



面包酵母的麦芽糖发酵力和葡萄糖阻遏作用对不加糖面团发酵力的影响
姜天笑 肖冬 光刘青
(天津科技大学天津市工业微生物重点实验室 天津 300222)
摘要:对8株供试菌株的麦芽糖发酵力、抗葡萄糖阻遏能力和不加糖面团发酵力三者之间的关系得出:高麦芽糖发酵力且抗葡萄糖阻遏作用的菌株(BY-14)其不加糖面团发酵力较高,其中面包酵母的麦芽糖发酵力与不加糖面团发酵力存在一定的相关性(r=0.917) 。对菌株BY-14进行紫外-氯化锂复合诱变,得到突变株BM-72和BM-11,其不加糖面团发酵力均有不同程度的提高。通过比较得出:菌株的麦芽糖发酵力和葡萄糖阻遏作用对不加糖面团发酵力都产生影响,其中葡萄糖阻遏作用可能对面团发酵力影响较为显著。
关键词:面包酵母;发酵力;阻遏作用
面包酵母是面包制作生产工程中不可缺少的四大要素原料之一,是重要的微生物发酵剂和生物疏松剂。在面粉中含有少量的葡萄糖、果糖、麦芽糖等可发酵糖类,其中大部分为淀粉[1],淀粉在面粉中液化酶和糖化酶的作用下破裂转化为麦芽糖,因此麦芽糖是存在于不加糖面团中最为丰富的可发酵糖类,麦芽糖的利用能力是决定面包酵母发酵性能的关键因素。在面包酵母中有5个不相连的MAL基因位点(MAL1-4和MAL6) ,编码麦芽糖透性酶和α-D-葡萄糖苷酶(麦芽糖酶) ,用于将麦芽糖运输至胞内并水解为葡萄糖可供细胞利用[2]。在不存在葡萄糖时,少数几个麦芽糖分子通过一种非专门的运输机制进入细胞,然后诱导麦芽糖酶和麦芽糖透性酶的合成,从而进一步使麦芽糖快速进入细胞被水解为两个葡萄糖分子。葡萄糖阻遏麦芽糖的利用机制是复杂的,一般,葡萄糖在麦芽糖利用过程中起到两个作用:一是它的存在干涉了由麦芽糖诱导的MAL基因的转录从而抑制麦芽糖利用酶系的表达;二是葡萄糖诱导麦芽糖透性酶的失活[3,4]。酶产生出现的迟滞期将导致麦芽糖发酵速度的降低,面包酵母若能快速从代谢葡萄糖和果糖转入利用麦芽糖,甚至在葡萄糖存在时仍可利用麦芽糖,也就提高了面包酵母的发酵速度,从而提高了不加糖面团的发酵力[5]。
