



云南元谋干热河谷地区6种植物种子的抗逆生理特性
崔 凯,李 昆,孙永玉,杨文云,刘方炎,李 立
摘 要:以云南元谋干热河谷地区的苏门答腊金合欢、新银合欢、木豆、车桑子、余甘子和山合欢6种植物种子为试材,在4月份干热气候的地表环境下,测定各类种子的生活力、呼吸代谢、渗透调节物质、膜质过氧化和膜保护酶系统活性。结果表明,随着处理时间的延长,各类种子的生活力显著下降;相对电导率、脯氨酸(Pro)和丙二醛(MDA)含量呈现持续增加趋势;呼吸强度,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性,可溶性糖含量呈单峰曲线变化;种子抵御逆境的能力是其呼吸代谢、膜保护酶系统、保护性物质协同作用的结果;影响各类种子抗逆能力的关键物质并不一致。
关键词:干热河谷;种子抗逆生理;膜质过氧化;膜保护酶系统
干热河谷是我国生态环境最为脆弱的地区之一,由于特殊的地理位置和独特的地形地貌,气候十分恶劣。元谋盆地是金沙江干热河谷的典型代表,年平均降雨量613.7mm,90%以上的降雨量集中在雨季(5~10月份),年均蒸发量3 426.3mm;年均相对湿度54%,无霜期350~365 d[1],旱季地面温度可达60~70℃。种子是高等植物特有的繁殖器官,种子期是植物重要的物候阶段,种子成熟掉落后如何在干热河谷高温干旱的地面环境下保持正常的发芽能力,是植物繁衍后代、实现植被持续恢复的先决条件。目前关于干热河谷植物种子抗逆性的研究鲜有报道,本研究以元谋干热河谷植被恢复中6种优良树种为研究对象,着重研究其种子在4月份干热气候的地表环境下其内部一系列生理生化变化及其与种子活力的关系,旨在为种子储存、育苗造林和天然更新,乃至维系该地区生态平衡提供基础数据。
1 试验材料
表1 1996~2005年元谋干热河谷4月份每日各气象指标的平均值

4月份是元谋干热河谷地区雨季来临前的一个高温干旱时期,所研究的6类种子在4月份都已经成熟并掉落于地面上。本研究探讨的是种子在此期间的抗逆生理过程。试验选用苏门答腊金合欢〔Acacia.glauca(L.)Moench〕、新银合欢〔Leucaena 1eucocephala(Lam.)Dewit〕、木豆〔Cajanus cajan(L.)M illspaugh〕3个引进树种,以及车桑子〔Dodonaea viscosa(L.)Jacq.