



五、以脂肪细胞发育分化为例介绍真核转录调控的研究
本实验室开展脂肪细胞的分化调控工作,在工作中应用多种手段来研究转录调控的机制。下面就以脂肪细胞的分化过程为例介绍基因转录调控以及相关的研究方法。
脂肪细胞在分化过程中获得形态学表型(例如脂肪细胞内脂滴的聚集)的同时,在分子水平上也发生了各种基因的表达变化。根据对分化前、分化中和分化后的细胞抽提液的双向电泳分析结果,发现在开始分化的5h内,至少有100种以上的蛋白质表达情况发生了变化,其中就包含了重要的转录激活因子。在分化过程中有利于脂肪细胞分化或脂肪细胞所特有的蛋白质组分会随分化过程增加,抑制脂肪细胞分化或对脂肪细胞的生理功能不必要的基因表达受到抑制。无数基因的这种有严格时序的变化,大部分都发生在转录水平上。
近年来,对脂肪细胞分化的调控程序的研究,取得了重大的进展。与此调控程序相关的反式激活因子已获得鉴定,它们相互协调而顺序地引发前脂肪细胞分化为成熟脂肪细胞,脂肪细胞的分化是由几个转录因子协同调节的,其中最主要的是C/EBP(CAAT/enhancer binding protein)家族和PPAR(peroxisome proliferator activated receptor)家族的转录调控因子。在分化过程中,它们各自成员的表达,也都遵循着严格的时序性,从而协调作用完成前脂肪细胞向成熟脂肪细胞的转变。C/EBPβ和δ在脂肪细胞分化早期被诱导产生,并且是诱导PPARγ和C/EBPα产生的主要转录调控因子。PPARγ和C/EBPα共同诱导一系列脂肪细胞特异表达的基因,从而使细胞得以最终分化。转基因实验表明,PPARγ和C/EBPα可以把非脂肪系细胞诱导成为脂肪细胞。其他一些转录调控因子也在脂肪细胞的分化过程中发生一定的变化,如ADD1/SREBP1,但它们在诱导脂肪细胞分化中的功能还有待进一步研究。

图2-11 C/EBP家族与C/EBP类似蛋白通过亮氨酸拉链形成异二聚体
(一)C/EBP家族
与脂肪细胞分化相关的3个成员为C/EBPα、C/EBPβ和C/EBPδ。它们的羧基端的碱性氨基酸区/亮氨酸拉链区,含有55~65个氨基酸,具有与DNA结合以及形成二聚体的能力。它们可以形成同二聚体,也可与别的蛋白形成异二聚体(如图2-11)。与C/EBP反式激活因子相结合的顺式作用元件通常是ATTGCGCAAT。它们的N端具有转录激活和转录抑制作用。在脂肪细胞分化过程中,C/EBPβ和C/EBPδ在分化早期会暂时性升高,而C/EBPα的表达较迟,在诱导后36h才开始表达。
