



(一)抗盐机制
王宝山、赵可夫(1997)认为作物耐盐机理是一个十分复杂的问题,但通过大量研究表明,绝大部分作物主要通过拒盐方式来抵抗盐害,即在盐胁迫下,作物地上部分特别是叶片中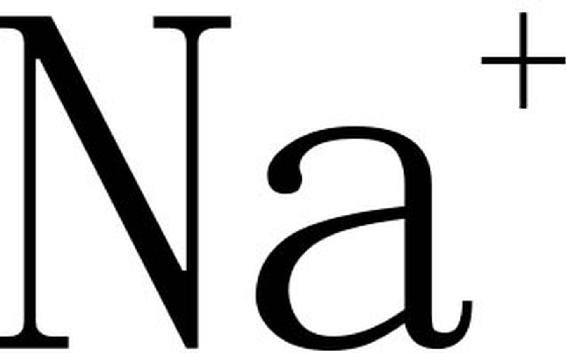 是根中的几分之一,甚至几十分之一。因此,作物的耐盐性与盐的吸收、运输、分配、再循环、排泄及细胞水平上的区域化有密切关系。
是根中的几分之一,甚至几十分之一。因此,作物的耐盐性与盐的吸收、运输、分配、再循环、排泄及细胞水平上的区域化有密切关系。
小麦、甜菜和玉米等在一定范围NaCl 胁迫下,根中比地上部分高3~7倍。我们的结果表明100mmol/ L NaCl 胁迫7天,高粱根和茎基部木质部液中 比穗轴木质部液中高十几倍(王宝山等,1997)。因此,叶片中也远低于根和茎基部。说明许多作物有显著的排机制,目前人们认为作物拒机制涉及以下过程。
(1)作物不将吸入根细胞内,即使进入细胞又通过/H+泵再排出,这可能与作物及品种细胞质膜组成等有关。如相同NaCl 胁迫下,耐盐品种高粱根中明显低于不耐盐品种。
(2)作物把吸收的贮存于根、茎基部、节、叶鞘等薄壁细胞发达器官组织中,而且将 封闭在这些细胞的中央液泡中,从而阻止向叶片中运输,这在高粱中已得到证明。
(3)吸收的在木质部中向上运输过程中被木质部或韧生部传递细胞吸收,并分泌到韧皮部中再运回根中,最后分泌到环境中,即作物通过脉内再循环把再运到体外,这在豆(Drew 等,1987)和高粱均已得到证明。
(二)液泡膜转运蛋白分子生物学原理
(三)细胞质溶质的积累
(四)植物对元素的吸收及运输选择性改变
(五)自由基清除系统活性增加
在一定范围盐胁迫下,作物内源SOD活性增加,MDA含量下降,质膜透性下降,外源自由基清除剂可降低MDA含量,而SOD抑制剂预处理则提高MDA含量(赵可夫等,1993)。可见,SOD等自由基清除系统对保护膜结构,提高作物耐盐性有一定作用。
根据武德(2007)报道:盐胁迫下植物最直观的影响就是生长受到抑制,此外光合下降(吴永波,2002)、耗能增加、酶活性改变、膜透性增加、衰老加速(马焕成等,1998)。
1.盐胁迫使植物生长受到抑制
盐分对树木最普遍、最显著的效应是抑制生长,影响种子出苗率、成苗率、苗木生长量及嫁接成活率(唐伟英等,1988)。Yeo(1991)发现盐胁迫下植物的生长受阻早于光合下降。一些长期试验的结果表明生长的减少量大于光合的下降量。因此,许多研究者认为盐胁迫下造成生长下降的原因是叶面积减少造成光合面积减少,导致同化产物的供应不足,而非光合速率下降所致。盐分对根的影响超过对枝干的影响(翁迈东,1980)。盐分胁迫下一年生沙枣幼苗鲜重、干重、叶面积随外界盐分浓度的增加而减少,这种压制生长的途径可能是由于盐胁迫增加了根系下表皮的木栓化作用,从而抑制根对水分和无机离子的吸收。当植物被转移到盐逆境中几分钟后,生长速率即有所下降,其下降程度与根际渗透压成正比。Kulieva(1975)发现盐胁迫通过影响有丝分裂,减少细胞分化而减少生长。但NaCl胁迫下外加一定浓度的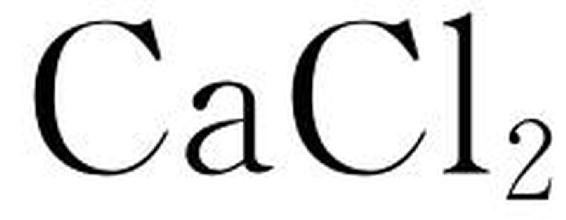 可以缓解盐胁迫对生长的抑制,说明这种抑制作用既有渗透效应也有离子效应。