〕、余甘子(Phyllanthus emblica L.)、山合欢〔Albizia kalkora(Roxb.)Prain〕3个乡土树种。供试种子于当年采自云南省元谋县干热河谷,采集时选择性状优良的母树上的成熟饱满种子,试验前统一在室内进行为期1个月的晾晒处理。试验用气象数据从元谋县气象局获取,包括元谋干热河谷地区1996~2005年历年4月1~30日每天02:00、08:00、14:00、20:00的地面温度、空气湿度、日照时数3类气象指标,各项指标取历年的平均值,用RX-智能型人工气候箱(宁波江南仪器厂)来模拟,模拟期为30 d。模拟期每天的平均气象指标值见表1。
2 研究方法
2.1 种子处理
选取饱满、大小较为一致的各类植物种子,1%的NaCl消毒15 min,消毒后的种子均匀摆放于垫有2层湿润滤纸的培养皿内,每个培养皿内放100粒,每类种子8个重复,然后置于人工气候箱内培养。
2.2 测定项目
每6 d为1个周期,用TTC法[2]检验种子生活力的保持情况,同时测定种子呼吸强度、相对电导率、丙二醛(MDA)含量、脯氨酸(Pro)含量、可溶性糖含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性8项生理生化指标。其中相对电导率测定参照宋松泉等[2]的方法;MDA含量的测定采用硫代巴比妥酸(TBA)法[3];Pro含量测定采用茚三酮比色法[3]; SOD活性的测定采用氮蓝四唑(NBT)光还原比色法[3],以抑制NBT光化还原的50%为1个酶活性单位;POD活性的测定采用愈创木酚法[3],以每分钟内A470nm变化0.01为1个酶活单位;CAT活性的测定采用高锰酸钾滴定法[3],酶活性以每克鲜质量样品1min内分解H2O2的毫克数表示;呼吸强度测定采用静置碱液吸收法[4];可溶性糖含量的测定采用蒽酮比色法[5],各项指标在测定时做3次重复,取平均值。
2.3 数据处理
试验数据用SPSS13.0进行分析,分析前数据经必要的标准化处理(其中结果为百分数的数据经平方根反正旋变换)。
3 结果与分析
3.1 处理期间种子生活力的变化
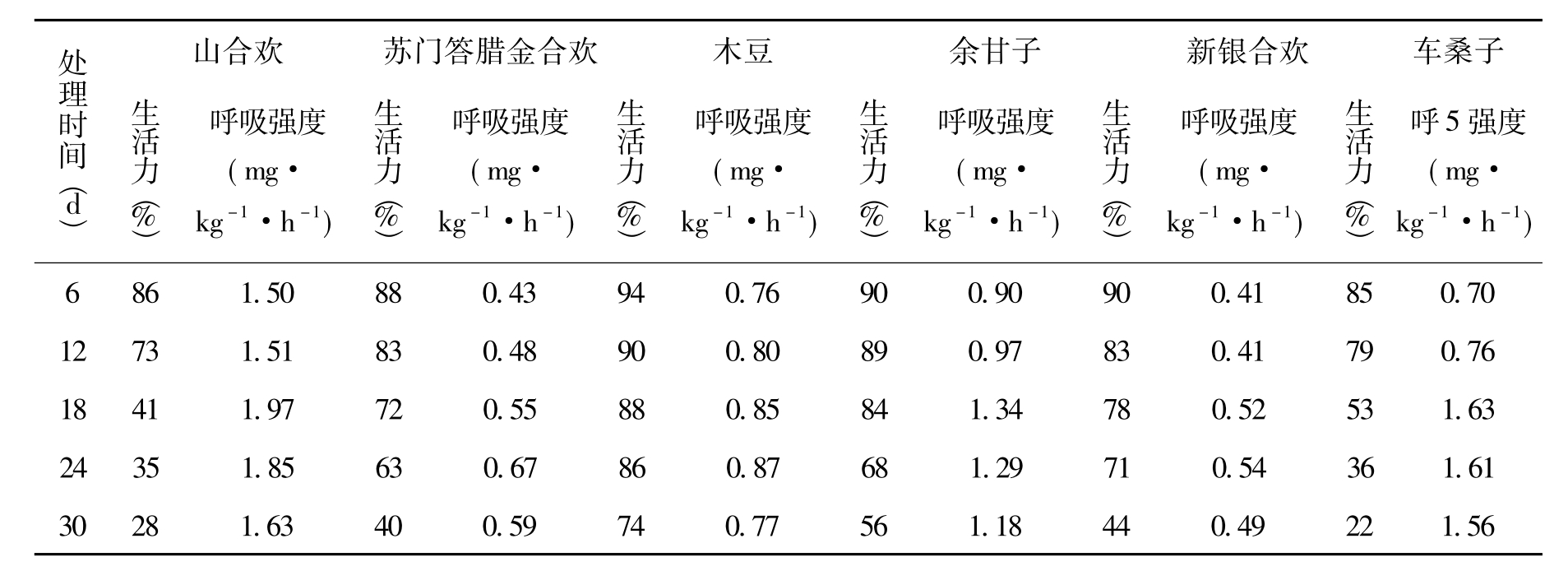