在本文中,我们通过复合诱变筛选出在不加糖面团中具有高发酵性能的突变株BM-72,讨论了麦芽糖发酵力与葡萄糖阻遏作用对面包酵母不加糖面团发酵力的影响。其中,葡萄糖阻遏作用可能对不加糖面团发酵力影响较为显著。
1 材料与方法
1.1 实验材料
1.1.1 菌株
实验室保存的面包酵母菌株,编号为BY-6、BY-11、BY-12、BY-14、BY-15、BY-16、A6、B6。
1.1.2 培养基
YEPD培养基: 2%葡萄糖,1%酵母粉,2%蛋白胨。
糖蜜培养基:将处理后糖蜜(30~35Brix) ,加水稀释为12Brix,添加酵母粉0.5%,硫酸铵0.05%,pH5.0。
模拟面团培养基[6]:麦芽糖4%,硫酸铵0.25%,尿素0.5%,磷酸二氢钾1.6%,磷酸氢二钠0.5%,硫酸镁0.06%,烟酸22.5ppm,泛酸5.0ppm,维生素B12.5ppm,维生素B6 1.25ppm,维生素B21.0ppm,叶酸0.5ppm。 (若含有葡萄糖,其加入量为0.5%) 。
YEPM-LiCL平板: 2%麦芽糖,1%酵母粉,2%蛋白胨,0.3%LiCL,2%琼脂。
D-氨基葡萄糖平板[7]:麦芽糖2%,磷酸二氢钾0.1%,硫酸镁0.05%,硫酸铵0.05%,D-氨基葡萄糖0.6%,处理琼脂粉2%,pH6.0。
1.2 实验测定方法
1.2.1 葡萄糖含量测定
采用生物传感仪测定,单位为mg/100ml。
1.2.2 麦芽糖含量的测定
由于麦芽糖和葡萄糖都为还原糖,则培养基中麦芽糖的测定为菲林热滴定法[8],若培养基中同时含有葡萄糖,麦芽糖含量(%)=总糖(%) -葡萄糖含量(%) 。
1.2.3 不加糖面团发酵力的测定[3]
面粉4.0g,鲜酵母泥0.2g (干物质含量约为16%) ,30℃水7ml,加入20ml的刻度试管中,用玻璃棒迅速混匀成糊状,30℃静置培养45min,记录面团上升高度,单位为ml/(0.01g酵母干重· 45min) 。
1.2.4 发酵初筛实验
从斜面上挑取一环菌体于1.5ml无菌水中,30℃饥饿培养4~6h,吸取0.5ml菌悬液于9.5ml的模拟面团培养基中,静置培养12h,在660nm下,测定吸光度A0和A12,A0和A12分别为培养基初始和培养12h后的吸光度值。 (A12-A0)之值大于出发菌株10%初步认定为正突变株。