1.C/EBPα 它能与多种脂肪细胞特有基因的启动子相结合而激活它们的转录过程(如胰岛素受体)。它的基因表达刚好在大多数脂肪细胞所特有基因的转录启动之前发生。有证据表明C/EBPα在脂肪细胞分化的转录激活过程中起着十分重要的作用。
第一个C/EBP家族的转录因子C/EBPα首先是在肝细胞内发现的,后来发现它分布于许多不同的组织中。它的DNA结合位点存在于许多基因的调控区域内并具有抑制细胞有丝分裂的作用。以后又陆续发现了C/EBP家族的其他成员。
C/EBPα是第一个被证明在脂肪细胞分化过程中起重要作用的转录因子。它属于含碱性区亮氨酸拉链(basic region/leucine zipper,bzip)类蛋白。C/EBP家族成员的分子结构中含有1个可调节与DNA结合的碱性区(basic region domain)以及1个可以和另1个C/EBP分子形成同源二聚体的亮氨酸拉链模体(leucine zipper motif)。
C/EBPα在3T3-L1细胞的分化过程中本身是被分化诱导表达的基因。在3T3-L1前脂肪细胞内,C/EBPα基因是不表达的。当细胞在分化诱导激素的作用下,C/EBPβ和C/EBPδ首先开始表达并且诱导C/EBPα基因开始表达,C/EBPα的表达具有自身诱导作用,即在C/EBPα基因的调控区内有一个它自己的结合位点,一旦C/EBPα基因开始表达后,产生的转录因子就能不断地与自身基因调控区的位点结合,并进一步诱导自身的表达,从而产生大量的C/EBPα转录因子诱导细胞的分化(图2-12)。
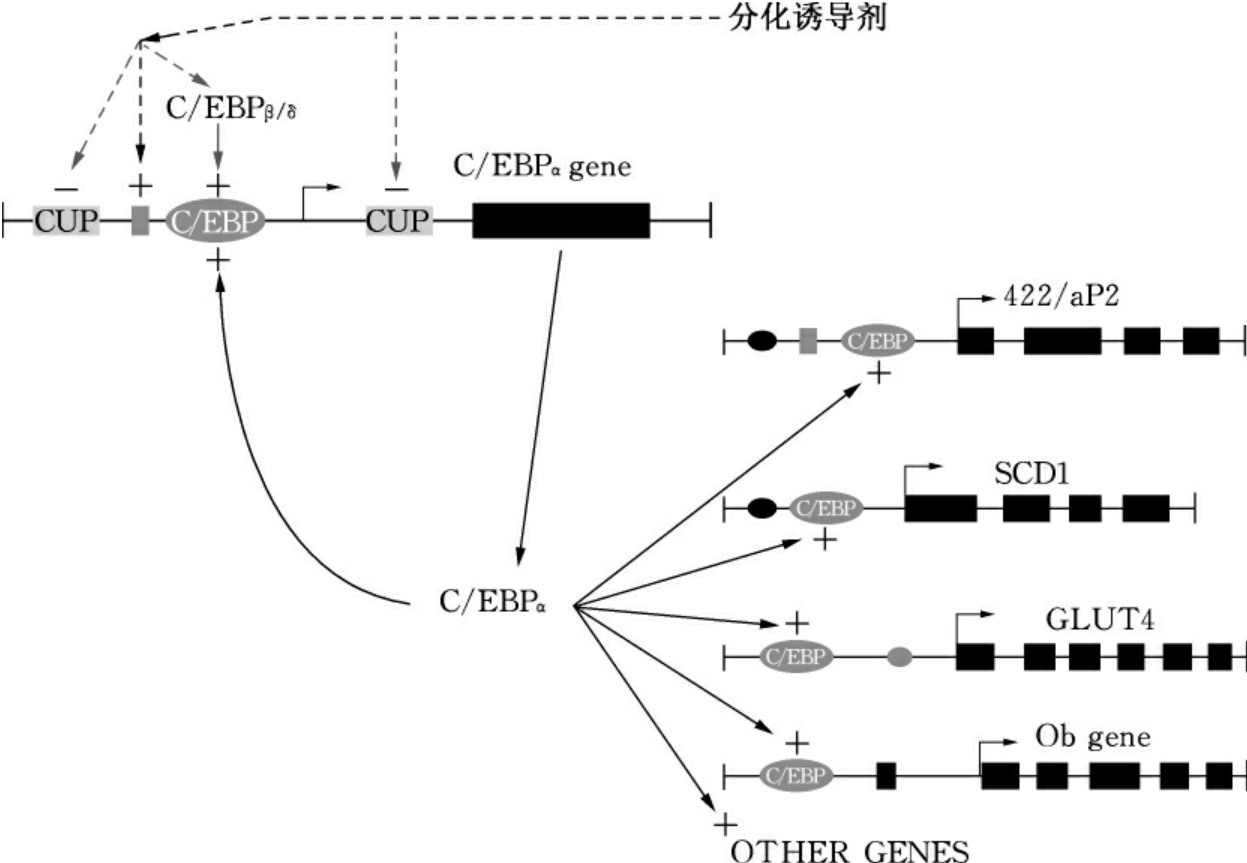
图2-12 C/EBP家族在脂肪细胞分化过程中的转录调控作用
对脂肪细胞特别是3T3-L1细胞的研究表明,C/EBPα在脂肪细胞分化过程中起着重要作用。虽然C/EBPα的表达不是脂肪细胞专一性的,但C/EBPα能启动脂肪细胞特异性表达基因的转录。在脂肪细胞特异表达的基因中,如aP2基因、十八烷酰辅酶A去饱和酶(SCD)1基因、葡萄糖转运体(GLUT)-4基因、磷酸烯醇丙酮酸羧激酶(PEPCK)基因、瘦蛋白(leptin)基因和胰岛素受体基因的调控区域内,都含有C/EBPα的结合位点。并且体内外的实验都证明,C/EBPα不但能和它的位点结合,而且结合后能启动基因的表达。进一步的实验证明,使C/EBPα的位点突变后,这些基因的调控区就不能诱导报告基因的表达。所以C/EBPα不但是调控这些基因表达的转录因子,而且是启动基因表达的主要因素。