如表2所示,各类种子经过处理后,其生活力都显著下降,整个处理期种子活力变化阶段可大致分为缓慢下降期和快速下降期。其中木豆在处理24 d之前属缓慢下降期,之后进入快速下降期;余甘子处理到18 d时,从缓慢下降期进入快速下降期;新银合欢、苏门答腊金合欢、车桑子3类种子缓慢下降期和快速下降期的分水岭分别是24a、24a、12d;较为特殊的是山合欢,它处理12 d时从缓慢下降期进入快速下降期,到18d时又进入缓慢下降期。按处理期结束时各类种子活力大小排序为木豆74%、余甘子56%、新银合欢44%、苏门答腊金合欢40%、山合欢28%、车桑子22%。单因素方差分析表明,各类种子活力保持水平有显著差异(P<0.05)。
3.2 处理期间种子呼吸强度的变化
呼吸强度是植物生理代谢的重要指标之一。由于呼吸作用消耗了种子内部大量的碳水化合物,过高的呼吸强度通常被认为不利于种子生活力的保持。试验表明(见表2),6种植物种子在处理期间的呼吸强度比较微弱,基本都在2.5 mg·kg-1·h-1以下。其中,山合欢种子呼吸强度最大,其次是车桑子、余甘子、木豆、苏门答腊金合欢,新银合欢最低。随着处理时间的延长,各类种子的呼吸强度大体都经历了一个由缓慢增长到达高峰期,再缓慢降低的过程。但每个树种到达高峰期的时间并不一致,山合欢、木豆和余甘子在处理的第18天呼吸强度出现最大值,其余3种稍晚并且峰值不明显。另外,前3种植物在处理12~18 d之间的呼吸强度剧增,后3种植物则比较平缓。
表2 处理期间种子生活力和呼吸强度的变化
3.3 处理期间种子相对电导率和MDA的变化
植物在受到逆境胁迫时,由于细胞质膜损伤引起透性增大,电解质大量外渗,因此细胞质膜透性常作为植物抗逆指标之一[6-9]。处理后的测定结果表明(见表3),随着处理时间的延长,各类种子相对电导率都呈上升趋势,但其上升幅度有所差异。余甘子升幅最高,其次为车桑子和新银合欢,苏门答腊金合欢、木豆、山合欢等树种的相对电导率增幅较低,均不足8.5%。余甘子相对电导率是山合欢的8.5倍。单因素方差分析表明,除木豆和山合欢外,其他种子的相对电导率差异均达到显著水平(P<0.05)。
表3 处理期间种子相对电导率和MDA的变化
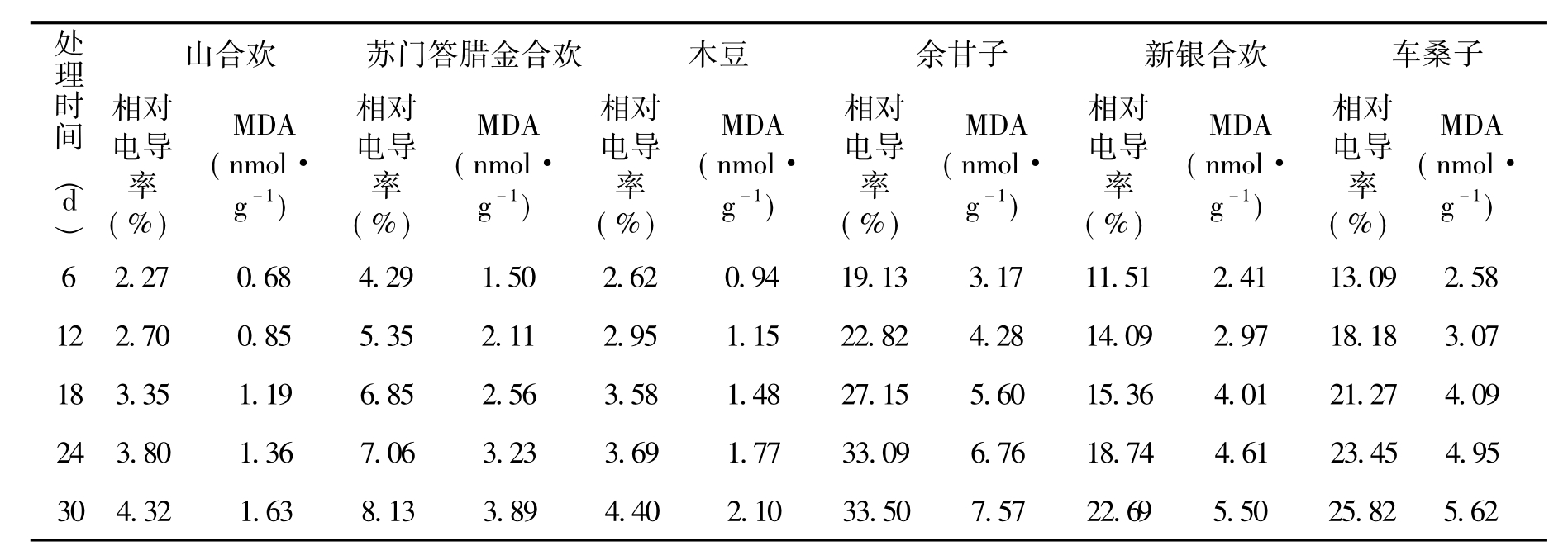