可以缓解盐胁迫对生长的抑制,说明这种抑制作用既有渗透效应也有离子效应。
2.盐胁迫下膜受到伤害
盐胁迫对植物的伤害作用,在很大程度上是通过破坏生物膜的生理功能引起的。植物的细胞膜系统是盐害的原初部位和主要部位(冯利波,2005)。在盐胁迫条件下,植物一旦脱水,便会造成膜结构的破坏,导致质膜半透性的改变。另外,由于Na竞争性地取代了质膜上的Ca,也会使膜结构改变,稳定性下降,质膜的透性增强,质膜的透性越大,植物体所受到的伤害程度也就越大(夏尚光,2005)。
3.活性氧伤害和酶活性的变化
盐分胁迫下,植物通过调节其体内酶活性来适应盐胁迫对其造成的伤害。过氧化物酶(POD)、超氧化物歧化酶(SOD)的主要功能是通过消除盐分胁迫诱导产生的细胞内活性氧,抑制膜内不饱和脂肪酸的过氧化作用,维持细胞质膜的稳定性和完整性,提高植物对盐分胁迫的适应性。SOD是生物体内重要的保护酶类之一,可有效清除生物体内的氧自由基。盐胁迫条件下植物体内自由基的产生与清除之间的平衡被打破,自由基累积量增加,积累的活性氧可直接攻击膜系统中不饱和脂肪酸,导致膜脂过氧化的发生,膜脂过氧化产物MDA含量增加,膜脂组分发生变化。盐胁迫下植物体内主要的清除酶类(SOD、POD、CAT)及主要清除类物质(Vc、谷胱甘肽)的含量会明显下降,膜脂组分的变化破坏了膜的结构,使膜的流动性下降,生物膜的结构和功能遭到破坏或损伤,进而引起一系列生理生化功能的改变。
4.盐胁迫下植物体内激素的变化
根据报道发现脱落酸(ABA)、细胞分裂素(CTK)和乙烯(ET)与植物的抗盐性有很大关系。赵可夫(1998)研究表明,植物生长在盐渍条件下,植物根系的ABA合成速度大增,并且将合成的ABA通过木质部液流迅速运输到地上部分。近年来又发现利用一定浓度的ABA处理植物可以提高其抗盐能力。另外,刘桂丰(1998)研究表明,采用不同浓度NaCl处理苗木,发现苗木叶片的脱落酸(ABA)、细胞分裂素(CTK)、吲哚乙酸(IAA)、赤霉素(GA)4种激素的含量变化有较大差异。
5.盐胁迫对光合作用的影响
光合作用为植物生长提供能量,是植物生命维持的重要生理过程,也是植物生长发育的基础。众多研究(吴永波等,2002)表明,盐胁迫下叶片净光合速率和暗呼吸减弱,蒸腾作用和气孔导度下降,胞间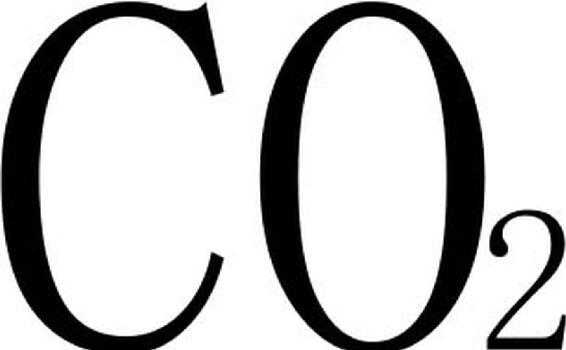 浓度增加。Luangjame(1990)通过分析桉树的耐盐性发现,随着盐分浓度的提高,光合作用降低,补偿点增加,蒸腾作用、光呼吸和暗呼吸在低浓度时增加,高浓度时降低。国槐受盐胁迫光合作用的变化规律与大多数树种不同,它的净光合速率是增加的,这可能与不同的植物进行不同的渗透调节方式有关(张川红,2002)。目前,关于盐胁迫下植物光合作用下降的原因还没有形成统一的认识(郭书奎,1999)。盐分胁迫对光合作用的影响也是通过气孔因素和非气孔因素来实现的,一般情况下,在低盐浓度时气孔因素占主导作用,而在高盐浓度时非气孔因素起主要作用。
