了细胞核的种系发展,从简单的含RNA的植物病毒,到含DNA的酵母和细菌,最后到高等生物含染色体的细胞核。比较原始的遗传物质定位在异染色质,它在那里产生组蛋白。组蛋白积聚在核仁里,最后转移到细胞质蛋白质中;这一幅简图,看来合乎卡斯帕森对分泌细胞的核酸含量所作的观察。
如果只根据我们现在了解的情况,那就难以理解卡斯帕森所勾画的简图;但我们可看到为了区分常染色质和异染色质的功能,他并未认为所有蛋白质都依赖于特殊物质从细胞核到细胞质的转移。好似大部分结构基因在没有RNA的情况下在细胞核里合成它们的蛋白质产物;少部分基因则是通过涉及细胞质RNA的更复杂的途径,才做到这一点。不管怎样,一个事实是很突出的——基因被认为是蛋白质,不管是病毒、细菌、真菌、高等植物还是动物。
基因是蛋白质的证据
就我所知,卡斯帕森从来没有说过在间期内,染色体中已除去了所有核酸。但由于只能检出极少量的核酸促使卡斯帕森排除核酸是遗传物质;不过在整个三十年代和四十年代中更有影响的却是这样的信条,即核酸结构远远不够多样化。他成功地证明了唾腺染色体纵长上全有蛋白质,在横纹之间的区段里则查不到核酸,这个观察也许鼓励了他的正统观念。他在1936年的那篇著名论文中写道:
构成大横纹之间或横纹复合区之间那部分染色体的蛋白质成分,很容易被消化和只吸收极少量的紫外线。在这里,机械因素和机械处理很容易打断染色体的连续性。用紫外线看不到有任何结构。
在横纹中与核酸紧密混合的也有蛋白质成分。经过长时间连续消化后,这些蛋白质分解了。
如果把基因想象为化学物质,那么只有一类化合物可以符合条件,那就是蛋白质,因为蛋白质可为变异提供无穷无尽的可能性。已表明两种染色体区段都含有蛋白质。
确定基因在染色体上的位置时,那些因核酸结构而有高度组织的横纹更可能是基因的载体,而横纹之间分化程度较低的非吸收区段不大可能是基因载体。
如果是这种情况,那么核酸的作用很可能是一种决定结构的支持物质。
(卡斯帕森,1936,138)
一般认为,对数据资料的解释应尽可能多地同有关领域的资料符合一致,通常是使解释合乎已经建立的框架或规范。卡斯帕森勾画的简图是核酸在基因复制中起监督的和非专一的作用,这同研究染色体结构揭示的染色体复制方式相吻合。在这里,所有的讨论都是以下列假设为基础:染色体用来汇集亚单位以生成其自身复制品的引力,也就是在减数分裂时把相同染色体——同源染色体——拉到一起的拉力。因此,人们感到研究染色体的机械运动,可用来弄清基因复制的复杂过程。
染色体机械运动
减数分裂和有丝分裂中染色体的行为明显不同,这是细胞学揭示出的最令人感到奇怪的悖理。在减数分裂时,所有染色体都彼此相吸引,配合成对或成为双价休,然后分裂成染色单体;但在有丝分裂时,染色体彼此相斥,而且当它们的形状刚刚看得清楚时马上就观察到纵裂成染色单体。就整个来说,细胞学家满足于用目的论的方式来解释这一悖理,即他们认为染色体在减数分裂中的配对或联会是有性过程的最后阶段;通过有性过程,父方和母方的染色体最后结合在一起,进行物质交换后彼此分离。减数分裂形成4个细胞而不是2个细胞,这被认为是出于一种需要:受精以前染色体数目要减半。
正确描述有丝分裂中的事件和正确理解有丝分裂中所看到的染色体怎样从间期核里产生出来,对于历史学家来说最为重要的是区分这两者。博维里对染色体个体性的全部工作,还不足以澄清连续的染色质纽这种错误概念;这种概念认为染色质纽分段而生成前期细胞核里的染色体。另一本细胞学史的名著完全忽视了这一点(休斯,1959)。假如没有对染色质纽概念的信奉,可能会更快地了解减数分裂的事件。那些相信这种概念的人,简单地假设在减数分裂中染色质纽分段的数目为有丝分裂中生成的染色质线的数目之半,并以此来解释减数分裂中染色体数目减半,以及有丝分裂时染色体数目相等(减数分裂和均等分裂)。当更详细地弄清减数分裂的细节时,染色质纽学说遇到了困难,因而又提出一些错误的过程去拯救这个学说,如端部配对或“端部联会”等。研究月见草的多倍体和环状染色体联合时采用了端部联会解释,造成了更大的混乱。一位年轻的细胞学家C.D.达林顿澄清了这种混乱局面,提出了一个简明的机械论图式,包括了减数分裂和有丝分裂,二倍体和多倍体,环状染色体,正常染色体和落后的或不配对的染色体。
