



18.3.1 转SOD基因对农作物的抗逆研究
目前SOD基因已被克隆和转化。Cu,Zn-SOD基因早在1987年就被克隆了,1998年又先后从菠菜、矮牵牛的叶绿体中克隆到Fe-SOD,1989年从Nicotiana Plumbaginifolia克隆出线粒体Mn-SOD基因,这都为SOD基因转化提供了基础。近年来,利用SOD氧化代谢基因工程提高植物对环境胁迫的耐性已有许多研究。生理学研究表明,抗氧化醛类是植物抵抗氧化胁迫的关键因素,其主要依据有:①逆境胁迫引起抗氧化酶活性增加,且增加的酶活性与耐性直接相关;②用一种胁迫对植物预处理可产生对其他不同胁迫的交叉耐性(crosstolerance),如对百草枯(paraquat,除草剂)有抗性的品系,其抗氧化酶活性增加并对热激、冷害等具有交叉耐性。因此,为提高植物抗氧化能力而进行的基因转移多集中于植物中高表达单个抗氧化酶类基因,其中最常用转化的是编码SOD和GR的基因,这除了它们在抗氧化胁迫中的重要作用外,还因为在细菌和动物系统中的相关研究已积累了相当的基础。现在所研究的转SOD基因植物中,基因产物得以正确的转录及表达,表达量比起始材料提高了1~50倍不等。然而,在胁迫试验中,这些转基因植物表现出了抗性的差异,有的提高了对胁迫的抗性;有的则没有明显的变化。
对氧化逆境的研究引发了利用基因工程对氧化代谢过程进行修饰使植物获得对多种胁迫具有普遍抗性(generalized resistance)的构想,转单个基因得到的保护效果使其可行性得以肯定,但转基因植物在胁迫试验中的不同反应也使我们看到在氧化代谢的生理、生化及调控机制还没有完全搞清楚的情况下,这一理论设想距实际应用还有相当的距离。从目前对氧化代谢过程的了解来看,用分子操作加以改造至少可以从以下三个方面进行。
首先,在一定浓度范围内提高活性氧含量,使其作为毒性分子直接杀死病原,或促进植物体内的信号传导、打开防卫系统、从而提高抗逆性。Wu等在马铃薯中提高SOD含量获得的对软腐病、晚疫病的抗性显示了这一策略的可行性。植物在进化过程中已经形成了对逆境的各种保护机制,如AOS清除系统及防卫体系等。植物之所以被各种环境胁迫所伤害,常常是由于这些防卫系统不能及时启动,因此,通过加强信号传导使植物整个防卫系统快速启动将具有整体效果,其优越性比改变防卫系统内的单个成分应更为显著。然而,这一途径有两个问题必须解决:第一,由于AOS对生物有毒性的一面,因此,其提高的量对受体植物而言应有一定的上限,组成型的超表达可能并不是最理想的,分离诱导表达并快速启动的特异启动子同样十分重要;第二,氧负离子究竟如何完成信号传导过程及促进防卫基因的表达也有待进一步阐明。
其次,就目前状况看,表达结构基因仍是主要的可行对策。为了维持抗氧化系统的平衡,可以考虑两个或多个基因同时表达,如SOD和H2O2清除酶类基因等。这方面的研究在植物中虽未见报道,但在动物研究方面,同时表达SOD和CAT基因,使果蝇寿命延长了11年,且蛋白质氧化损伤程度降低、体征衰老延迟。在这一途径中,值得继续深入研究的是,究竟哪种SOD和氧负离子清除酶类组合才是最佳组合,植物中多种同工酶同时存在,由不同基因编码,每一种形式的确切功能仍不是十分清楚。利用反义RNA技术创造转基因植物功能缺失突变体将会为弄清酶功能带来新的进展。此外,分析这些结构基因启动子区的调控元件及基因产物的亚细胞定位也是影响表达和生物学效果的因素。
最后,在抗氧化酶类基因的调控上加以修饰。前文提到生物体内SOD与氧负离子清除酶类的平衡是提高抗性的一个先决条件。已知对百草枯有抗、感两种类型,抗性生物型中的SOD、APx、GR含量与敏感型相比普遍升高,且抗、感生物型之间呈现结果一致:F1代均抗百草枯,三种酶的活性与抗性亲本一致;F2代抗、感呈3∶1分离,抗性株中的SOD、GR含量均比敏感株系高(因缺乏APx抗体,故文中没有分析F2群体的APx水平),这表明对百草枯的抗性受单个核基因控制。此外,原核系统中氧诱导醛类和氧负离子诱导醚类部分重叠的现象也暗示抗氧化系统的调控可能有共同的机制。因此分离抗氧化系统的调控基因也不失为一条可考虑的途径。但因植物抗氧化系统的基因调控十分复杂,因此,分离其调控基因并加以利用将依赖于植物抗氧化系统分子调控机制的研究进展。