在3T3-L1前脂肪细胞中,反义C/EBPαmRNA片段可以抑制3T3-L1细胞的诱导分化。在表达反义C/EBPαmRNA片段的3T3-L1细胞中,不但激素诱导的细胞分化被抑制,而且激素诱导的脂肪细胞特异表达基因的表达也被抑制。如果利用转基因使3T3-L1细胞在没有激素诱导的情况下表达C/EBPα,则细胞可以在没有诱导激素的情况下分化成为脂肪细胞。这主要是因为转基因的C/EBPα的表达诱导了分化基因的表达,使细胞进入分化程序。在不表达C/EBPα基因的非脂肪系成纤维细胞中,通过转入外源C/EBPα基因并使它在细胞内表达,结果这些成纤维细胞分化成了脂肪细胞。所以C/EBPα可以诱导非脂肪细胞系的细胞分化成为脂肪细胞。由于C/EBPα在脂肪细胞分化过程中的表达比较晚,所以它和脂肪细胞的分化有关,但和分化前的细胞定向没有太大关系。
2.C/EBPβ和C/EBPδ 对C/EBP家族成员在脂肪细胞分化中的作用目前的了解还比较少。但在诱导3T3-L1细胞分化的过程中,C/EBPβ和C/EBPδ的表达远比C/EBPα要早,并且它们只有在分化诱导的早期有短暂的表达增加。由于它们在表达时序性上较其他的转录因子早,所以一般认为它们和启动其他对分化重要的转录因子有关(图2-13)。
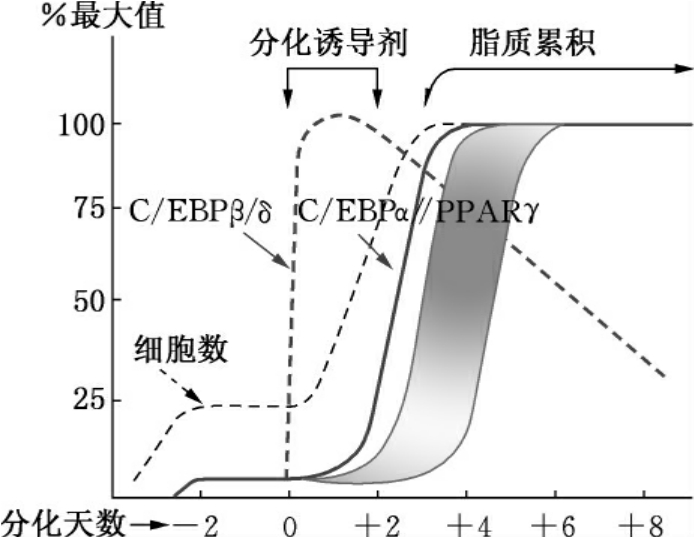
图2-13 3T3-L1前脂肪细胞分化过程中各转录因子表达上的时序差异
在脂肪细胞分化过程中,每个C/EBP家族成员都有明显的表达上的时序差异(见图2-13)。除了C/EBPα的表达发生在分化的相对后期外,C/EBPβ和C/EBPδ少量存在于前脂肪细胞中,并在分化早期它们的表达出现短暂的增加。但在分化后期,C/EBPβ的表达量降低至50%左右,而C/EBPδ则几乎无法检测到。由于C/EBP家族的转录因子均不是脂肪细胞专一性的,它们一般是在脂肪细胞分化中起作用,而不是分化前的细胞命运的决定者。
在脂肪细胞分化早期,C/EBPβ被诱导表达,大约在4h达到最高水平。在脂肪细胞分化过程中,C/EBPβ被认为是C/EBPα的转录激活因子,它与C/EBPα启动子上的C/EBP结合位点结合后激活C/EBPα的表达。C/EBPβ表达初期没有DNA结合活性,因而不具有转录激活功能。当本来处于静止状态的前脂肪细胞重新进入细胞周期后,C/EBPβ才获得DNA结合活性。C/EBPβ除具有激活C/EBPα的功能外,还参与调节前脂肪细胞重新进入细胞周期(mitotic clonal expansion)——与脂肪细胞分化形成同步的一个过程。