



车桑子种子抗逆生理学特性及其对天然更新的影响
张春华1,2唐国勇1,2,孙永玉1,2,刘方炎1,2,李昆1,2,和菊3
(1.中国林业科学研究院资源昆虫研究所,昆明650224; 2.国家林业局云南元谋荒漠生态系统定位研究站,昆明650224;3云南省林业科学研究院,昆明650204)
摘要:车桑子〔Dodonaea viscosa(L.)Jacq.〕是干热河谷植被重要的优势种或常见种。本文通过模拟金沙江干热河谷典型地区元谋盆地4月气候条件下,研究了车桑子种子的活力散失机制及其有关生理生化变化情况,并对车桑子天然群落的自然更新状况进行了调查。结果表明:随着高温干旱处理时间延长,种子活力不断降低,相对电导率、MDA含量呈持续上升趋势;呼吸强度、SOD活性、POD活性、CAT活性、可溶性糖含量呈单峰曲线变化;膜保护酶系统活力降低和保护性物质含量减少是造成种子活力下降的主要因素;实验处理的第12天至第18天是车桑子种子抵御逆境的临界期;每年3月下旬至4月上、中旬,应采取人工辅助措施来提高其种子的天然更新能力。
关键词:干热河谷;车桑子;种子活力;天然更新
中国西南横断山区及临近其东部地区分布的金沙江、澜沧江、怒江和红河流域河谷两岸,海拔1600m(红河河谷为海拔800m)以下地带,由于受到大气环流、地形地貌和地理位置等影响,形成了独特的干热河谷气候类型。在恶劣气候和人为干扰双重压力下该地区原生植被几乎无存,现存植被为稀树灌草丛,许多地方甚至成为单一的禾草丛,或土壤裸露的“光板地”,水土流失严重,泥石流频繁发生,严重威胁各江河中下游地区的人民群众生命财产的安全。在此环境适应生长着一批以热性为主的耐旱植被类型,在其植物群落组成上,多数为热性耐旱的种类,有长期适应干热河谷植物群落的特征种和植物区系标志种。作为植被优势种或常见种,多数为生态适生种或耐干热种,无患子科的车桑子又名明油子〔Dodonaea viscosa(L.)Jacq.〕是其中常见的物种之一[1],集中分布在云南金沙江干热河谷中部两岸,垂直分布于海拔900~2300m,在国内的福建南部、台湾、广东、广西、海南、四川以及国外的热带非州、太平洋群岛亦有分布;车桑子是西南干热河谷重要的本土树种,对该地区植被恢复、水土保持、提高土壤有机碳含量具有重要作用。其种子含油率15.7%,可供制皂,民间还用于点灯,叶研细可治烫伤和咽喉炎,根有大毒可杀虫,全株用于治风湿;由于其喜光、耐旱、耐瘠薄的生态学特性,近年来广泛应用于干热河谷荒山的植被恢复[2]。