浓度增加。Luangjame(1990)通过分析桉树的耐盐性发现,随着盐分浓度的提高,光合作用降低,补偿点增加,蒸腾作用、光呼吸和暗呼吸在低浓度时增加,高浓度时降低。国槐受盐胁迫光合作用的变化规律与大多数树种不同,它的净光合速率是增加的,这可能与不同的植物进行不同的渗透调节方式有关(张川红,2002)。目前,关于盐胁迫下植物光合作用下降的原因还没有形成统一的认识(郭书奎,1999)。盐分胁迫对光合作用的影响也是通过气孔因素和非气孔因素来实现的,一般情况下,在低盐浓度时气孔因素占主导作用,而在高盐浓度时非气孔因素起主要作用。
6.盐胁迫对植物物质代谢和渗透物质积累的影响
盐分胁迫下植物生成一些渗透调节物质,并积累一些大分子物质,淀粉蛋白质分解成小分子物质如糖、氨基酸。这些小分子物质有较强的亲水性,可以稳定胶体性质,在组织代谢中使植物细胞免受伤害或使伤害减轻。在渗透调节作用中脯氨酸(Pro)具有举足轻重的地位。除脯氨酸外,甜菜碱及多胺类化合物的合成和积累也存在于一些植物体内,在逆境环境下两者都有重要作用(刘家尧,1994;殷晓军,2002)。
7.盐胁迫引起离子失调与单盐毒害植物生长
(一)细胞膜透性
生物膜系统是细胞骨架之一,膜的稳定和流动性是细胞及整个生命活动赖以生存的基础。生物膜作为细胞和细胞器生命活动与环境的一个界面,任何一个刺激首先为细胞膜所接受,然后传导放大一系列过程,最后才由细胞膜和细胞器作出反应,并且在逆境过程中植物膜最先受到伤害。在胁迫下,植物细胞的质膜透性增加。耐盐性较强的植物细胞稳定性较强,质膜透性增加较少,伤害率低而耐盐性弱的植物则相反。盐分通过破坏植物细胞膜的选择性和细胞透性平衡,影响各种生理代谢,最经常影响植物生长发育,一般耐盐树木细胞质膜透性小,而不耐盐树种细胞质膜透性大。
(二)保护酶类活性
超氧化物歧化酶SOD的生理功能主要是清除超氧自由基,在一定范围内,随盐浓度的增加SOD活性增加,但浓度过高会降低SOD活性。增加的酶活性与耐盐性直接相关。过氧化物酶POD,POD参与多种生理代谢,催化多种细胞壁结构合成,控制细胞生长发育,清除活性氧POD随盐浓度增加,活性升高。CAT(过氧化氢酶)其活力在低盐浓度时被破坏。这些酶抑制细胞膜脂中丙二醛(MDA)的积累,维持细胞膜的稳定性和完整性,提高树木对盐分胁迫的适应性。这些酶活性高低及MDA的含量在一定程度上反映树木的耐盐性。MDA是膜脂过氧化作用的主要产物,其含量高低和细胞质膜透性变化是反映细胞膜脂过氧化作用强弱和质膜破坏程度的重要指标。一般耐盐植物MDA含量明显低于敏盐植物。
(三)脯氨酸含量
树木在盐胁迫下都发生脯氨酸积累且有利于树木耐盐性提高。当植物遭受浓度逆境时,植物体内积累脯氨酸,无盐条件下脯氨酸低,而当盐浓度增加时脯氨酸的含量升高。但在逆境条件下,植物体内脯氨酸是否作为树木耐盐指标尚没有一致意见,因为脯氨酸累积既可能是适应性又可能是细胞结构和功能受损的表现。但根据我们对树木耐盐性测定,认为它可以作为树木耐盐性指标。
(四)光合作用及叶绿素含量