当十九岁的达林顿来到伦敦郊外的默顿时,他有一张农学学位证书,向约翰·莫尼斯研究所的第一任所长威廉·贝特森申请安排工作。虽然贝特森总是要雇用牛津大学和剑桥大学的毕业生,但他还是被说服让这个伦敦的毕业生在研究所里无偿劳动三个月,接着又是三个月不领工资,最后于1924年让达林顿作为选修研究生,达林顿拒绝了贝特森要他去皇家棉花育种研究联合会工作的建议;当1926年这位孟德尔主义的伟大宣传家去世时,达林顿留下来在丹尼尔·霍尔手下工作,最后于1933年接替了他。
减数分裂的早成学说
1931年达林顿提出一个观点,认为减数分裂是有丝分裂的一种异常形式。在有丝分裂前期,每条染色体已分成两条染色单体。他认为的从染色体释放出的引力,此时被姐妹染色单体的赫附所抵消。姐妹染色单体缩短,彼此盘绕,变得更易染色,但整条染色体并不相互吸引;其实染色体之间好似有一种斥力,所以在中期赤道板上染色体之间有一定距离。另一方面,在减数分裂中,第一次分裂的前期开始于染色体分裂为染色单体之前:它是提前早成的,引力只能完成染色体之间的配对。“因此有可能把减数分裂前期未分裂的同源染色体的配对,看作是被它们提前收缩所打乱的状态的重建。”(达林顿,1931,234)根据这一说法,我们可预期同源染色体一旦分裂为染色单体就会分开。事实是它们仍保持联会,达林顿认为这是由于染色单体之间有一部分发生了交换(达林顿,1929)。
分子螺旋
早成学说自然引起了许多争论,达林顿的《细胞学的新进展》(1932)一书就是如此,用霍尔丹的一段话来说:“该书是无数抨击的对象。事实上,1932年第六届国际遗传学大会有一次会议主要就是批驳达林顿博士的结论。”(霍尔丹,1937,vi)。在这里我们关心的不是早成学说的证据,而是这个学说对细胞学中的还原论研究方法的促进和推动。
达林顿有意识地按照牛顿力学的例子,用特定的引力和斥力来解释染色体的移动;如配对染色体在分裂之前彼此之问的特殊引力,在分裂之后染色单体之间的特殊引力;不同的配对染色单体之间的非专一性斥力。他引用牛顿的力学,就无须去规定他所假设的力的精确性质和原因。达林顿以下述语句结束了他对染色体运动的分析:
“对于引力定律的作者,我们还不能信赖吗?”(1932,352)可是,达林顿还是在探索使染色体盘旋和解旋的这些力的性质。他指出,染色体的拉紧不是由于T.H.摩尔根所认为的纵长上的弯曲,而是由于交叉十字形的扭曲(1932,289)。达林顿问道,染色体为什么要盘旋呢?他开始区分出两种盘旋,一种是一条染色单体围绕另一条染色单体的盘旋(相关螺旋),另一种是一条染色单体围绕自身的轴盘旋(内螺旋)(见图7.4)。他观察到细胞开始下一次分裂之前还有一些螺旋未解开(残留螺旋),他也观察了新出现的螺旋。在最后分析中他得出结论说,造成可见螺旋的扭力,一定是由看不见的分子螺旋的生成所引起。他看出染色体中连续盘旋的程度,正是克服染色单体生成时所产生的扭力的方法;因为如果连续盘旋的方向相反,就可达到平衡。他写道:“所有旋转作用的目的,就是产生粗线期的平衡。”(达林顿,1937,549)饶有兴趣地看到这个观点发展为解释DNA合成中的“解旋问题”(珀森和铃木,1968)。

图7.4 两条染色体配对关系的示意图,表明染色单体的内螺旋,姐妹染色单体的相关螺旋,同源染色体的相关螺旋(录自达林顿,1937,549)。
螺旋化的化学基础