目前国内外已有许多培育SOD基因高表达(SOD活性显著增强)的转基因植物的研究,此类转基因植物提高了对各种逆境环境的抗性,下面分别加以介绍。
18.3.1.1 转SOD基因对作物抗旱、抗盐等逆境的研究
水是植物及其他生物赖以生存的物质基础及先决条件,植物的各种生理活动都离不开水分。但自然界中水资源是有限的,水资源短缺是目前公认的一个全球性焦点环境问题,全世界每年干旱造成的经济损失超过其他自然灾害的总和,成为农业生产的主要自然灾害。干旱灾害具有普遍性、区域性、季节性和持续性的特点,是植物生长最大的不良特点。研究干旱对植物的伤害及植物对干旱的适应和调节机制,通过人为手段培育高效抗旱新品种,是一条经济有效的途径。人们研究发现植物抗旱性是由多基因控制的,是多方面、多层次、多途径共同作用的结果。植物在遭受干旱胁迫时,会通过产生一些调节渗透压物质、抗逆蛋白而增强抗旱性;同时发现作物在干旱、盐胁迫条件下SOD的表达量有不同程度的提高,从而提高对这些不良条件的耐受性。因此可以认为提高作物品种SOD的表达量也可相应提高对这类不良环境的抗性。
人们试图通过基因工程技术提高植物的SOD活性以提高植物的抗逆性。1989年Bowler等首次将烟草的Mn-SOD基因导入到苜蓿中,发现转基因植物SOD活性增强。随后,烟草的Mn-SOD基因再次被分别导入到苜蓿与棉花中,结果表明SOD基因的过量表达能明显提高转基因植株的SOD活性和抗冻性。目前,已将不同来源的Cu,Zn-SOD基因成功导入烟草和西红柿。研究者发现,豌豆SOD基因表达不仅提高了转基因烟草的SOD活性,而且也增强了转基因植株抵抗光抑制的能力;另外,一些研究表明Mn-SOD或Fe-SOD基因在烟草和玉米叶绿体中的过量表达能够增强转基因植株对质膜和光系统Ⅱ的保护作用。McKersie等将烟草Mn-SOD基因通过根癌农杆菌转移到苜蓿基因,通过3年的大田试验,表明转基因苜蓿的产量和存活率都显著提高,支持了抗氧化胁迫程度的提高可以改进作物抗逆性的假说。国内覃鹏、刘飞虎等研究了转SOD基因对烟草抗旱性和相关生理指标的影响,供试材料为从国外引进的烟草叶等基因系,即非转基因品系(non transgenic line,简写Non)、转Fe-SOD基因品系(Fe-SOD gene transgenic line,简写Fe-SOD)和转Mn-SOD基因品系(Mn-SOD gene transgenic line,简写Mn-SOD)。通过用浇水来控制干旱程度,然后用NBT光还原法测定SOD活性、愈创木酚法测定POD活性、硫代巴比妥酸法测定丙二醛和可溶性糖含量、考马斯亮蓝(G250)法测定蛋白质含量、茚三酮法测定脯氨酸含量、电导率法测定叶片质膜透性、LI26400便携式光合仪测定光合作用、干重法测定叶片含水量说明抗旱情况,结果显示:外源Mn-SOD基因的导入能切实提高烟草抗旱能力,而导入的Fe-SOD基因虽能提高烟草体内的SOD活性水平,但不能提高烟草的抗旱性,说明Mn-SOD可能与烟草的抗旱性关系较大;当遭受干旱胁迫时,所导入的两种SOD基因可能在某种程度上影响了植物体内MDA、蛋白质、光合作用以及脯氨酸等的正常生理代谢;脯氨酸、MDA、蛋白质等生理指标的变化在抗旱与不抗旱品系之间并没有表现出明显的规律性,因此能否作为抗旱育种的重要指标应该通过更多的试验来确定。
18.3.1.2 转SOD基因对作物抗高温逆境的研究
刘明求、刘齐元等研究了转基因SOD,POD高表达烟草幼苗抗热性影响,以转基因Mn-SOD高表达烟草品系(简称Mn-SOD高)、转基因POD高表达烟草品系(简称POD高)和非转基因烟草对照品系(简称非转基因)。经过对高温条件下转基因系及对照组生物量及生理生化指标的测定,结果表明,转基因品系SOD的活性更强,丙二醛的含量更低,生物量受到的影响更小。由此可认为,转基因烟草品系幼苗比非转基因品系的抗热性更强,其中转基因POD高表达烟草品系的抗热性最强,研究结果表明通过转基因方法提高植物的抗热性是可行的。
18.3.1.3 转SOD基因对作物冷害逆境的研究