盐胁迫影响下植物光合作用、净光合作用速率明显降低,叶绿素含量减少,叶绿体对盐分最敏感。盐胁迫对叶绿素的形成起到破坏作用,叶绿素是重要的光合作用物质,盐胁迫对叶绿素合成与分解之间的平衡产生影响,进而影响植物光合作用强度。对耐盐性强的树种盐胁迫反而提高光合作用效率,它们为了生存必须将部分应用于生产、生长和竞争能力的同化物转化用于形成能量,以提供细胞内离子运输和保持细胞内离子不平衡分布之需要,耐盐力强的树种盐胁迫后净光合速率虽然没有明显下降,但其生物量却下降。光合作用和叶绿素含量变化可以作为植物耐盐性鉴定指标。
树木耐盐性是植物生理学中一个比较复杂而涉及面很广的问题,但又是一个非常实际的问题,盐碱对树木的胁迫,都能从树木各个生理过程中表现出来,适者生存,不适应被淘汰,只有通过深入细致的研究,查明造林树种耐盐特性,对环境的胁迫反应和造成的危害才能提出比较有效的防御措施,从理论和实际上对筛选树种都是非常必要的。
近来对树木耐盐性研究非常活跃,现概括如下:杨传平(1997)研究也认为细胞膜透性是研究树木抗盐性的理想指标,抗性强的树种细胞膜易被破坏,透性小,抗性差的树种细胞膜被破坏严重,透性大,同时认为目前白榆、水曲柳抗盐强,黑标3号、中绥12号杨及紫椴抗盐性中等。张川红(2002)研究盐胁迫对树生长及光合作用影响,研究结果表明,盐胁迫下中林46号杨死亡率最高,核桃次之,绒毛白蜡再次、国槐没有死亡,盐胁迫下对上述4个树种净光合速率影响结果是中林46号杨与核桃苗木的净光合速率对盐比较敏感,无论高盐胁迫还是低盐胁迫,它们净光合速率比对照明显下降,并且净光合速率下降幅度随盐浓度的增加而增大,因此认为这2个树种抗盐能力低,对盐较敏感,绒毛白蜡的特点是苗木净光合速率时盐胁迫表现出有一定的抗性,无论低盐、高盐胁迫下,它们净光合作用与对照相比都没有明显下降,并认为绒毛白蜡能忍受一段时间的盐胁迫,而国槐的净光合速率受盐胁迫后不显下降而呈上升,明显不同于中林46号杨、核桃及绒毛白蜡。国槐能在一定时间内(至少34天)一定浓度(至少200mmol·L-1NaCl)盐胁迫下维持较高的光合速率,说明具有强的抗盐性,是一种忍耐盐分的树种。
刘桂丰等(1998)研究盐逆境条件下14个树种的激素、脱落酸(ABA)、细胞分裂素(CTK)、吲杂乙酸(IAA)、赤霉素(GA)变化及抗盐性。发现不同树种的同一激素含量及同一树种的不同激素含量均有较大差异。随着盐分浓度的增加ABA的含量呈上升趋势,CTK、IAA、GA含量呈下降趋势,当盐浓度增加到一定程度,激素含量变化趋势减弱。综合考虑苗木在盐胁迫下,以苗木的激素抗盐平衡系数为选择目标,将参试的树种抗盐排序由强至弱为:白榆、樟子松、水曲柳、沙松、红松、紫椴、红皮云杉、118号杨、白桦、798601杨、兴安落叶松、中绥12号杨、日本落叶松、黑林3号杨。王志刚(2000)在野外进行12个树种耐盐性试验,主要以苗木株高、胸径、地径等测树因子为评价指标。试验表明:沙枣、沙棘、多花柽柳、细穗柽柳和蒙古柽柳耐盐性强,即便在粗放管理下也适宜作为盐碱地造林树种。