目前,对车桑子的研究主要集中在造林技术、抗旱机理、光合水分生理特点[2-3]等方面。另外,车桑子在石漠化地区的生态恢复中也有突出表现[4]及造纸等用途[5]。本文主要研究车桑子种子的抗逆性及其对天然更新的影响,旨在揭示保持车桑子种子活力和促进天然更新的制约因素,为人工促进或天然恢复干热河谷退化植被提供科学依据。
1 材料与方法
1.1 供试材料
试验选用车桑子种子于2008年初采自云南省元谋县,采集时选择性状优良母树上成熟、丰硕、无病虫害的果实,晒干暴裂后筛选出饱满种子,并在实验前统一在室内进行1个月的晾晒处理。
1.2 模拟气象资料
试验用气象数据(1996~2008年历年4月1~30日,每天2:00、8:00、14:00和20:004个时段的地面温度、空气湿度、日照时数等)由元谋县气象站提供,各项指标取历年的平均值,用RX智能型人工气候箱进行种子模拟处理。实验主要模拟元谋干热河谷最干热季节4月的地表温度、空气湿度、日照时数(光照强度约为2000lx),实验期30d,具体模拟的气象数据见表1。
表1 1996~2008年元谋干热河谷4月平均气象指标

1.3 种子处理
选取饱满、大小较均匀的种子,经1%NaCl消毒液消毒15min,然后把种子均匀摆放于垫有2层湿润滤纸的培养皿内,每个培养皿内放100粒种子,重复8次,并将其置于人工气候箱内培养,气候箱按上述(1.2)描述的气象资料进行设置。按6d为一个周期,采用TTC法检验种子活力,同时测定种子呼吸强度、相对电导率、丙二醛(MDA)含量、脯氨酸(Pro)含量、可溶性糖含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性等8项生理生化指标。
1.4 测定方法
采用TTC法快速检验种子活力;电导率仪测定相对电导率[6];硫代巴比妥酸(TBA)法测定MDA含量;茚三酮比色法测定Pro含量;氮蓝四唑(NBT)光还原比色法测定SOD活性,以抑制NBT光化还原的50%为1个酶活性单位;愈创木酚法测定POD活性;以每1min内A470nm变化0.01为1个酶活单位;高锰酸钾滴定法测定CAT活性,酶活性以每1g鲜质量样品1min内分解H2O2的“mg”数表示[7]。静置碱液吸收法测定呼吸强度[8];蒽酮比色法测定可溶性糖含量[9]。各项指标的测定均重复3次,取平均值。
1.5 天然更新调查