在逆境中的植物,细胞原生质膜中的不饱和脂肪酸会发生过氧化作用产生MDA,MDA进一步可与蛋白质、核酸、氨基酸等活性物质交联,形成不可溶的化合物,从而干扰细胞正常生命活动,使质膜系统受到伤害。MDA含量在一定程度上反映了植物体内自由基的含量,即MDA越少,自由基含量越低[10-12]。试验处理初期,各类种子的MDA总体数量很低(8 nmol·g-1以下),处理30 d后,余甘子、车桑子、新银合欢、苏门答腊金合欢、木豆、山合欢种子MDA的质量摩尔浓度分别比处理前提高了153%、190%、180%、202%、173%和194%。各类种子MDA含量变化与相对电导率具有显著的正相关性(P<0.05),相关系数达到0.91。
3.4 处理期间种子Pro含量和可溶性糖含量的变化
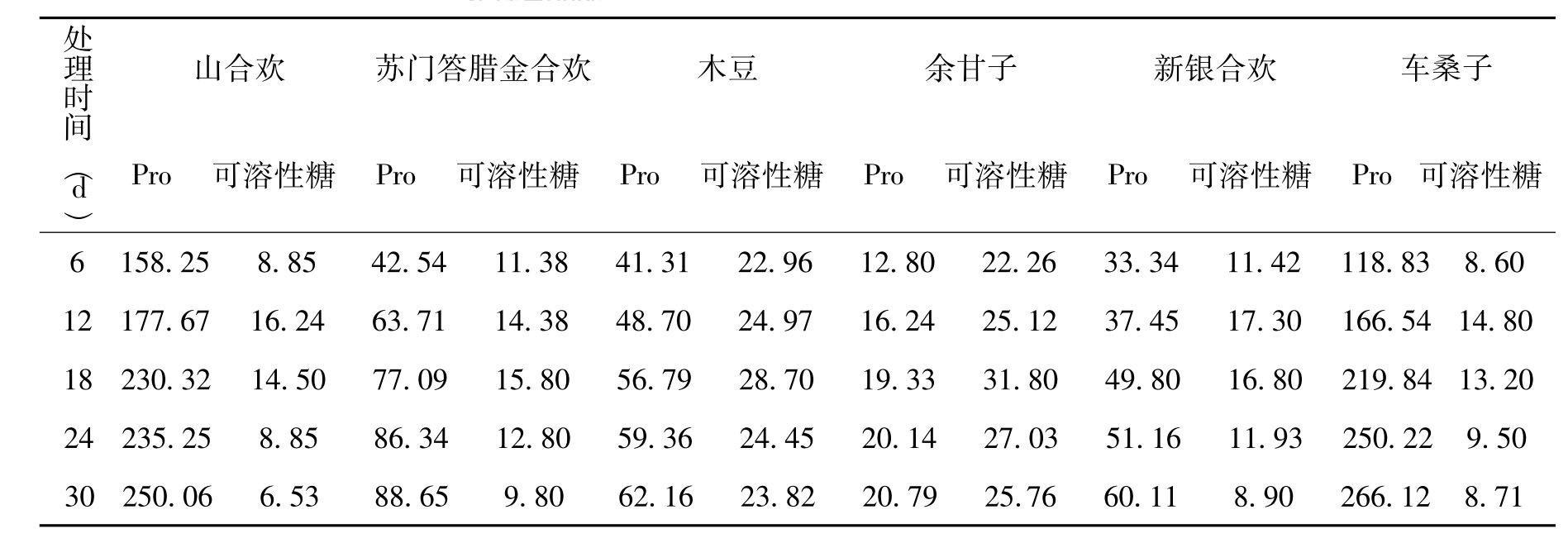
干旱胁迫下植物体内积累脯氨酸(Pro)具有一定普遍性,Pro可作为渗透调节物质及自由基清除剂而起保护作用从而增强植物的抗旱性[13]。Pro通常可以作为一种渗透调节剂或细胞质酶的保护性物质[14],保持细胞中生物聚合物的结构及膜的完整性,亦可能作为氮源或碳源的贮藏物质。同时,也有一些研究认为,逆境下Pro含量提高是一种受害症状[15]。各类种子在处理前内部并没有Pro,处理后迅速积累,且随着处理时间延长,积累量也不断增加。其中,山合欢和车桑子在整个处理期间Pro的积累量显著高于其他4个树种,在处理6d时,山合欢和车桑子的Pro质量分数分别达到1.58×10-4和1.19×10-4,是同期其他树种的4~5倍,另外4个树种种子中的Pro的积累较为平缓(见表4)。
可溶性糖是干旱胁迫诱导的小分子溶质之一,主要包括葡萄糖、海藻糖、蔗糖等。这些可溶性糖类参与渗透调节和复水后的生理恢复及修复过程,并可能在维持植物蛋白质稳定方面起到重要作用[16-17]。经过高温干旱处理后,各类种子可溶性糖呈现出先上升后下降的变化趋势,山合欢和木豆种子的可溶性糖含量显著高于另外4种植物;山合欢、木豆和苏门答腊金合欢种子的可溶性糖含量在18 d时达到最大值,其他3种植物则在12d时达到峰值。可溶性糖含量按大小排序依次是山合欢、木豆、新银合欢、苏门答腊金合欢、车桑子、余甘子(见表4)。说明含羞草科(M imosaceae)的豆类种子在高温干旱环境胁迫下,更容易积累可溶性糖,如山合欢和木豆种子,而含油量较高的车桑子和余甘子种子则相反。
表4 处理期间种子Pro含量和可溶性糖含量的变化(×10-6)
3.5 处理期间种子SOD、POD、CAT活性的变化
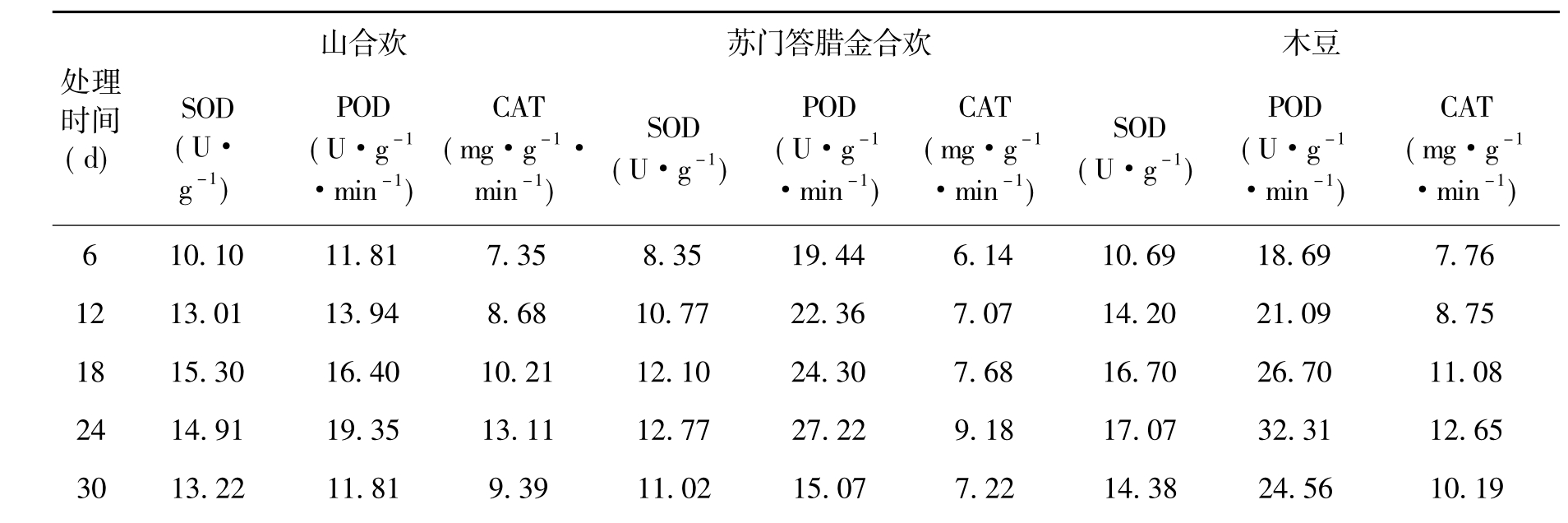