值得注意的是,直到1939年达林顿还没有讨论过染色体的化学。但他确实注意到了染色体盘旋程度与对核染色的亲和性之间的平行关系(1937,307),以及低温对于巴黎染色体[3]上某些区段染色强度的影响(达林顿和拉科,1938)。可是直到1939年12月,他才写信告诉阿斯特伯里,他发现了“核酸的差别”(达林顿,1939),并同拉科一起写了《百合染色体中核酸的匮乏》。在这篇论文中,在《核酸的供需》小标题下讨论了核酸的循环(达林顿和拉科,1940,201)。1938年到1940年间发生了什么事情呢?1939年8月,他参加在爱丁堡举行的第七届国际遗传学会议,卡斯帕森虽未出席,但他那篇1936年的重要论文《细胞核结构的化学组成》很快就到了达林顿的手里,并产生了深远的影响:
我终于能拜读您的1936年的大作。我是向科勒借来的,他是在国际遗传学会议上向德梅雷茨借的。在看到这篇论文之前,我并未注意到它的重要意义。拉科和我现已作出的一些发现,同您的发现十分相似。我们发现巴黎染色体上的不同区段……在休止期带有过量的核酸,而在中期时则核酸含量减少。
(达林顿,1940)
达林顿对卡斯帕森的工作深感兴趣的结果是在1940年到1947年间,不断发表有关核酸代谢同细胞遗传学之间关系的论文,论文以如下论述开始:
在休止期间,细胞核里的核酸含量较少,大部分不在染色体上。核酸的最简单的单位的相对分子质量至少是1300(古兰德,1938),由这些单位构成的核酸也许以聚合物的形式存在于细胞核里。在有丝分裂期间,核酸的量增加,并附着在染色体的蛋白质框架上(卡斯帕森,1937)。核酸可能通过核酶的消化而完整地离开框架(梅齐亚和耶格,1939)。但核酸不在螺旋化的染色体的表面上,而是同核酸自身的螺旋盘曲密切有关……
(达林顿和拉科,1940,201~202)
在上一年八月,他在爱丁堡知道了阿斯特伯里的论文《与基因问题有关的蛋白质和病毒研究》,在这篇论文中,阿斯特伯里强调了核苷酸和氨基酸彼此之间的距离都是3.34Å,认为这对回答下列问题是很重要的:“……为什么选中蛋白质和核酸为细胞核分裂中的反应物……”(阿斯特伯里,1939,50)达林顿又进了一步提出:“核酸是由排列在多肽链上的基因的作用而形成的,而且附在基因上作为基因的复制、分离甚至螺旋化的一个因子。”(达林顿和拉科,1940,202)
这样推广卡斯帕森和达林顿的工作,目的在于找到使化学进入细胞遗传学的通路。从二十世纪三十年代起,细胞化学得到前所未有的普及。虽然仍有人否认通过孚尔根染色确定核酸位于染色体上的证据(斯特德曼和斯特德曼,1947),虽然有关这个反应的化学专一性的争论沉寂下去了,但一方面确定了在异染色质区同核酸含量之间的联系,另一方面确定了基因作用同基因在异染色质中位置之间的联系,从而使化学同细胞学结合起来。在这一发展中最有效的工具是绘制染色体图,孚尔根染色和紫外显微镜。最起作用的材料是双翅目昆虫唾腺细胞的染色体组。不幸的是孚尔根的细胞核反应未能满足一些人的要求,他们希望获得比有争议的化学特异性试验所提供的化学特性更为可靠的依据。因此,有人写信给达林顿促使他不要介入关于常染色质和异染色质的化学同一性问题,并说卡斯帕森缺少确凿证据表明他实际上研究的是核酸(卡尔,1947?)。幸而没有持保留态度,因而与核酸的联系加强了。
基因复制