在元谋黄瓜营和物茂乡选择车桑子集中分布、保存较完好的5片天然林分进行自然更新调查。每片林地面积均在3215hm2以上。土壤类型为燥红土,坡向东南坡和南坡,坡度18°~20°。根据典型随机的原则,在山坡中、下部开花结实植株多的地块,每片分别设置20m×20m样地3个,每个样地分上、中、下3带各均匀设置2m×2m小样方3个。在小样方内进行落果数量和种子散布、发芽和保存情况调查。调查期为2005年1月-2008年4月。1月底至2月初调查样方内的果实数量,3月中、下旬调查种子散布情况,6月下旬调查种子发芽情况,翌年4月调查幼苗保存情况。林分密度4950株/hm2,平均株高1.28m,平均地径3.05cm,开花结实植株占78%。林下植被主要是由扭黄茅(Heteropogon Contortus)、孔颖草(Bothriochloa pertusa)等禾本科草本植物组成,旱季枯死后林下空旷,其中30%的地表裸露,有轻度的水土流失。
1.6 数据处理
采用SPSS13 0对种子活力及各项生理生化指标进行单因素方差分析,指标间进行相关分析。
2 结果与分析
%1.1种子活力、相对电导率和MDA含量的变化
如图1所示,车桑子种子经过模拟气候处理后种子活力显著下降,其变化过程大致可分为缓慢下降期和快速下降期2个阶段。12d以前被处理种子活力缓慢下降,此后进入快速下降期,从12d的79.3%下降至30d的22.4%,其降幅占总降幅的84.93%。单因素方差分析表明,不同时期的种子活力差异显著(P<0 05)。随着处理时间的延长,种子相对电导率呈直线上升趋势,到30d提高了229.31%。种子中的MDA含量亦呈现同样变化趋势,到30d提高了257.94%。
%1.2种子Pro、可溶性糖含量和呼吸强度随处理时间的变化
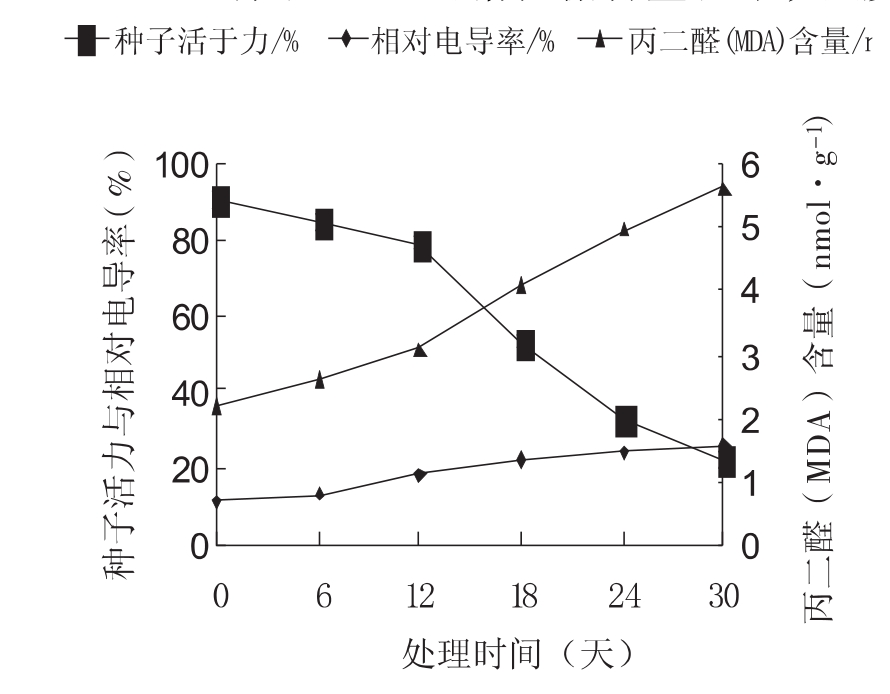
图1 种子活力、相对电导率和MDA含量的变化
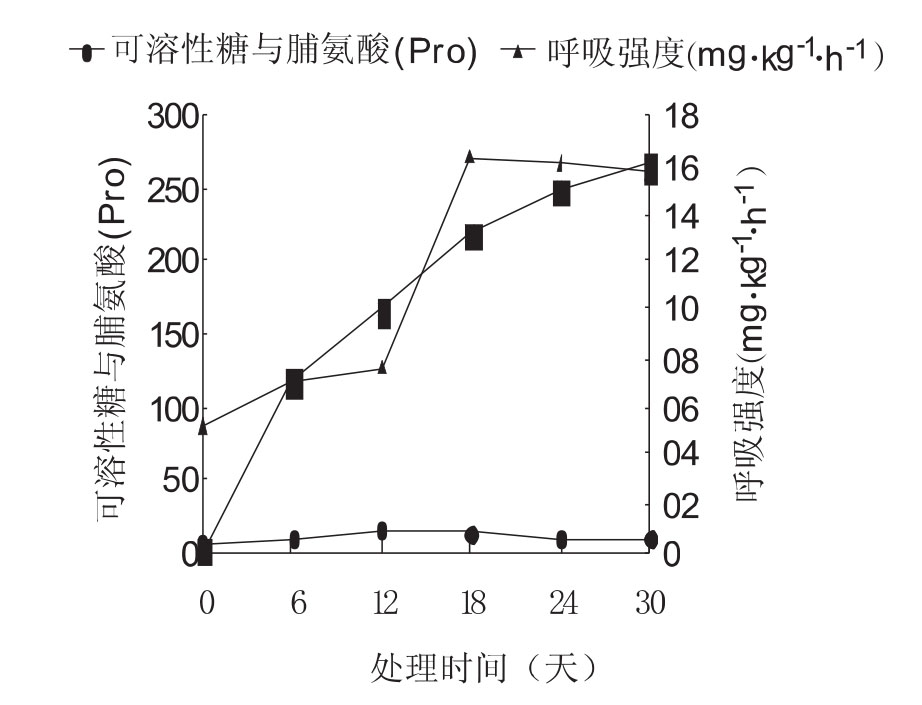
图2 种子Pro、可溶性糖含量和呼吸强度
如图2所示,处理前种子内部没有Pro积累,经过6d处理,Pro的含量急剧从0增加到118.83mg·kg-1·h-1,随着处理时间的延长,其积累量一直不断增加,但6d以后增幅变缓。另外,经过6d处理,种子中的可溶性糖含量立即增加了1.02倍,到12d达到最大值,然后缓慢下降,呈现出急剧上升和相对缓慢的下降趋势。随着试验处理时间的延长,种子的呼吸强度呈现出由缓慢增强到达高峰期,再缓慢降低的单峰格型变化,到第12d时呼吸强度出现最大值,呼吸强度比最初提高了1.8倍。