李际红(2001)研究3个杨树新无性系耐盐性,5个盐分梯度处理45d,测定各无性系处理下苗木生长率、被害状、积累、质膜透性、叶绿素含量5个指标,测定其耐盐性,认为窄冠黑青杨LUT无性系[I-69x山海关杨]耐盐力在参试的3个无性系中耐盐最强,在含盐量0.3%条件下可正常生长。
赵明范(1997)虽然是研究我国西北干旱地区次生盐碱地主要造林树种的耐盐力排序,但对海岸带树木耐盐性研究有很大的参考意义。所研究树种可以试验引种到北方泥质海岸,如沙枣、沙棘、胡杨、新疆杨、枸杞、柠条。我们研究早已引入,并且成功,赵氏试验材料是3~4年生胡杨、新疆杨、枸杞、多枝柽柳、花棒、杨柴、沙枣、沙棘、梭、杜梨、拧条共11个树种,在野外测定细胞膜透性、光合作用、灰分含量和渗透调节能力4项指标。通过测定指标分析及综合长期野外树木生长情况,认为参试树种抗盐碱土上限指标,新疆杨可以在土壤含盐量<6g/L时生长正常,杜梨<4g/L、柠条<7g/L、花棒<5g/L、沙棘<6g/L、沙枣<4g/L、枸杞<7g/L、梭<30~50g/L、胡杨<8g/L、柽柳<50g/L、杨柴<6g/L时生长正常,参试树种的耐盐碱排序由大至小为梭、柽柳、胡杨、枸杞、柠条、杨柴、沙棘、新疆杨、花棒、沙枣、杜梨。
马焕成、王沙生(1998)比较胡杨和群众杨在盐分胁迫下的生化反应发现:① 胡杨的细胞膜能忍耐200mmol NaCl的高盐处理,其膜透性在高盐处理3d后没有显著增加,群众杨的膜透性随盐分浓度的升高而升高。胡杨的SOD酶活性上升较快,随盐胁迫强度的加强而增加,群众杨受盐胁迫SOD酶活性上升较慢,且强迫下活性反而减弱,说明群众杨SOD酶活性本身的盐稳定性差,不是有效膜系统。盐胁迫蛋白电泳谱带也有相似的变化趋势:群众杨的谱带由10条到3条,胡杨则由4条增加到7条。说明胡杨在盐胁迫下出现了稳定膜结构成可兼容溶质有关的蛋白。② 胡杨有较强的代谢调节能力。盐处理后胡杨、群众杨的脯氨酸和精胺的增加水平没有显著差异,差异主要是腐胺含量:低盐7d,胡杨叶的腐胺含量是群众杨的3.4倍,高盐胁迫7d则是4.4倍。多胺别是其中的腐胺能稳定生物大分子和保护细胞膜的完整性。因此,可以认为胁迫条件下大量增加腐胺的生物合成是胡杨抗盐机理的一部分,可以作为抗盐的一种生理指标。
戴蒲英(1998)在水培条件下进行刺槐、马尾松、水杉种子和幼苗的耐盐性试验,盐浓度为0%、0.1%、0.2%、0.3%,同时用250×10-6多效唑(MET)测定预处理种子后的耐盐性,结果表明:盐浓度越大,种子发芽势及发芽率越低,简化苗木活力指数(SVIS)越小,MET对种子发芽势、发芽率及SVIS有抑制作用。但经MET处理后,盐浓度越大,种子发芽率、SVIS越大,根茎比随浓度的增加而降低,MET有增加根茎比值的趋势,幼苗体内游离脯氨酸含量随盐浓度的增加而增加,MET能提高脯氨酸的含量;在试验盐浓度下,刺槐、马尾松耐盐能力为0.2%~0.3%,水杉0.1%~0.2%。在沙培条件下,用250×10-6硼酸、0.3%氯化钙和250×10-6MET处理刺槐、湿地松、侧柏种子,其耐盐能力增强,其中,氯化钙处理湿地松效果较好,硼酸处理侧柏效果好,MET处理居中,3种处理对刺槐的耐盐能力影响不大。