由于某些明显的理由,达林顿的早成学说很快使美国遗传学家H.J.穆勒感兴趣。回溯到1917年穆勒读到L.T.特罗兰的著名论文《生物学之谜和酶作用学说》时就已考虑基因复制的性质,第九章将讨论这一情况。特罗兰的基因复制概念是把基因复制作为一种生长方式,即基因的亚单位通过“力场图式”的相似性而被吸引到基因上,四年后这一概念使穆勒认为所谈到的这种力,也就是在减数分裂开始对把同源染色体拉到一起的力:“显然正是同样的力使基因生长和使相似的基因彼此吸引,但引力更强,因为在这里,基因不同部分的所有单个引力都相加在一起了。”(穆勒,1922,38)于是在1921年的多伦多演说中,我们发现了把自动催化引力和联会引力作了很有影响但却是错误的类比,他明确地说:“如果这两种现象都依赖于基因组成的共同原理,则研究其中一种现象取得的进展必将有助于解决另一现象。”(同上)他从三倍体同源染色体配对时要排除第三条染色体的现象中,找到了联会吸引的极性证据。达林顿推广并修正了贝林的研究,加强了穆勒的观点,英国细胞学家关于配对引力的假设,同穆勒想通过物理学家来解决基因问题的热忱不可能协调一致。穆勒认为这些力的作用“超过了显微镜下可看到的距离”(1936,211),“其距离远远大于物理科学已知的所谓内聚力、黏着力和吸附力等的作用距离”(1922,37)。他催促物理学家“去探讨物理学的可能性并告诉我们这是些什么样的力,以及如何产生的……”(1936,211)。当他结束这篇不合实际的讲演时,他面对细胞学家所揭示的一些谜——位置效应、联会、染色体相斥、盘旋和解旋——他承认:“遗传学家本身是无法独自进一步去分析这些性质的。一定要有物理学家和化学家的介入。谁愿意自愿从事这方面的工作?”(穆勒,1936,214)
在穆勒访问物理学家时,J.B.S.霍尔丹访问了化学家,他深受阿奇博尔穗·加罗德爵士的生化个性概念的影响,他是从F.G.霍普金斯处很好地了解这个概念,他给霍普金斯纪念文集撰写文章的题目是个体的生物化学。文章讨论了基因的复制。他论证说,这不可能仅仅是生长接着是分裂。
相反,这必定是一种“拷贝”过程。被看作是一个分子的基因,必定要展开成为厚达一个构造单位的薄层。否则它不可能被复制。最有可能的复制方法是通过类似结晶的过程,第二层类似的构造单位铺在第一层上。但我们可设想这个过程类似于以一张也许与原版片有关的“负片”作过渡而复制唱片,就像抗体同抗原的关系一样……
蛋白质合成问题是生物化学的一块处女地。这是一个极其基本的问题。如果遗传学所做的只是以最尖锐的方式提出问题,那么这个问题对生化研究仍是一个有价值的刺激剂。
(霍尔丹,1937,8~9)
克兰本博会议
洛克菲勒基金会对于把物理学应用于生物学的研究,采取了积极支持的方针,并曾出资召开了3次会议。与会的物理学家和生物学家讨论了有关基因和染色体结构的问题。1936年,在哥本哈根召开的首次会议上,讨论了X射线引起的突变问题及靶子学说。第二次会议是1938年4月在哥本哈根附近的克兰本博举行的。会上研讨了染色体的结构、分裂及接合。第三次会议是1938年10月在比利时的斯帕举行的,这是第二次会议的继续。克兰本博会议上的讨论意见在达林顿、鲍尔的报告中,以及沃丁顿准备的总结里都有记述。沃丁顿总结报告的摘要,发表在1969年《自然》杂志上。1939年,沃丁顿的《现代遗传学导论》、达林顿的《遗传系统的进化》,以及1939年阿斯特伯里在爱丁堡会议上发表的论文——《有关基因问题的蛋白质及病毒研究》和1940年贝尔纳的《细胞及原生质》等文章中,都讲到这些意见。
正如沃丁顿所指出的,克兰本博会议的重要性在于这是遗传学家和晶体学家的首次聚会。诚然,达林顿、贝尔纳和阿斯特伯里曾在1935年英国科学促进会的布莱克普尔会议上邂逅相逢,并讨论了染色体配对问题。但是,贝尔纳对有丝分裂和减数分裂的研究热情,却是在克兰本博会议之后才开始的。沃丁顿在回忆这些英国人从哈里奇到埃斯堡的颠簸旅途时,曾这样写道:
我们乘坐的是二等舱(那时候的旅费补贴很少),我们中的多数人设法在大厅的长凳上打盹,由于达林顿通宵达旦对贝尔纳讲了“每个人都需知道的遗传学和细胞学”,他俩也就把晕船的事儿忘得一干二净了。破晓前,贝尔纳已断定有丝分裂纺锤体一定是正半晶体,而染色体的分离可能是着丝粒产生某种流体的结果,这种流体在较大的正半晶体内形成负半晶体。