表5 处理期间种子3种保护酶活性的变化
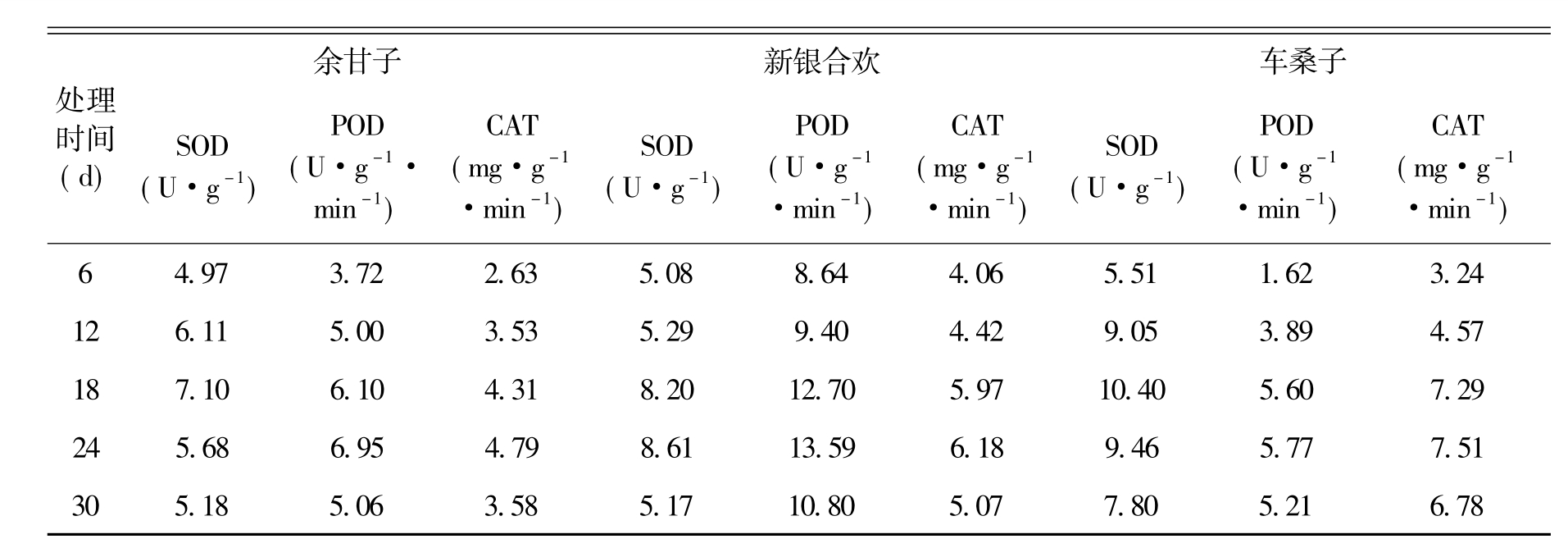
SOD是植物重要的活性氧清除酶,具有专一清除O2ˉ的功能,当外来胁迫导致大量活性氧产生时,能及时有效地将O2ˉ歧化为H2O2,保护细胞免受活性氧胁迫的伤害,在自由基清除系统中占有重要地位[18-19]。POD是植物体内重要的抗氧化酶,能有效的清除H2O2[20]。CAT是植物体内H2O2的清除酶,主要定位于过氧化物体中,分解H2O2为H2O和O2.[11]。
SOD活性的测定结果(见表5)表明,各类种子在整个处理期间的SOD活性变化差异较大,但都呈单峰曲线变化。总体上看,随着处理时间延长,各类种子SOD活性逐渐加强,然后迅速降低。余甘子、苏门答腊金合欢和山合欢在处理18 d时达到峰值,其余3种植物则在处理24 d时达到峰值,在处理期间各树种SOD活性大小分别是木豆、余甘子、新银合欢、苏门答腊金合欢、车桑子、山合欢。各树种Pro含量和SOD活性之间,基本呈现出相反的变化趋势,从这一结果看Pro的积累更像是种子受害后的症状和反应。
从各类种子POD和CAT活性测定结果(见表5)看,2种酶活性的变化趋势与SOD非常接近,也呈现出逐渐加强到迅速降低的基本变化形式。
4 结论
本研究中6种植物种子生活力的高低与3种保护酶活性高低显示了很强的一致性,即保护酶活性高的其生活力亦高。其中较为例外的是车桑子与山合欢,车桑子3种保护酶活性高于山合欢,而生活力低于山合欢,这可能归因于山合欢的Pro和可溶性糖含量都高于车桑子。新银合欢、苏门答腊金合欢、木豆、车桑子4类种子MDA含量与种子活力基本呈负相关,但MDA含量与种子活力在量化关系上并非严格对应,如余甘子在6种植物中MDA含量最高,而种子生活力却不低;山合欢MDA含量较低,但生活力并不高。这可能归因于山合欢和余甘子相比,呼吸强度相对较高,从而影响了种子生活力。可见,各类种子的抗逆性是受到抗氧化酶、保护性物质和呼吸代谢协同控制的,本研究中保护酶的活性基本上能反映各种子的抗逆性,6类种子抗逆性的强弱依次为木豆、余甘子、新银合欢、苏门答腊金合欢、山合欢、车桑子。