吴永波等(2002)通过盆栽实验研究盐胁迫对大叶白蜡、小叶白蜡、绒毛白蜡幼苗生长与光合作用的影响,盐分处理为0.1%、0.2%、0.3%、0.4%、0.5%、0.6%6个水平,测定幼苗光合速率、叶绿素和丙二醛含量和幼苗生物量。研究表明,随着NaCl处理浓度的增加,3个白蜡树种幼苗的总生物量和叶片的叶绿素含量、净光合速率、气孔导度和蒸腾速率均呈下降趋势;而饱和水气压差呈上升趋势,幼苗叶片的胞间浓度呈上升或下降趋势。叶片和丙二醛含量并无明显规律性变化。
植物在生长过程中对盐胁迫最为敏感。因此,生物量提供了评估盐胁迫程度和植物耐盐能力的可靠标准。在盐胁迫下,叶肉导度和光合面积下降是植物生长受抑制的主要原因,而叶肉导度的下降必然导致光合速率下降。当然,在盐胁迫下,导致光合速率降低的原因有两种可能:一是由于气孔因素引起的,二是由于非气孔因素造成的,实验中的3个树种中,大叶白蜡和小叶白蜡属于前者,而绒毛白蜡属于后者。
通过以上指标分析可以初步认为,小叶白蜡幼苗的耐盐性优于其他两种幼苗。
汪贵斌、曹福亮等(2001)通过盆栽试验,对我国南方银杏、侧柏、火炬松和刺槐4个造林树种苗木叶片中K+、na浓度和na/K+比进行了测定,盐处理水平为0(CK)、0.1%、0.3%和0.5%。随着盐浓度的提高,各树种叶片中na浓度和na/K+比呈逐步升高的趋势,但K+浓度变化较小,随着盐胁迫时间的推移,各树种叶片中Na+、K+浓度和 Na+/K+比都没有明显的变化规律,方差分析和多层比较表明,侧柏、火炬松和刺槐叶片中Na+、K+浓度和Na+/K+比在4组处理间的差异均达显著或极显著水平,4个树种中刺槐和侧柏的耐盐能力最强,银杏次之,火炬松最弱。
此外,他们采用温室盆栽试验方法,研究了不同土壤水分下盐胁迫对1年落羽杉实生苗的生长及营养吸收的影响。试验共12种处理,3种水分水平(W1,淹水;W2、W3土壤含水量分别为田间持水量的75%和25%),每种水分下设4种盐分处理(NaCl含量为土壤干重的0%、0.15%、0.3%、0.45%),处理时间为130d,结果表明:土壤盐分胁迫下落羽杉幼苗的生长及对营养的吸收发生了显著的变化,这种变化又因土壤含水量的不同而不同。落羽杉相对高生长、相对地径生长和生物量增量随着土壤盐分浓度的增加和土壤水分含量的减少而减少。在盐水条件下,随着土壤盐分浓度的增加,落羽杉幼苗根、茎、叶全N、全P、全Na,以及叶片中的全Ca、全Fe的含量均随着升高,而根、茎中的全Fe、全Ca、全Mg含量差异较小。在W2水分条件下,各种盐分处理都在不同程度上增加了根、茎、叶中全N、全Na、全Ca和全Fe含量,茎和叶中全P、全K和全Mg含量,而根系中的全P、全K和全Mg含量在盐分浓度较高时有所下降。在W3水分条件下,各种土壤盐分不同程度地增加了叶片全N、全P、全K含量,茎和叶中全Mg和全Ca含量,以及根、茎、叶中全Na含量,而茎和根中全N、全P、全K含量,根、茎和叶中全Fe含量均随着土壤盐分浓度的增加而下降。
总之,在盐胁迫下,木麻黄幼苗SOD活性下降,CAT、POD活性及蛋白质含量升高,适宜的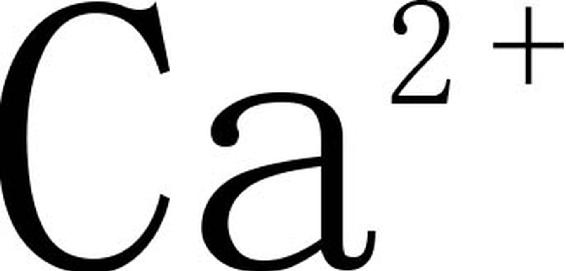 可增加木麻黄幼苗对盐胁迫的耐受能力,调节抗氧化酶活性并使之相对稳定,从而提高植物的抗逆性。根据本文结果,提高木麻黄幼苗抗盐力的最适Ca(NO3)2浓度为48g/kg土。