(沃丁顿,1939,318)
后面我们将讲到贝尔纳对在有丝分裂中起作用的力的看法。这里我们仅提一下,贝尔纳同穆勒一样(穆勒未参加克兰本博会议),在研究远程力方面没有取得结果,但他正确地论证了远程力存在于核蛋白胶体中(贝尔纳和范库亨,1942)。阿斯特伯里主要致力于推进核酸在基因复制中的作用的观点。他认为,既然蛋白质中的氨基酸都呈左螺旋,因此在基因复制时一定存在某种机制,以防止产生亲代分子的镜像。这可能是通过“外分子‘模板’”(阿斯特伯里,1939,50)或多核苷酸链的中间连接实现的。这个问题
……有两个方面。一个方面是嘌呤残基,另一方面是磷酸。其中之一会连接到原来的蛋白质链上,而另一个会使一种氨基酸构成一种新的链。核酸的插入,保证了新链具有与原来链相同的构型,但又不是原来的镜像。
(沃丁顿的报告,1938,1969,320)
一年之后,阿斯特伯里在爱丁堡会议上再次提出他的上述看法。他推测说:“核苷酸行列的功能,在于连接对映结构体间的层级。”(阿斯特伯里,1939,50)许多年以后,在思考蛋白质与核酸缔合的原因时,他写道:“我认为,部分原因是多肽链与多核苷酸行列在尺寸方面很一致。不管还有其他什么要求,至少分子之间一定是‘相配’的……”(阿斯特伯里,1948,21)
生理物理学家弗里德利希-弗雷克萨也得出了类似阿斯特伯里的基因复制观点。他认为,在组蛋白与核蛋白酸性磷酸基团之间存在静电引力。弗里德利希-弗雷克萨关于基因复制的概念,是同克兰本博会议上的讨论分不开的。因为正是贝尔纳的1938年在哥本哈根(当然在克兰本博会议上)的一篇讲演启发了物理学家帕斯卡尔·乔丹,而乔丹的论点又促使弗里德利希-弗雷克萨提出了上述概念。贝尔纳曾试图不用特殊引力而用远程非特殊力来解释染色体的配对。物理学家乔丹摒弃了贝尔纳的看法,他提出了谐振产生特殊引力的结论。乔丹认为,处于同一位置上的两个完全相同或几乎完全相同的分子,如果具有不同的能态,也就是这两个分子分别出于受激状态和基态,那么,这两个分子之间就会产生量子力学上稳定的相互作用。该作用足以说明染色体配对,也许还可说明染色体合成。他怀疑正常的价力不够大,这一点可从不太复杂的合成过程中看出;但乔丹没能解答:为了使分子中的一些部分提高到激发状态,分子的刚性是怎么被打破的。后来,他抛弃了染色体分子模型,引进了原子“拟液态凝聚”来说明这一问题。因此,他认为原子只是疏松地结合着,因而能经受位置变化(乔丹,1938,714)。两年后,莱纳斯·波林看到乔丹的这篇论文后,立刻批驳了乔丹的理论,并与德布吕克合写一篇文章发表在《自然》杂志上。他们确信,生物特异性问题的答案,只能从众所周知的共价键和氢键,范德瓦尔力和静电相互作用中去寻找(波林、德布吕克,1940)。弗里德和希-弗雷克萨则提出了在核蛋白的碱性组蛋白和酸性磷酸根之间存在静电引力的观点,并以此来取代乔丹的谐振引力论(弗里德利希-弗雷克萨,1940)。
当时柏林德皇威廉研究所的全体工作人员,只能使用协会里唯一的那个游泳池(梅尔彻斯,1970)。在游泳池里,弗里德利希-弗雷克萨遇到了乔丹,他们一起讨论了基因复制问题。《染色体结合时的有效力及其对核蛋白复制的意义》就是讨论的结果。人们可能会提出一个问题:弗里德利希-弗雷克萨是怎样认为现存基因的多肽链序列传递到新基因中去的。除了谈到酸性或碱性氨基酸的局部浓度外,弗里德利希并未考虑到这个问题。
基因的化学序列