2 实验结果
2.1 酵母菌株的麦芽糖发酵力
酵母细胞培养至生长稳定期后接入10ml液体模拟面团中(以麦芽糖作为唯一碳源) ,接入量为5%,30℃培养一定时间,测定培养基中的麦芽糖含量,结果见表1。由表1看出,菌株BY-11和BY-16的麦芽糖发酵力较低,BY-6、A6、BY-12、BY-15和B6麦芽糖发酵力相当; BY-14最高。
表1 各菌株麦芽糖发酵力u (g耗糖· h-1· g-1酵母干物质)
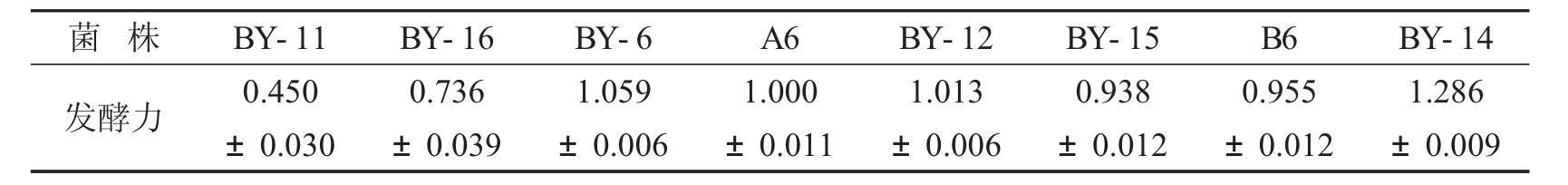
注:麦芽糖发酵力指30℃下培养液中单位酵母量(g干物质)每小时的麦芽糖消耗量(g) 。每个实验重复两次取平均值作为实验结果。
2.2 葡萄糖对面包酵母麦芽糖发酵速度的影响
以菌株BY-6和BY-14为例,以0.5%的接种量接入10ml模拟面团中(同时含有葡萄糖和麦芽糖) ,每30min测定模拟面团中麦芽糖和葡萄糖含量,绘制耗糖曲线,见图1。
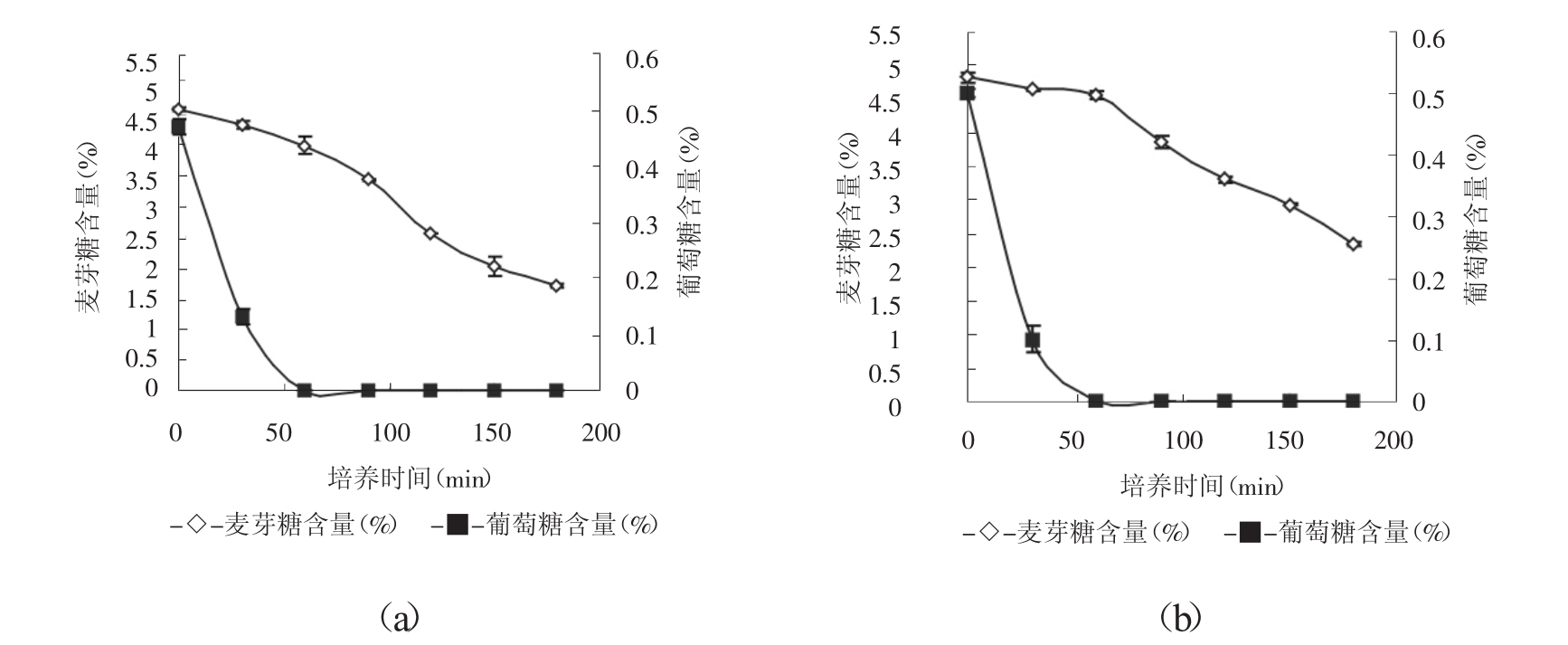
图1 麦芽糖和葡萄糖发酵速度曲线(a:菌株BY-14,b:菌株BY-6)
由图1可知,菌株BY-6在培养前60min,麦芽糖消耗量基本为0,随着葡萄糖消耗殆尽后,麦芽糖消耗才开始呈明显的下降趋势,说明菌株BY-6受葡萄糖阻遏作用明显;而菌株BY-14在有、无葡萄糖存在下,麦芽糖消耗曲线一直呈下降趋势,即在葡萄糖存在下,菌株BY-14仍可利用麦芽糖,但此时麦芽糖的消耗速度较慢,当葡萄糖含量为0后,麦芽糖的消耗速度相对较快,说明菌株BY-14受葡萄糖阻遏程度较小。各菌株受葡萄糖阻遏程度见表2。
表2 各供试菌株受葡萄糖阻遏作用程度比较

注:“-”表示菌株的麦芽糖消耗量过低,未进行测定。
各菌株受葡萄糖阻遏程度I如下计算:
![]()
2.3 面包酵母菌株不加糖面团发酵力
供试8株酵母菌株不加糖面团发酵力比较,见图2。
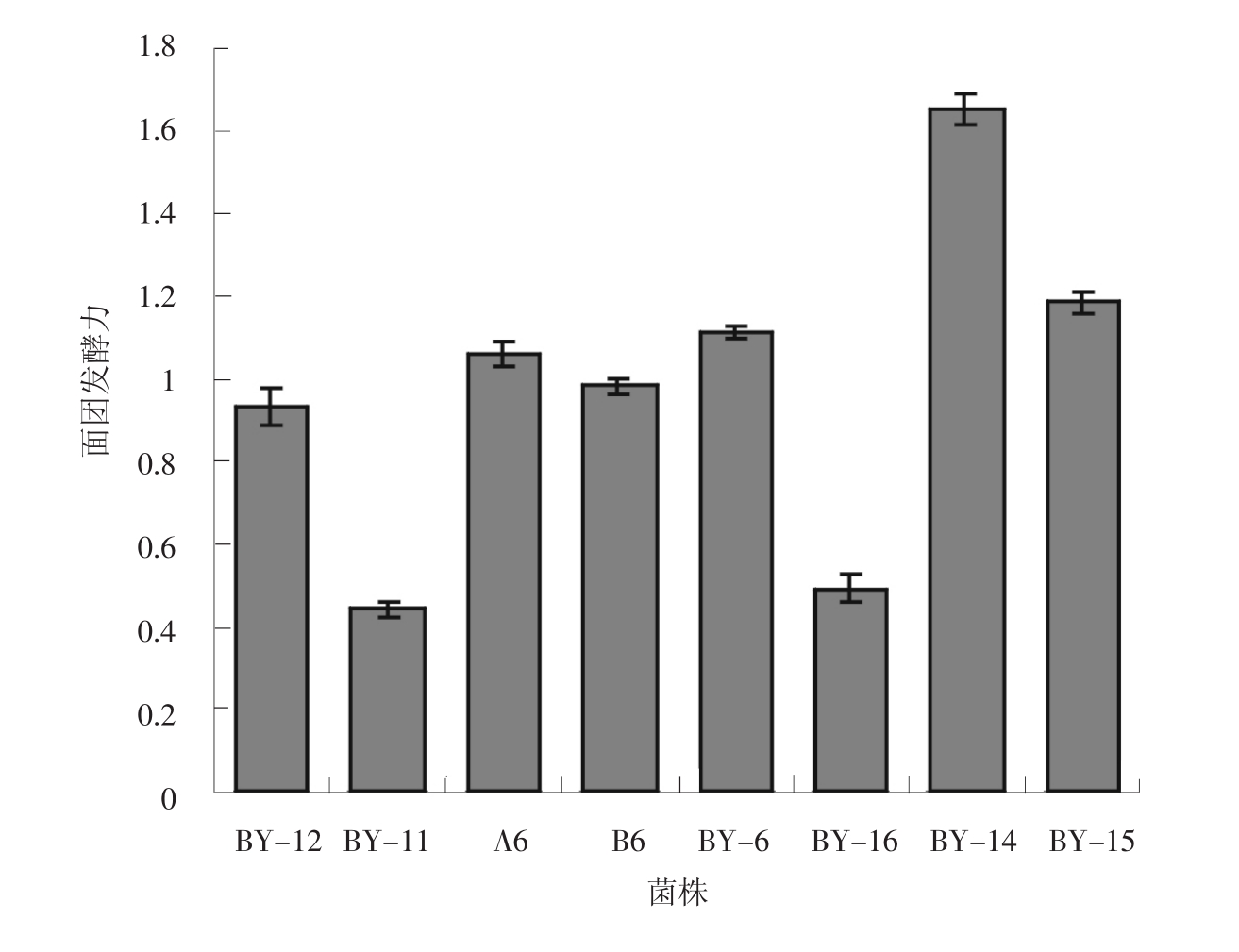