可增加木麻黄幼苗对盐胁迫的耐受能力,调节抗氧化酶活性并使之相对稳定,从而提高植物的抗逆性。根据本文结果,提高木麻黄幼苗抗盐力的最适Ca(NO3)2浓度为48g/kg土。
近年来,研究表明,钙能提高植物组织和细胞多种抗性,包括抗盐性。
徐呈祥等(2005)以2年生金丝小枣嫁接苗为试材,研究了盆栽条件下硅对受胁迫金丝小枣生长和生理效应。结果表明,金丝小枣在NaCl 3.0g/kg和5.0g/kg土壤中长期生长,给土壤施硅(0.4g/kg)较不施硅处理的植株叶片相对含水量(RWC)分别提高7.30%和10.05%,叶片细胞质膜相对透性(PMP)分别降低234.60%和108.13%,光系统II(PSII)的潜在光化学活性(Fv/Fo)和最大光化学效率(Fv/Fm)分别提高32.25%、30.71%和11.26%、12.15%,同化速率(Pn)分别提高10.31%和21.13%,枣头枝长度和叶干重分别增加87.40%、36.73%和238.16%、40.38%。这说明盐胁迫下,给土壤施硅可改善金丝小枣树的生长和生理状况。
李国旗、张纪林等(2003)对生长在苏北沿海防护林不同土壤盐度下的主栽树种杨树(欧美杨无性系69杨)进行了解剖,描述了杨树次生木质部结构的特征,测量了次生木质部的16个数量特征:年轮宽度(ARW)、导管长度(VL)、早材和晚材的导管分子面积(VA)、导管周长(VP)、导管直径(VD)、导管单孔率(PSV)、导管频率(VF)、纤维长度(FL)、纤维宽度(FW)、射线高度(RH)和射线频率(RF)。实验结果表明,在不同土壤盐度胁迫下:① 杨树次生木质部形态特征的差别不明显,但是组成分子趋向于小型化。② 次生木质部数量特征的变化比较明显,即当土壤含盐量从0.036%逐渐升高到0.289%,杨树的年轮宽度变小,即0.91mm→0.77mm;纤维长度和宽度的变化分别为693.8μm→570.4μm和14.9μm→13.5μm,射线高度和频率的变化分别为284.3μm→249.2μm和58.9mm-2→75.5mm-2,表明在盐胁迫下杨树的生长受到抑制,随着盐度的增加(0.036%→0.289%),杨树导管长度减小,367.6→341.5μm;在早材和晚材中,导管面积变化分别为1575.6μm2→1703.6μm2和 760.1μm2→947.7μm2,导管周长变化分别为167.7μm→195.1μm和120.8μm→143.7μm,导管直径的变化分别为41.8μm→43.4μm和29.1μm→33.1μm,导管频率的变化分别为141.8mm -2→144.2mm-2和160mm -2→206.7mm -2;导管单孔率的变化分别为65.9%→30.5%和59.4→40.8%。这些都表明在盐胁迫下杨树的导管缩短变粗,呈聚集分布,输水效率和输水安全性都有所提高。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