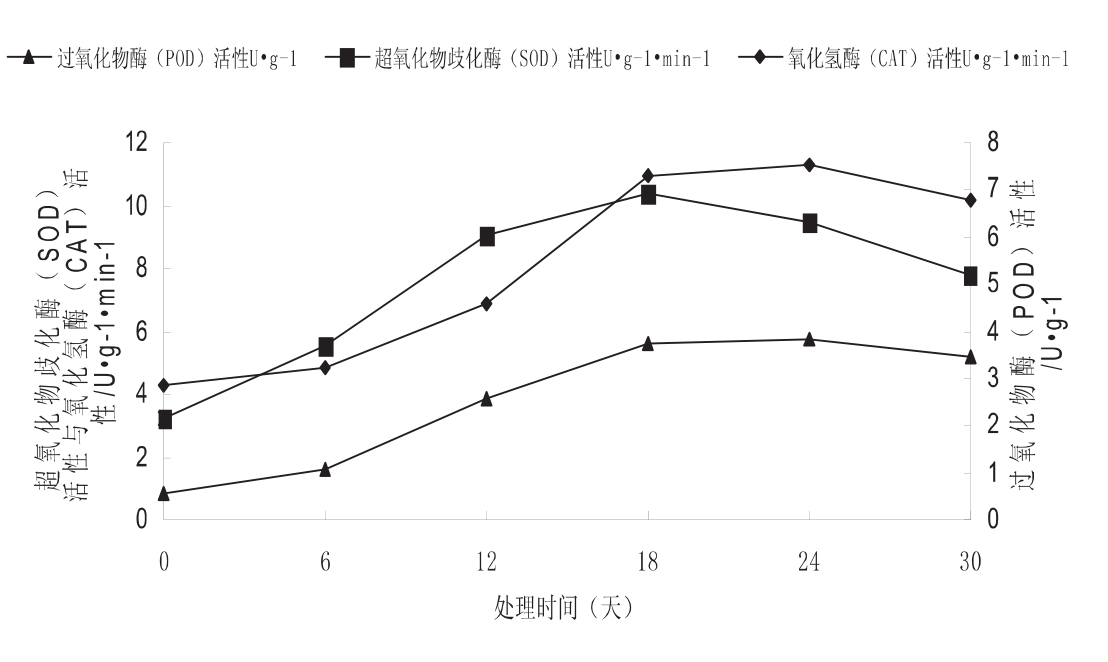
图3种子SOD、POD、CAT活性随处理时间的变化
2.3 种子SOD、POD、CAT活性随处理时间的变化
如图3所示,在30d的处理过程中,车桑子种子的SOD、POD性呈现出单峰曲线变化趋势非常接近,在处理至第18天达到高峰,随后急剧下降;而CAT在处理至第24天时才达到高峰,然后缓慢下,整个变化过程相对比较平缓。
2.4 各项指标间的相关分析
表2列出了经相关分析后各指标间的相关系数,可以看出车桑子种子活力与相对电导率等8个指标呈负相关,其中与相对电导率、相对电导率、MDA含量呈极显著的负相关;与Pro含量、种子POD活性、种子CAT活性都达到显著负相关水平;相对电导率等其他7个指标间的相关性均呈现正相关,其中相对电导率与MDA含量、Pro含量,Pro含量与POD活性,种子呼吸强度与POD活性、CAT活性,POD活性与CAT活性、SOD活性呈现极显著正相关;而相对电导率与种子呼吸强度、CAT活性,MDA含量与POD活性、CAT活性,Pro含量与种子呼吸强度、CAT、SOD呈显著正相关。
表2 各指标间的相关性分析
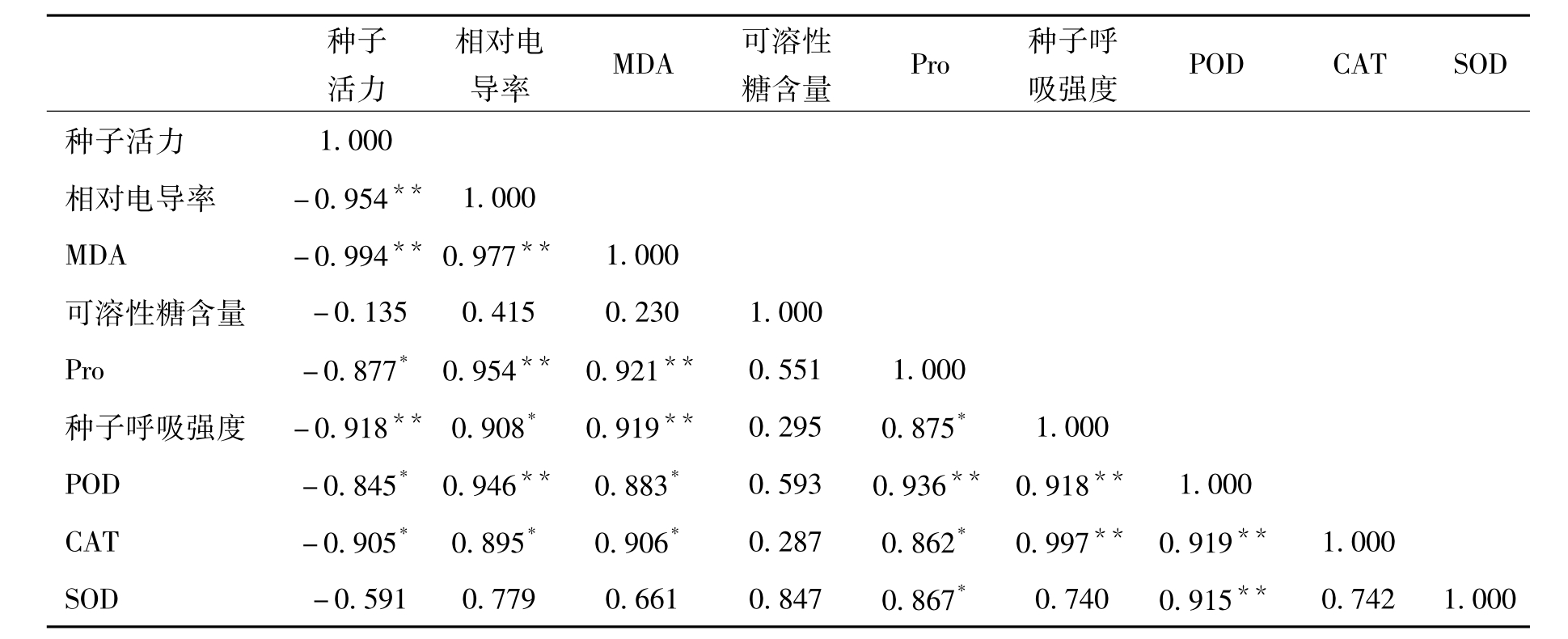
注:*P<0.05显著水平,**P<0.01。
2.5 种子林地天然更新
对车桑子天然更新连续2年的监测结果表明,4m×4m=16m2样方内平均的落地种子数目475粒,仅112粒种子萌发,发芽率仅为23.58%,到雨季结束只有63株成苗,在翌年旱季中有12株在干旱环境中保存下来(见表3)。
表3 车桑样地自然更新情况
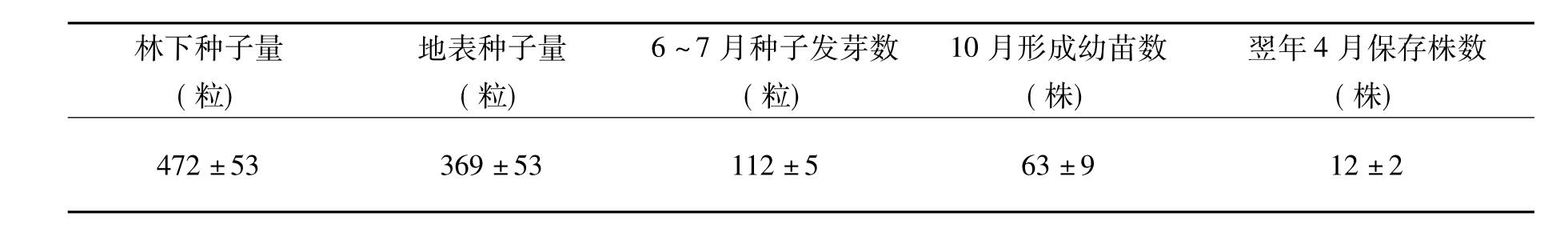
注:平均值±标准差。
2 讨论
暴露于高温干旱环境下的车桑子种子具有一定的抗逆能力,在实验处理的12d内,发芽率仅以几个百分点的速率缓慢下降,而保护酶、可溶性糖和呼吸强度有大幅度的变化,车桑子种子的抗逆能力已达极限,生活力也将快速散失。但从高温干旱逆境处理后的变化来看,到18d出现高峰时,呼吸强度比开始实验时提高了179.3%,此后呼吸强度减弱,其变化趋势与种子生活力非常类似,可能是由于种子生活力的下降导致了呼吸强度降低。