值得一提的是,在1938年克兰本博会议上,来自哥本哈根卡尔斯堡实验室研究蛋白质的化学家艾伯特·费希尔是出席该会议的第一位化学家。物理学家与遗传学家之间的联系,在二十世纪三十年代研究X射线诱变时已建立起来,而同细胞学家的联系,则是通过研究染色体运动才建立的。卡斯帕森的细胞化学,在细胞学家和化学家之间建立联系起了牵线搭桥的作用。他们主要是共同研究了核组蛋白。但是,他们之间的联系是到三十年代末才形成。而且,即使在那时,核组蛋白的化学性质还未真正搞清,因此,大多数生物学家对此仍半信半疑。所以,这还不是把多肽链中基因的和氨基酸的线性序列联系起来的一个明显步骤。在三十年代,真正做到这一点的是英国培养的南美数学家多萝西·林奇。1934年,她在剑桥理论生物学俱乐部的一次非正式会议上提出这样的看法:基因的特异性,在于其氨基酸序列的特异性。1934年,《自然》杂志有一条简短评论提到了林奇在这次非正式会议上的发言。李约瑟写的这条评论谈到了林奇在俱乐部上的讲话,她用长度为3.5Å的T片段来描述基因的组件,因此长为100Å的基因将包含28个T片段。这些T片段的主干代表核酸的横向“纬线”,而T片段的顶部则代表多肽染色体线的纵向亚单位。这种蛋白质“经线”,包含了许多平行的和(对任何一个基因来说)相同的多肽链。1935年,林奇在曼彻斯特大学物理系作的一次讲演中,再次提出了上述结构图。一年后出版的《原生质》一书中,也记述了这结构图。1935年,林德斯特龙-兰格和拉斯马森已分析了鲱精蛋白的氨基酸成分,发现丙氨酸、缬氨酸、丝氨酸、羟基缬氨酸、脯氨酸和羟基脯氨酸中的每两个氨基酸就有4~6个精氨酸残基。这两位科学家以A代表精氨酸,M代表这6种单氨基酸,并根据各种氨基酸所占百分比排出了下列顺序:MAAMAAAAMAAMAAMAAA。据林奇计算,即使多肽的七分之五由精氨酸残基组成,余下的由这6种单氨基酸构成,那么在40µm米长的片段中可以有1050000种排列方式(林奇,1936,558)。
倘若林奇的染色体分子模型不遭到抨击,那么她提出的序列假说就更会受到人们的青睐。她假定环状四核苷酸同染色体轴线成直角,所以染色体应是正双折射而不应为负双折射。可是,施密特在吉森立即对林奇的模型提出异议,因为这同他观察到的DNA纤维和染色体的负双折射不相一致(施密特,1932)。有丝分裂过程中获得的正双折射证据(卡斯帕森,1940),不足以支持林奇模型,因为这是由于中期的超螺旋引起的。好在提出这种序列假说的不只是林奇一个人,至少还有另外四个人。他们是J.B.利斯、N.K.考佐夫、马克斯·伯克曼和卡尔·尼曼。利斯在1926年对英国科学促进会生理学组所作报告,考佐夫于1927年在列宁格勒一次会议上的发言,伯克曼和卡尔·尼曼1937年在《科学》杂志上发表的文章都持同样的观点。利斯和考佐夫只指出不同的氨基酸序列可以产生多样性。利斯宣称,很可能“分子中每种氨基酸的次序和所占比例是固定的,由此可解释蛋白质的形状和特性”(利斯,1926,388)。他还进一步作了计算:由50个连接的19种不同氨基酸组成的多肽链,可以有1048的排列方式。戈特纳那本颇有影响的教科书《生物化学大纲》(1929,358)和古利克发表在1937年《生物学评论季刊》上的重要论文,都摘引了利斯的报告。正是戈特纳促使卡麦蒂注意到利斯的观点。
考佐夫把氨基酸不同顺序的特殊性的讨论,同染色体结构联系起来。染色体结构序列是不变的。就像由同一块印刷版印出来的序列一样。所有新的染色体来自已有的染色体,细胞来自细胞,“我们现在增加一个新的命题:自然界中产生的每一个白蛋白分子来自已有的白蛋白分子,这是通过白蛋白分子周围溶液中的氨基酸和其他白蛋白碎片结晶生成的。分子来自分子”(考佐夫,1928,368)。