图2 各酵母菌株不加糖面团发酵力ml/ (0.01g酵母干重· 45min)
由图2可知,菌株BY-14较其他几株菌株表现出良好的不加糖面团发酵力,达到1.65ml/ (0.01g酵母干重· 45min) 。菌株BY-12、A6、B6、BY-6和BY-15不加糖面团发酵力相差不大,而BY-11、BY-16不加糖面团发酵力较低,基本在45min内不起发。
对供试8株菌株分析得出:高麦芽糖发酵力、具有葡萄糖阻遏抗性的菌株,在不加糖面团中的发酵力较高。
2.4 出发菌株的诱变处理和突变株的筛选
以菌株BY-14作为出发菌株进行紫外-氯化锂诱变,紫外线照射剂量为60s,而后涂布于YEPM-LiCL平板上,以D-氨基葡萄糖平板为初筛平板。 D-氨基葡萄糖为葡萄糖的结构类似物,可被酵母吸收、磷酸化,但不会被进一步代谢,它可产生与葡萄糖相似的阻遏作用[7]。在D-氨基葡萄糖平板上筛选出抗性突变株38株,再经过发酵初筛、复筛得到突变株BM-72和BM-11,在不加糖面团发酵力上较出发菌株BY-14均有不同程度的提高。将两株突变株在麦芽糖发酵力、抗葡萄糖阻遏程度和面团发酵力几个方面与出发菌株进行比较,结果见图3、表3。
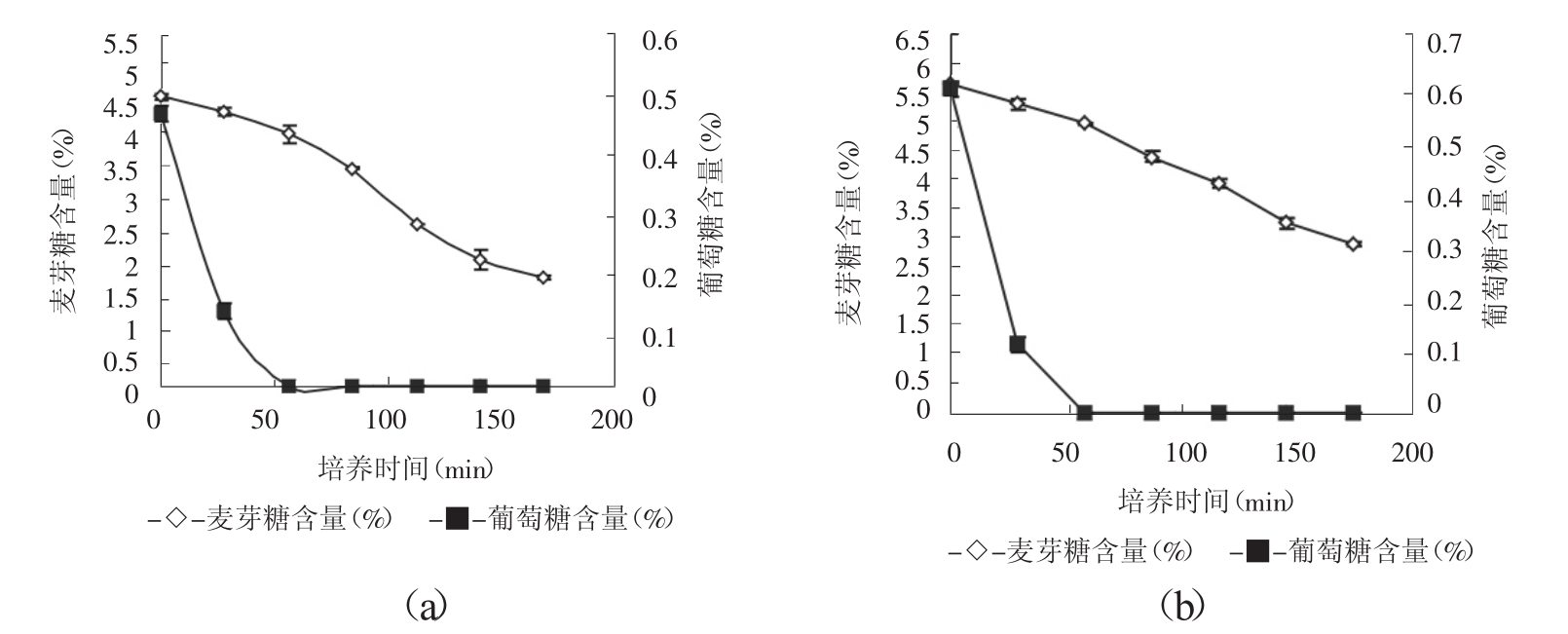
图3 麦芽糖和葡萄糖发酵速度曲线(a:菌株BM-11,b:菌株BM-72)
由图3可知,突变株BM-72和BM-11在葡萄糖存在下(培养前60min) ,与出发菌株BY-14相比,具有较高的麦芽糖发酵速度,且在有、无葡萄糖存在下,两突变株的麦芽糖发酵速度基本相同。
表3 BY-14与突变株BM-72、BM-11发酵性能比较

由表3可知,与出发菌株BY-14相比,突变株BM-72具有较高的麦芽糖发酵力,而突变株BM-11则较低;突变株BM-72和BM-11麦芽糖发酵速度曲线相近(图3-b)且两突变株受葡萄糖阻遏程度都较小,为抗葡萄糖阻遏菌株,即葡萄糖的存在对麦芽糖的利用基本没有影响。
3 讨论