与植物抗旱性与SOD、POD和CAT活性增强幅度成正相关,而与MDA含量呈负相关[10]不同的是车桑子种子的抗逆性与SOD、POD、CAT、MDA活性、相对电导率、MDA含量、种子呼吸强度和可溶性糖含量呈显著正相关。反映出车桑子种子抵御高温干旱逆境的能力是其生理代谢过程综合作用的结果,其中包括了细胞膜保护酶系统、种子呼吸代谢和多种渗透调节物质等。逆境条件下,植物O2-产生的速率随胁迫强度加大而递增,致使细胞内的O2-代谢失去平衡,出现O2-过剩。这些过剩的O2-可在细胞内诱发自由基连锁反应而产生H2O2·OH等更多的自由基和活性氧,从而氧化膜细胞中不饱和脂肪酸的双键,使之断裂、分解而破坏[11]。POD可将H2O2分解为H2O2和O2,CAT也有此作用,CAT活性可能影响SOD和POD活性变化[12]。实验处理18~24d的车桑子种子,3种抗氧化酶活性先后达到峰值,其后下降。表明随着高温干旱逆境胁迫时间延长,种子保护酶系统遭到破坏,3种酶活性与胁迫程度成比例地下降,对种子活力的保护功能亦已开始迅速减弱。在通常状况下,SOD和POD活性变化是同步的,本研究结果与之相同。可溶性糖含量下降时间出现在实验处理12d以后,12~18d为缓慢下降期,含量下降时间稍早于SOD、POD和CAT等抗氧化酶。可溶性糖的积累是植物对干旱逆境胁迫的一种适应性反应,其在细胞中的积累对稳定渗透调节能力的相对贡献比较大[13]。但植物的渗透调节能力和调节幅度是有限的,并受到水分胁迫程度、发展速度和其他环境条件的影响;通常水分胁迫发展快或严重时,渗透调节能力变小,甚至消失,并常常在几天内消失[14]。可溶性糖作为一种渗透调节物质,可能在车桑子种子遭遇高温干旱逆境胁迫初期(12d内)迅速积累,有利于保护种子生活力,随着峰值的出现,可溶性糖含量降低,其保护功能很快消失,并预示着有关保护酶系统将遭受破坏,种子活力也将大幅下降。在本研究中,这种变化的指示作用比较明显。在相同的环境胁迫条件下,耐旱树种或品种比不耐旱的具有较低的电解质外渗率[15,16],环境胁迫强度直接影响相对电导率大小。MDA是具有细胞毒性的物质,可与膜结构的蛋白质和酶结合、交联而使之失去活性,从而破坏膜结构[11],因此MDA含量常常与植物抗旱性呈负相关[10]。本研究的实验处理过程中,随着高温干旱逆境胁迫的持续,车桑子种子的相对电导率和MDA含量变化表现增加的趋势,到30d相对电导率提高了239%,MDA含量提高了257%;与此相反,种子活力却下降了77.6%,相对电导率和MDA含量的这种变化趋势,明显与种子活力的变化相反。