伯克曼和尼曼合写了一篇不同凡响的论文。他们试图论证千差万别的蛋白质在组成上有某些规律性。他们的氨基酸序列学说是以下述假设为根据:一个氨基酸对蛋白质分子的全部贡献在于按一定比例平均地分布在多肽链的全长上。斯维德伯格已有证据证明蛋白质是由大小相同(相对分子质量为17600)的亚单位所构成;伯克曼和尼曼指出,这些亚单元的序列受到一种总的约束,即链中任何片段的组成,必定是从整个分子中得到的氨基酸组成。他们论证说,如果对序列没有这种约束,那就不可能发现序列。知道了这种约束,就可从总的组成上获得关于序列的线索。例如,丝蛋白中的氨基酸50%是甘氨酸。因此人们认为,在链上每隔一个氨基酸残基就有一个甘氨酸。同样,每四个氨基酸有一个丙氨酸和每十六个氨基酸有一个酪氨酸(见表7.1)。科赛尔和达金关于鱼精蛋白的数据也差不多,后来林德斯特龙-兰格和拉斯马森在哥本哈根的研究工作也是如此,这些支持了林奇关于基因的氨基酸序列假说。
表7.1 根据氨基酸组成计算的氨基酸序列
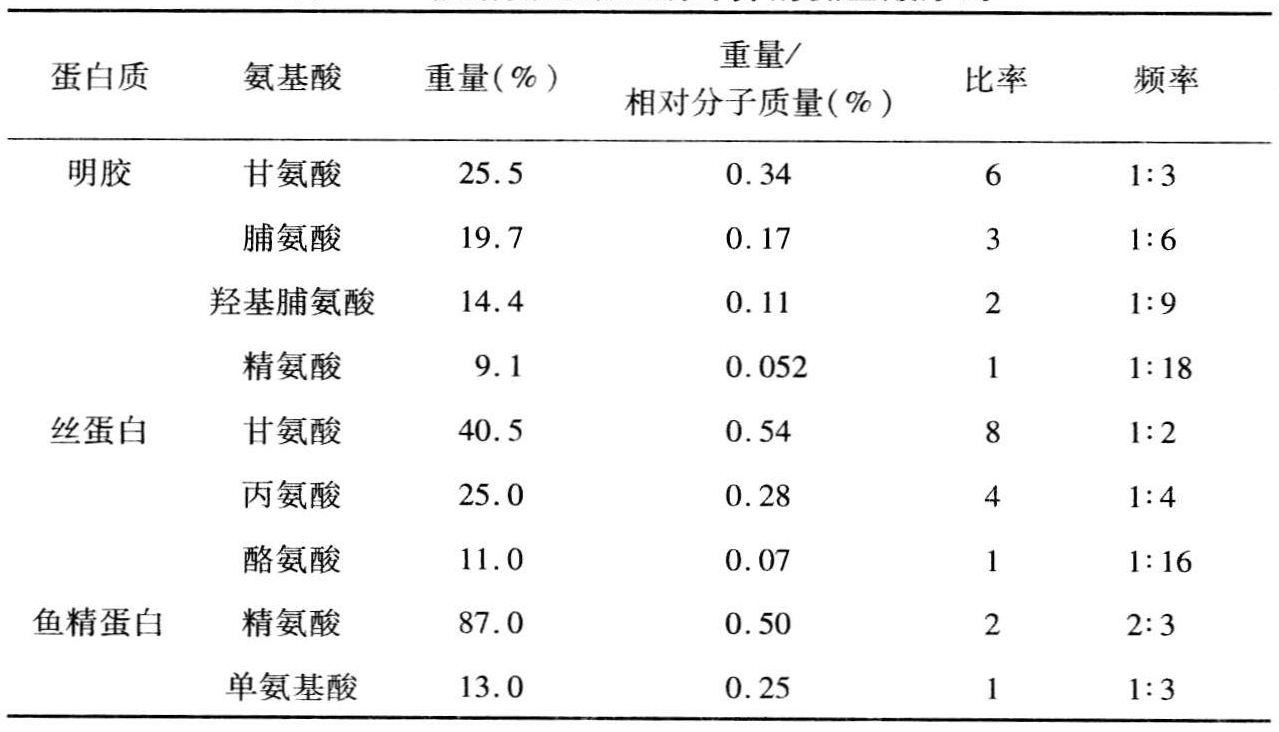
介绍了为得到一个言之成理的序列假说的这些早期努力以后,这些努力有什么用呢?其作用恐怕是微乎其微。但毫无疑问,他们所作的这些尝试至少起到了这样的作用,即把序列问题提了出来,而这个问题在埃米尔·费希尔之后是被人们所忽视的。对于早期从事分子结构研究的人们来说,伯克曼和尼曼的看法是可以接受的。
他们的观点明显地影响到弗洛伦斯·贝尔在1939年提出的命题。阿斯特伯里也认为一定存在着这些规律,从而开始了他的研究。另一方面,波林和尼曼在1937年时则对这个问题持审慎态度。
国际遗传学会议
1939年8月,爱丁堡第七届国际遗传学会议就基因问题召开了几次专题会议。在瓦维洛夫缺席情况下,由F.A.E.克鲁担任会议主席。会上宣读了未到会者的论文,并就基因和病毒的结构进行了讨论。用穆勒的话来说,讨论是“热烈和透彻的”(穆勒,1939,816)。由阿斯特伯里主持的一次讨论基因和染色体的会议上,达林顿说:“这是一次卓有成效的生物学和物理科学的聚会,是一次标志着一个新纪元开始的聚会。”(达林顿,1939,817)
开幕那天,来自55个国家的600位遗传学家参加了会议,会议洋溢着成功的热烈气氛。但是到第二天,“外界的议论越来越厉害,会议气氛受到了干扰”(克鲁,1939,496)。首先离开大会的是由34位代表组成的德国代表团,接着17人的荷兰代表团也离开了会场,因荷兰已发出了战争动员令。“从此以后,爱丁堡的公民们把这次大会看成是“国际形势”可靠的晴雨表。每小时我们都会收到关于意大利、法国和波兰代表团是否还在的询问……”(达林顿,1939,497)
风云突变也未能遏止国际上为解决基因复制这一重大问题的热情。直至1942年,柏林达仑的德皇威廉研究所和美国的一些生物学研究也许没有受到严重影响(梅尔彻斯,1971),瑞典为卡斯帕森提供了一幢新大楼,使他从1944年起一直继续进行研究(卡斯帕森,1944)。但是,英伦三岛的科学界却很快感到了第二次世界大战的影响。1942年12月,达林顿飞往瑞典途经英国时,阿斯特伯里请他转告卡斯帕森他遭到的困难:“工作甚多而助手极少,还要担任大学飞行中队的领航员。”(阿斯特伯里,1942)1942年初,阿斯特伯里在给美国范库亨的信中这样写道:“有数不清的工作要做,而我既无秘书,又无私人助手,甚至连实验室的杂务工也没有!该死的战争!”(阿斯特伯里,1942)
冷泉港会议
1941年在长岛北岸冷泉港的静谧环境里,再次讨论了上面提到的基因复制问题。哥本哈根会议的两位参加者——穆勒和德布吕克也出席了这次会议。
当时,穆勒问林奇是否仍坚持在爱丁堡会议上提出的染色体分子模型:在构成基因的多肽链四周的四核苷酸环被打开,从而进行基因和染色体的复制。她谨慎地回答说:
我想这一观点也许是值得考虑的。但在这段时间里,我一直在设想这样的可能性:蛋白质合成是发生在天然蛋白质的表面,也许像我现在提出的原生质那样的结构类型。
(林奇,1941,234)
她在阐明细胞质的这种组织结构时自然没有提到氨基酸序列。而且我也不知道自从1936年她详细说明染色体分子模型后,是否曾提到过序列问题。