我国幅员辽阔,低温伤害是限制植物自然分布和栽培区带的主要因素,也是全国性的自然灾害之一,每年因植物受低温伤害而造成了巨大的经济损失。因此,研究低温胁迫对植物的伤害作用及其机理,探索植物抗冷冻机制及其预防措施,对于引种栽培具有指导意义。在低温冷冻条件下,植物叶中NO大量合成,诱导抗氧化酶活性增强,抗氧剂积累,及时清除氧自由基防止膜脂过氧化,是维持细胞膜完整性、保证其安全越冬的重要生理反应。
随气温下降,过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸氧化酶(APx)活性和谷胱甘肽(GSH)、脯氨酸(Pro)、类胡萝卜素(Car)含量增加并在冬季达最大值,NO释放率、NOS和NR活性以及丙二醛(MDA)含量也显著增加,并在秋季和初春出现两峰值;两树种叶中的抗氧化酶活性和抗氧剂含量都与温度和MDA含量明显负相关,表明抗氧化系统在清除氧自由基、抑制膜脂过氧化、提高抗冷冻性方面具有重要作用;而且NO的产生总是发生在抗氧化酶活性和抗氧剂含量增加之前,表明NO作为信号分子激活了抗氧化系统以诱导植物的抗冷冻性。在抗冷冻过程中祁连圆柏叶的NO释放率、NOS和NR活性、抗氧化酶活性以及抗氧剂含量都高于圆柏,表明祁连圆柏具有较强的抗冷冻适应性策略。
Mckersie等研究了烟草的Mn-SOD cDNA转入苜蓿中对越冬存活性的影响,为了检测活性氧提高冬季存活率,两个苜蓿品种进行了转Mn-SOD基因研究,得到转基因植株总SOD酶活性是对照的2倍,提高了冷害的耐受性。豌豆叶绿体Cu,Zn-SOD转化到烟草得到的转基因植株的SOD活性提高3倍,低温强光下产物也会增加。
Ashima Sen Gupta,Jeanie L.Heinen等研究了转基因Cu,Zn-SOD植物在低温及强光照射对产量的影响,采用豌豆叶绿体来源SOD转入烟草中,将该转基因植物放置于冷室及不同光强条件环境中,当在适度光照中转基因植株利用光照能力比对照条件下与对照不同,转基因植株包含几乎90%的光合能力,同时这些转基因植株也显示了对产生超氧化物除草剂的抗性,可降低光影响下对细胞的损伤。
18.3.1.4 转SOD基因对作物早衰的研究
目前在许多作物的育种工作中早熟性是主要的目标性状之一,但早熟性一般会伴随着早衰现象的出现,而早衰则影响作物的产量、品质、抗性等,因此在育成早熟品种的同时必须提高其抗早衰的性状。植物体内抗氧化酶SOD活性的提高能增强植物对多种氧化胁迫的抗性,而现在关于早衰的研究还很少。喻树迅等研究发现,SOD与棉花的早衰有着密切的关系,早衰品种SOD酶活性显著低于不早衰品种。基于这一点,马淑娟、喻树迅研究了异源SOD对烟草耐衰老影响,构建了棉花叶绿体Cu,Zn-SOD基因的植物表达载体,利用农杆菌介导法将其导入NC89烟草,PCR、Southern blotting检测结果显示有4个烟草株系中整合了棉花叶绿体Cu,Zn-SOD基因,SOD酶活性测定结果显示4株转基因烟草植株SOD酶活性都明显高于非转基因烟草,离体叶片暗处理试验结果也证明4株转基因烟草比非转基因烟草SOD酶活下降速度明显减慢,百草枯喷施试验表明,转基因烟草都比非转基因烟草对百草枯的耐性增强,这一系列试验间接地说明外源基因的导入增强了烟草的耐衰老能力。
18.3.1.5 转SOD基因对作物抗病害逆境的研究
植物在病原物的侵害或不适环境条件的影响下生理机能失调、组织结构受到破坏,植物病害是寄主植物和病原物的拮抗性共生,其发生和流行是寄主植物和病原物相互作用的结果。农作物和林木的病害大爆发,常使国家经济和人民生活遭受严重损失,有些患病作物能引起人畜中毒。在植物与病原物长期进化过程中产生了一系列抵抗病原物机制,分为以抗病基因为主的主动抗病体系及以防御基因为主的被动防御体系。超氧化物歧化酶是植物细胞内防御酶系统的重要成员之一,其生理作用是歧化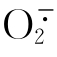 产生H2O和O2,在植物与病原物相互作用过程中,植物体内SOD活性发生变化,不同的植物病害系统或不同的相互作用类型,其SOD活性变化是不同的。烟草受TMV感染时,叶片组织中SOD活性上升,且过敏反应组织中的SOD活性比感病组织中高。烟草接种青枯病菌,接种后与接种前比较,抗病品种的SOD活性增长明显大于感病品种,证明了SOD活性与品种抗性呈正相关。