反映出在高温干旱逆境胁迫到达一定程度后,种子细胞膜遭受严重损伤,质膜透性明显增强,MDA在种子中的富积致使种子活力大幅下降。另外,与相对电导率和MDA含量的变化趋势相似,实验处理的种子中Pro含量一直在持续增加,在18d后与SOD、POD、CAT3中抗氧化酶活性和可溶性糖含量之间表现出相反的变化趋势,Pro的积累更像是种子受害后的症状和反应,或是仅仅反映出高温干旱逆境胁迫的程度。游离脯氨酸的积累在许多植物中都被观察到[17],但有研究认为水分胁迫下植物体内游离脯氨酸的积累是个普遍现象,甚至认为可以作为植物抗旱性和育种指标[18]。不过,也有许多学者认为[17,19~21],游离脯氨酸的积累速率和积累量与植物抗旱性之间并不存在一个简单的关系。也有研究结果表明,脯氨酸积累的数量的多少只是反映了水分胁迫的程度,与植物的抗旱性并不无相关性[22]。在本研究中游离脯氨酸的持续积累,与种子活力从逐步散失转为快速散失的变化趋势差异较大,很难与植物的抗旱性联系在一起,但与薛吉全等的研究结果相一致[22],其持续增加的趋势反映余甘子种子遭受高温干旱逆境胁迫的程度。动物取食或病原侵害、人为干扰影响、种子休眠特性和生境是影响植物种子天然更新的4大因素[22~25]。在元谋干热河谷有如仓鼠科(Circetidae)、鹿科(Cervusaxis)、兔科(Leporidae)等的一些动物会取食车桑子种子,种子无明显的休眠特性。本文的调查样地只有轻度的人为干扰,4大因素中的前3个基本可以排除,调查结果表明:现存种子萌发率和幼苗保存率非常低,所以生境是种子萌发率低的重要因素。干热河谷的干湿季节非常明显,旱季高温干旱,从当年的11月到次年的5月,时间长达7个月。在此期间,降雨量仅占全年总降雨量的10%左右,即使雨季来临,间歇性干旱也频繁发生,少则10d,多则接近30d。车桑子种子通常雨后立即吸涨萌发,如遇间歇性干旱,刚萌发的胚根干枯,尤其是落在“光板地”上萌发的种子,对于车桑子的繁殖更新将是灾难性的。个别掉落于枯枝落叶或石缝中的种子,即便萌发后未立即遭受恶劣环境的影响,但由于土壤板结,幼根很难迅速扎入深层土壤,吸收足够的水分和养分,幼苗难以正常生长发育,因而也很难保证其成活率和保存率。从试验结果可看出,经历4月的恶劣气候影响后,车桑子种子的活力降至22.4%。调查表明:进入雨季后(6月)车桑子林内实际种子萌芽率仅为23.58%。