在大战初期曾经最精确地阐明中心法则的蛋白质版本的这位科学家也缄默不语了,此时这方面的研究也暂告段落。就核酸来说,现在可看成是表明核酸的遗传特异性的证据,在当时却遭到截然相反的处理。穆勒在总结1941年冷泉港会议时,他从基因的蛋白质性质角度上冷漠地回顾了紫外线诱变问题,并以对德布吕克的贡献提出异议来结束他的发言。德布吕克的贡献是基因复制的氨基酸共振模型;在这个模型里,他再次采用了相似氨基酸之间存在着立体化学配合的观点。德布吕克的论点没有涉及特定的远程力,这比穆勒有所前进。但关于立体化学配合问题,穆勒的不满不是没有道理的,他说:“无论从普通化学或从几何学的角度来看,都不能证明为什么应该是这种情况……”(穆勒,1941,305)至于核酸,他俩只是对德布吕克的模型没有解释核酸的作用表示关切。
穆勒不禁又回到了他深信不疑的想法——存在着在染色体配对和基因复制中起作用的特殊远程力。他很想知道,“我们能否肯定负命题:即在基因之间,在距离大于一般原子之间的距离上,不可能存在特定的引力”(同上,306)。他回顾了帕斯卡尔·乔丹模型,并对该模型作了修改以对付1940年莱纳斯·波林和马克斯·德布吕克对该模型提出的批评。穆勒几乎没有认识到波林说服德布吕克作为合作者所写的那篇论文已预示了DNA复制的对称原理:
在推测自体催化的可能机制时,从结构化学家的观点来分析互补和同一可能同时发生的条件时,它看起来是最合理的。
(波林、德布吕克,1940,78)
八年后,波林陈述了这些条件可能是:
如果用作模板(基因或病毒分子)的结构是由互补结构的两部分所组成,则每一部分都可作为产生另一部分复制品的模型;两个互补部分的复合体也就可作为复制自身的模型。
(波林,1948,10)
波林并不是唯一发现在一个相同结构中出现互补与同一的人。如图7.5所示,阿斯特伯里已根据反向平行多肽链而设想出互补复制。
阿斯特伯里是在斯维德伯格的影响下提出这种设想的。斯维德伯格发现蛋白质分子量是一个基数的整倍数的模式。阿斯特伯里用一条基本多肽链连续复制产生更大分子量来解释这一点。他认为这个论点的核心是:
蛋白质链并列排列形成一定的图式,这种图式中的蛋白质链互不相干,他们之间的关系好比是物体同它自己的镜像。遗传理论的最新进展需要有一种可以复制出与每个“基因”相联系的特性的机制,人们现已承认,所提议的链系统可提供必要的基础。
(阿斯特伯里,1931,21)

图7.5 阿斯特伯里提出的基因复制的可能简图,(b)由互补多肽链组成(录自阿斯特伯里、伍兹,1931,664)。
我们可看出,1939年时,阿斯特伯里已改变看法,把核酸看作是亲代和新生多肽链的中间物。由于基因的核蛋白理论而忽视了直接互补复制,但DNA有了它自己的作用。
结语
试图用直接或间接的方法来确定遗传物质的化学特性全都失败了,这是不容讳言的事实。卡斯帕森的新细胞化学又一次求助于化学,但期待出现化学方法与细胞学方法融为一体的人们失望了。穆勒沉湎于研究染色体力学作为揭示基因复制的有力的手段,林奇则试图论述整条染色体的行为及其基因组成的结构。但是,他俩都失败了。正如前面所暗示的,有效地研究方法只能是从头做起,并遵循量子力学的原理进行工作,先解决简单的有机化合物的结构,然后再研究从细胞中提取出来的比较复杂的有机化合物的结构,最后才回到研究细胞和染色体。染色体并未为阐明基因复制机制提供线索,而物理化学和结构晶体学倒能给人以启迪。
【注释】
[1] 这是直线对称型,不是空间对称性。
[2] 佩因特当然不是唯一作出贡献的人,第一位把带纹同遗传图联系起来的是科斯托夫(参阅达林顿,1937,175ff)。
[3] 原文为Paris chromosomes,也许是指用巴黎绿染料染色的染色体——译者注。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