根据当地气象站资料,元谋干热河谷4月份近10 a月平均地面温度为29.47℃,14:00地面温度更是高达50℃以上,月平均最高地面温度为57.83℃;空气湿度月均只有47.3℃。本研究表明,6种植物种子处在此干旱高温条件下时,总体上是通过增加可溶性糖含量、增加Pro含量、增强膜保护酶活性并使呼吸强度维持在一个非常低的水平来保持自身活力的。这与前人报道的一些植物抗逆生理机制大体相似[21-25],如许冬梅等[22]对赖草〔Leymus secalinus(Georgi)Tzvel.〕和中亚白草(Pennisetum centrasiaticum Tzvel.)的研究表明,不同生境的2类草是通过积累Pro和可溶性糖含量,同时增强抗氧化保护酶活性来抵御逆境的;路兴花等[21]也指出,不同生态稻区的水稻在逆境胁迫下,Pro、MDA和可溶性糖含量有增加的趋势。各类种子抗逆性很难用单一的指标来解释,原因在于种子处理期间,其呼吸代谢、糖代谢、细胞膜系统、抗氧化酶系统等生理生化指标都相应发生了变化。每种植物在抵御逆境中起关键作用的物质并不一致,比如山合欢的Pro、可溶性糖含量在6种植物中都是最高,而其3种保护酶活性在6种植物是最低的,它的相对电导率和MDA含量在6种植物中最低,可见山合欢主要是通过积累可溶性物质来调节细胞膜的渗透势;余甘子的Pro、可溶性糖含量在6种植物中都是最低,而其3种保护酶活性在6种植物中相对较高,其种子活性相对较高,可见余甘子主要是通过增强保护酶活性来清除种子内部自由基,从而维持活力的;木豆、新银合欢、苏门答腊金合欢、车桑子4种植物抗逆性则是受到可溶性糖、Pro的渗透调节和保护酶系统的双重调控。
本研究中,同一种植物种子的3种保护酶活性高低序列具有一致性,但各种酶达到峰值的时间不相同;不同种植物,酶活性有高低之分,且各种酶达到峰值的时间不同。这充分说明3种抗氧化酶作用过程并不同步,同时6类种子清除自由基的能力也是有差异的。6类种子在处理18~24 d,3种酶基本上都达到峰值并开始下降,这标志着干旱高温胁迫使种子保护酶系统活力降低。可溶性糖在种子处理12~18 d时开始下降,一方面可能是由于干旱高温对种子的胁迫,另一方面可能是由于种子萌芽前期的物质积累所致。总体来看,所研究的6种植物都具有很强的抗逆性,处理后第24天可以认为是各类种子抵御逆境的临界期,因为此时各类种子可溶性糖、保护酶活性明显降低,同时MDA含量明显增大,种子抗逆性的下降引起种子生活力的降低。