可见,经过5月恶劣气候的影响,种子活力大幅度降低,大致只有4月底时的1/2左右。综合以上分析,高温干旱是制约干热河谷区车桑子种子天然更新的关键因素。在当地4月份的气候条件下,若车桑子种子裸露于阳光暴晒的“光板地”表,其活力保持时间大约为12~18d,可以认为,这是其抵御当地高温干旱逆境的临界期。在干热河谷现存的车桑子天然林分内,即使有较多的植株开花结果,也有一定数量的种子保存于林地内,但其天然更新效果较差,落于林地内的种子萌发率不到40%,能成幼苗的只有15%左右,而正常生长发育的苗木数量很少。因此,提高其种子发芽率,促进干热河谷植被的恢复,在进一步加强天然林保护措施的同时,还必须采取提高林地覆盖率、松土除草、集水保水等人工辅助措施,而且必须在果实掉落晒干爆裂,种子被弹出落地后进行,实施时间约在3月中旬至4月上旬。这样可使弹出的种子掉落于疏松的土壤中,防止种子直接暴露于高温干旱的环境下,更长时间地使种子保持正常的生活力,并为种子发芽和幼苗生长提供良好的环境条件。
[1]金振洲,欧晓昆.干热河谷植被[M].昆明:云南大学出版社,云南科技出版社,2000.
[2]马焕成.干热河谷造林新技术[M].昆明:云南科技出版社,2001.
[3]赵琳.云南干热河谷旱季车桑子的光合水分生理特性的研究[J].西部林业科学,2006,35(1):12-17.
[4]罗蔚,李秀兰,靳灵江.车桑子在石漠化治理中的优势及推广[J].种子,2006,25:109-110.
[5]张进光.结合西部大开发战略利用车桑子造纸[J].实用技术,2002(2):24.
[6]宋松泉,程红炎,龙春林,等.种子生物学研究指南[M].北京:科学出版社,2005.
[7]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2003.
[8]高俊凤.植物生理学实验技术[M].西安:世界图书出版社,2000.
[9]上海植物生理学会.植物生理学实验手册[M].上海:科学技术出版社,1985.
[10]丁文蒋,海淑珍,徐柱,等.牧草种质资源抗旱性鉴定方法的初步研究[J].草业科学,1992,9(5): 57-60.
[11]陶晶,陈士刚,秦彩云,等.盐碱胁迫对杨树各品种丙二醛及保护酶活性的影响[J].东北林业大学学报,2005,33(3):13-15,37.
[12]王艳芳,韩冰,张占雄.锡林郭勒草原克氏针茅抗旱性生理变化的研究[J].草业科学,2006.23(2): 22-26.