烟草低头黑病菌接种烟草后,SOD活性表现为先升高后降低,而且在整个处理阶段抗病品种的酶活高于感病品种。烟草感病初期叶片SOD活性下降,随后升高,但是酶活性仍然表现为抗病品种高于感病品种。烟草接种黑胫病后抗病品种的SOD活性明显低于感病品种。植物受到病害侵染后,SOD活性会升高,说明病害侵染产生的会激发植株体内SOD合成酶基因的表达,表现为SOD活性上升。而感病品种的酶活性低于抗病品种可能是因为感病品种受到病原物侵染后积累的过多造成受害植株SOD防御酶体系崩溃,导致SOD活性降低。至于接种后感病品种SOD活性快速上升而高于对照,可能是因为这种高水平的酶活性可以清除植物-病原物亲和性互相作用产生的超氧阴离子,使感病品种受到病原物侵染后不发生过敏性反应,病原物可以在寄主内扩展,从而表现为感病。
产生H2O和O2,在植物与病原物相互作用过程中,植物体内SOD活性发生变化,不同的植物病害系统或不同的相互作用类型,其SOD活性变化是不同的。烟草受TMV感染时,叶片组织中SOD活性上升,且过敏反应组织中的SOD活性比感病组织中高。烟草接种青枯病菌,接种后与接种前比较,抗病品种的SOD活性增长明显大于感病品种,证明了SOD活性与品种抗性呈正相关。烟草低头黑病菌接种烟草后,SOD活性表现为先升高后降低,而且在整个处理阶段抗病品种的酶活高于感病品种。烟草感病初期叶片SOD活性下降,随后升高,但是酶活性仍然表现为抗病品种高于感病品种。烟草接种黑胫病后抗病品种的SOD活性明显低于感病品种。植物受到病害侵染后,SOD活性会升高,说明病害侵染产生的会激发植株体内SOD合成酶基因的表达,表现为SOD活性上升。而感病品种的酶活性低于抗病品种可能是因为感病品种受到病原物侵染后积累的过多造成受害植株SOD防御酶体系崩溃,导致SOD活性降低。至于接种后感病品种SOD活性快速上升而高于对照,可能是因为这种高水平的酶活性可以清除植物-病原物亲和性互相作用产生的超氧阴离子,使感病品种受到病原物侵染后不发生过敏性反应,病原物可以在寄主内扩展,从而表现为感病。
18.3.1.6 转SOD基因稳定性
外源基因在转基因植物中的表达水平及其遗传稳定性是基因工程育种者最关心的问题,也是转基因植物安全性评价的一个方面。影响转基因作物外源基因表达和遗传特性的研究较多,而关于转外源SOD基因进入植物中的稳定性表达还不明确。对转SOD和APx基因烟草(Nicotianatabacum L.)的外源SOD表达水平及其在不同世代的遗传稳定性研究发现,高表达亲本(Fe-H和Mn-H)为母本的杂交后代SOD活性水平高,低表达亲本(Mn-L)为母本的杂交后代SOD活性水平亦低;高表达杂种的SOD活性比高表达亲本的活性略有提高;杂交后代可以保持外源SOD基因高表达的特性,高表达特征可以在世代间稳定遗传。转基因SOD高表达品系的自交后代分析结果表明,从S0(自交亲本)到S3代,外源SOD基因表达稳定,表现在各自交世代之间、自交世代与其亲本之间的SOD活性基本上稳定,世代间差异不显著,而且都稳定高于非转基因对照品系。因此基于自交后代酶活性水平的稳定性分析,转基因烟草中的外源SOD基因表达水平高,而且这种高表达特性可以在后代中稳定遗传。
有些转SOD基因对抗逆性没有提高作用,Tepperman等报道获得高效产生叶绿体Cu,Zn-SOD的转基因烟草对百草枯抗性并没有增强,以及Van等报道转入的烟草和玉米对冷害和盐胁迫耐受性并没有提高。因此,虽然SOD基因过量表达,但不一定都能提高植物的抗性,并且只是在一定程度上可提高转基因植物对氧胁迫的耐受性,但这种提高有限。提高植物体内抗氧化酶活性及增加抗氧化代谢的水平是抗高植物抗逆性的有效途径之一,但各种保护酶存在部位不同,以及各种代谢产物间既协同又相互制约,所以单纯地转化某一基因使之过量表达很难达到预期的转化目地,Bowler等建议,同时增强SOD、POD及CAT活性可能会更好地提高植物的抗氧化胁迫能力。Kwon以Mn-SOD、Cu,Zn-SOD和APx单独或同时转入烟草,结果表明转入三个基因的比转入两个或一个基因的更能抗TMV引起的氧化胁迫作用。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