植物对逆境的适应性是体现在形态特征、水分平衡、生理代谢等诸多方面,而且随着物候期和生境因子的变化,其适应机制有很大不同[26-30]。本研究仅从生理生化角度初步分析了干热河谷6种植物种子的抗逆机制,由于试验中采用人工模拟自然环境的方法,因此不可避免的忽略了一些生境因子的影响,这有可能导致研究结果与实际状况有所出入。笔者将进一步依托国家林业局云南元谋荒漠生态系统定位研究站资源优势,采用野外定点观测和室内模拟相结合的方法,从植物的形态结构、物质代谢、群落构成等角度,从生态学、遗传学、分子生物学水平来对干热河谷植物的抗逆机制进行深入研究。
[1]马姜明,李昆,张昌顺.元谋干热河谷苏门答腊金合欢、新银合欢人工林天然更新初步研究[J].应用生态学报,2006,17(8):1365-1369.
[2]宋松泉,程红炎,龙春林,等.种子生物学研究指南[M].北京:科学出版社,2005:57-59.
[3]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2003:258-261.
[4]高俊凤.植物生理学实验技术[M].西安:世界图书出版社,2000.
[5]上海植物生理学会.植物生理学实验手册[M].上海:科学技术出版社,1985.
[6]刘宁,高玉葆,贾彩霞,等.渗透胁迫下多花黑麦草叶内过氧化物酶活性和脯氨酸含量及质膜相对透性的变化[J].植物生理学通讯,2000,36(1):11-14.
[7]Sgherri C L,Maffei M,Navari-Izzo F.Antioxidative enzymes in wheat subjected to increasing water deficit and rewatering[J].Plant Physiol,2000,157(3):273~279.
[8]张中华,杨建平,陈圣栋,等.低温胁迫对韭菜膜透性及保护酶活性的影响[J].西北农业学报,2006,15(2):124-127.
[9]缴丽莉,路丙社,白志英,等.四种园林树木抗寒性的比较分析[J].园艺学报,2006,33(3): 667-670.
[10]郭逍宇,董志,宫辉力,等.再生水对作物种子萌发、幼苗生长及抗氧化系统的影响[J].环境科学学报,2006,26(8):1337-1342.
[11]梁炫强,潘瑞炽,周桂元.活性氧及膜质过氧化与花生抗黄曲霉侵染的关系[J].中国油料作物学报,2002,24(4):19-23.