面包酵母(Saccharomyces cerevisiae)是重要的工业微生物,在食品方面占有重要的地位和作用,一些无糖或低糖面包、苏打饼干等面食的制作主要为不加糖面团发酵。在不加糖面团发酵过程中,所需的面包酵母应具有较高的麦芽糖发酵速度以缩短发酵周期,降低生产制作成本。麦芽糖发酵力是在无葡萄糖存在下,菌株自身发酵麦芽糖能力的体现,较低的麦芽糖利用能力必然会对不加糖面团的起发速度产生不利影响;而葡萄糖的存在导致麦芽糖利用酶表达的延迟,势必会降低麦芽糖的发酵速度从而影响不加糖面团发酵力。所以两者都会对不加糖面团发酵力产生不同程度的影响。
由供试8株酵母菌株分析得出,麦芽糖发酵力与不加糖面团发酵力的大小存在一定的相关性(r=0.917) ;高麦芽糖发酵力和低葡萄糖阻遏程度的菌株(BY-14)在不加糖面团中的发酵力最高;通过紫外-氯化锂复合诱变筛选出的突变株BM-72和BM-11的不加糖面团发酵力均优于出发菌株BY-14。其中菌株BM-72的麦芽糖发酵力较出发菌株提高了15.2%,受葡萄糖阻遏程度降低了66%,不加糖面团发酵力提高了19.4%,这也说明了高麦芽糖发酵力且抗葡萄糖阻遏的菌株在不加糖面团中的起发速度较快。麦芽糖发酵力较低的抗葡萄糖阻遏突变株(BM-11)在不加糖面团中发酵力较高,原因可能是葡萄糖阻遏作用对面包酵母不加糖面团发酵力影响较为显著。
参考文献:
[1] BRIAN J.B Wood主编.徐岩译.发酵食品微生物学(第2版) [M].北京:中国轻工业出版社,2001
[2] Oda Y and Ouchi K.Role of the yeast maltose fermentation genes in CO2 production rate from sponge dough[J].Food Microbiology,1990,7 (1):43~47
[3] ANA M.RINCóN,ANTONIO C.CODóN,FRANCISCO CASTREJóN,et al.Improved Properties of Baker's Yeast Mutants Resistant to 2-Deoxy-D-Glucose [J].Applied and Environmental Microbiology,2001,67(9) :4279~4285
[4] HUA JIANG,IGOR MEDINTZ,BIN ZHANG,CORINNE A.MICHELS.Metabolic signals trigger glucoseinduced inactivation of maltose permease in Saccharomyces[J].Journal of Bacteriology,2000,182 (3):647~654
[5] BW Hazell and PV Attfield.Enhancement of maltose utilization by Saccharomyces cerevisiae in medium containing fermentable hexoses[J].Journal of Industral Micerbiology and Biotechnology,1999,22: 627~632
[6] Teunissen,A.,et al.Isolation and characterization of a freeze-tolerant diploid derivative of an industrial baker's yeast strain and its use in frozen doughs [J].Applied and Environmental Microbiology,2002,68 (10) :4780~4787
[7] CORINNE A.MICHELS and ANDREW ROMANOWSKI.Pleiotropic glucose repression-resistant mutation in Saccharomyces carlesbergensis[J].Journal of Bacteriology,1980,143 (2):674~679
[8]天津轻工业学院编著.工业发酵分析[M].北京:中国轻工业出版社,1980
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