[13]张莉,续九如.水分胁迫下刺槐不同无性系生理生化反应的研究[J].林业科学,2003,39(4):162-167.
[14]李吉跃.植物耐旱性及其机理.北京林业大学学报[J].1991,13(3):92-100.
[15]刘彦琴,张丰雪,杨敏生.电导率在白杨杂种无性系耐旱性鉴定中的应用[J].河北林果研究,1997,12(4):301-305.
[16]章崇玲,曾国平,陈建勋.等.干旱胁迫对菜苔叶片保护酶活性和膜脂过氧化的影响[J].植物资源与环境学报,2000,9(4):23-26.
[17]王邦锡,黄久常,王辉.不同植物在水分胁迫条件下脯氨酸的累积与抗旱性的关系[J].植物生理学报,1989,15(1):45-51.
[18]高玉葆.植物对于水分胁迫的适应性反应及其生态学意义.见:李博.现代生态学讲座.北京:科学出版社.1995.
[19]汤章城.逆境条件下植物脯氨酸的累积及其可能的意义[J].植物生理学通讯,1984(1):15-21.
[20]曹仪植,吕忠恕.水分胁迫下植物体内游离脯氨酸的累积及其ABA在其中的作用[J].植物生理学报,1985,11(1):9-16.
[21]胡新生,王世绩.树木水分胁迫生理与耐旱性研究进展及展望[J].林业科学,1998,34(2):77-89.
[22]薛吉全,任建宏,马国胜,等.玉米不同生育期水分胁迫条件下脯氨酸变化与抗旱性关系[J].西安联合大学学报:自然科学版,2000,3(2):21-25.
[22]GorchovDL,CornejoF,JaramilloAM.TheroleofseeddispersalinthenaturalregenerationofrainforestafterstripcuttinginthePeruvianAmazon[J].Vegetatio,1993,107/108:339-349.
[23]李宏俊,张知彬.动物与植物种子更新的关系.Ⅱ.动物对种子的捕食、扩散、贮藏及与幼苗建成的关系
[J].生物多样性,2001,9(1):25-37.
[24]彭闪江,黄忠良,彭少麟,等.植物天然更新过程中种子和幼苗死亡的影响因素[J].广西植物,2004,24(2):113-121.
[25]刘足根,姬兰柱,郝占庆,等.松果采摘对长白山自然保护区红松天然更新的影响[J].应用生态学报,2004,15(6):958-962.
Physiological characteristics ofDodonaea viscosa(L.)
Jacq.Seeds stress-resistance and their influences on nature regeneration
ZHANG chun-hua1,2,TANG guo-yong1,2,
SHUN yong-yu1,2,LIU fang-yang1,2,LI kun1,2,HE ju3
(1.Research Institute of InsectsResources,CAF,Kunming,650224;2.Yuanmou Desertification Ecosystem Research Station,State Bureau of Forestry,Yunnan,Kunming,650224,China;
3.Yunnan Academy 0f Forestry,Kunming Yunnan 650204,P.R China)
Abstract:Dodonaea viscosa(L.)Jacq isthe dominantand constant species of the vegetable in xerothemeic valley,W ith the simulation of the climate in April in yuanmou basin,a type area in the xerothemeic valley of jinshaRaver.Thise paper study on the vigor dissipation of Dodonaea viscosa(L.)Jacq seeds,and related physiological and biochemicalmechanisms.The natural regeneration of Dodonaea viscosa(L.)Jacq population was also investigated.The result showed that with prolonging high temperature and drought,the seed vigor decreased,the relative electrical conductivity,Pro content,and MAD content in seed had an increasing trend,and the respiration intensity,activity ofSOD,POD,CAT and soluble sugar content in seed showed a single-peaked pattern.The decrease ofmember protective enzyme activities and of protectivematter contentwas the key factory for the decrease of seed vigor.The period from 12th to 18th day of stress treatment was the critical period for the Seeds stress-resistance ofDodonaea viscosa(L.)Jacq seed.In order to improve the nature regeneration ability of seed,some artificialmeasures should be taken from the last ten days ofApril to early andmidMarch of the year.
Key word:yuanmou xerothemeic valley;Dodonaea viscosa(L.)Jacq;seed vigor;nature regeneration
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