[12]孙文越,王辉,黄久常.外源甜菜碱对干旱胁迫下小麦幼苗膜脂过氧化作用的影响[J].西北植物学报,2001,21(3):487-491.
[13]Yamada M,Morishita H,Urano K,et al.Effects of free proline accumulation in petunias under drought stress[J].J Exp Bot,2005,56(417):1975~1981.
[14]陈英华,严重铃,李裕红,等.盐胁迫下红海榄脯氨酸与活性氧代谢特征研究[J].厦门大学学报:自然科学版,2004,43(3):402-405.
[15]赵福庚,刘良友,章文华.大麦幼苗叶片脯氨酸代谢及其与耐盐性的关系[J].南京农业大学学报,2002,25(2):7-10.
[16]赵江涛,李晓峰,李航,等.可溶性糖在高等植物代谢调节中的生理作用[J].安徽农业科学,2006,34(24):6423-6425,6427.
[17]邵艳军,山仑,李广敏.干旱胁迫与复水条件下高粱、玉米苗期渗透调节及抗氧化比较研究[J].中国生态农业学报,2006,14(1):68-70.
[18]刘亚云,孙红斌,陈桂珠.多氯联苯对桐花树幼苗生长及膜保护酶系统的影响[J].应用生态学报,2007,18(1):123-128.
[19]张玉秀,柴团耀,Gérard Burkard.植物耐重金属机理研究进展[J].植物学报,1999,41(5):453-457.
[20]王启明.铬胁迫对大豆种子萌发中膜脂过氧化作用及体内保护酶活性的影响[J].种子,2006,25(3): 15-18.
[21]路兴花,吴良欢,郑寨生,等.不同生态稻区覆膜旱作稻氮营养生理及抗逆生理特性探讨[J].应用生态学报,2005,16(2):273-278.
[22]许冬梅,王堃,龙澍普.宁夏中部干旱带沙质草地赖草和中亚白草的抗逆生理特性[J].中国草地学报,2007,29(2):31-34.
[23]郭丽红,王定康,杨晓虹,等.外源乙烯利对干旱胁迫过程中玉米幼苗某些抗逆生理指标的影响[J].云南大学学报:自然科学版,2004,26(4):352~356.
[24]阮成江,谢庆良.土壤水分对沙棘成活率及抗逆生理特性的影响[J].应用与环境生物学报,2002,8(4):341-345.
[25]孙方行,孙明高,夏阳,等.NaC1胁迫对紫荆幼苗保护酶系统的影响[J].中南林学院学报,2005,25(6):34-37.
[26]周海燕.荒漠沙生植物生理生态学研究与展望[J].植物学通报,2001,18(6):643-648.
[27]张永恩,李潮海,王群.植物抗旱相关功能基因研究进展[J].中国农学通报,2004,20(6):85-88,113.
[28]郑凤英,彭少麟.植物生理生态指标对大气CO2浓度倍增响应的整合分析[J].植物学报,2001,43(11):1101-1109.
[29]刘璞,陈珈.植物激素脱落酸的信号转导[J].植物生理学通讯,2000,36(2):165-169.
[30]台培东,郭书海,宋玉芳,等.草原地区不同生态类型的植物生理特性的比较研究[J].应用生态学报,2000,11(1):53-56.
Physiological Characteristics of Stress-resistant for Six Kinds of Plant Seeds in Yuanmou Dry andHot Valley of Yunnan
CUI Kai1,LI Kun1,2,SUN Yong-yu1,YANG Wen-yun1,LIU Fang-yan1,LI Li1
(1.Research Institute of Resources Insects,CAF Kunming 650224;2.Yuanmou Desert Ecosystem Research Station of the State Forestry Administration,Kunming 650224)
Abstract:Seeds vigor,respiration,osmotic adjustment,membrane peroxidation and activities ofmembrane protective enzyme system in 6 kinds of plant seeds were studied under April weather of Yuanmou dry and hot valley of Yunnan.The results showed that as the time of seeds in treatment increased,the seed vigor had a decreasing trend and the relative electrical conductivity,Pro content,and MDA content had an increasing trend.Meanwhile,the respiration intensity,SOD activity,POD activity,CAT activity and soluble sugar content had a peak and then declined.The ability of stress-resistant in seeds lay on the respiration,activity of membrane protective enzyme and protectivematter integrated action.A lso,the crucialmatter was different for affecting the ability of stress-resistant in 6 kinds of plant seeds.
Key words:dry and hot valley;physiology of stress-resistant of seed;membrane peroxidation;membrane protective enzyme
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