卫星DNA上存在C/EBP结合位点,C/EBPβ得到DNA结合活性后能够结合到卫星DNA上,通过免疫荧光染色证实,C/EBPβ在细胞周期各个阶段都和染色体DNA结合在一起。因此C/EBPβ在脂肪细胞分化过程中具有两方面功能:一是调节细胞分裂;二是激活C/EBPα,诱导脂肪细胞成熟分化。
C/EBPβ在前脂肪细胞诱导分化的早期不具有DNA结合活性的原因是由以下两种原因造成的。原因之一是在C/EBP表达早期,C/EBPβ和CHOP-10(CHOP-10是C/EBP类似蛋白,C/EBP homologous protein)形成异二聚体。CHOP-10可以与C/EBPβ的C端结合形成二聚体,从而使C/EBPβ缺乏和DNA相结合的碱性氨基酸区域,因此C/EBPβ和CHOP-10形成的二聚体不能和DNA结合。当前脂肪细胞受诱导剂诱导时,CHOP-10表达受抑制。CHOP-10的表达下降和C/EBPβ得到DNA结合活性时相上一致。此外,蛋白酶抑制剂ALLN可诱导CHOP-10表达,抑制C/EBPβ的激活,从而抑制脂肪细胞的分化。另一个原因是C/EBPβ在脂肪细胞分化过程中还受到磷酸化调节,通过二维电泳凝胶电泳结合Western证实,C/EBPβ初表达时处于相对低磷酸化状态,前脂肪细胞重新进入细胞周期后则处于高磷酸化状态。从低磷酸化到高磷酸化的过程和C/EBPβ得到DNA结合活性相一致。
(二)PPAR家族
PPAR属于核受体类转录因子,起先因能被配基激活并诱导过氧化酶体增殖而被鉴定。在过去几年中,大量研究证明,PPAR在脂肪细胞分化过程中起着重要作用。PPAR的激活不仅能促进前脂肪细胞的分化,而且也能促进非脂肪细胞系细胞分化成脂肪细胞。在调控脂肪细胞基因表达方面,PPAR通过PPAR-RXR(retinoid-X receptor)二聚体与一个相应元件结合而调控基因的转录,该元件是由间隔一个核苷酸的PuGGTCA的重复序列组成,它已在一些脂肪细胞特异表达基因的调控区中被鉴定。
PPARγ是PPAR家族中最具脂肪细胞专一性的成员,而且它在脂肪细胞分化诱导过程中是在大多数脂肪细胞特异基因的表达之前被诱导的。它有2个N-末端不同的异构体PPARγ1和PPARγ2,属于同一基因选择性地剪切的结果,两者在脂肪组织内均表达丰富。但PPARγ2基本上是脂肪组织中专一性表达的,而PPARγ1则在各种组织中均能低水平表达。到目前为止,还没有发现两者有功能上的不同。在典型生长的成纤维细胞系中,PPARγ的表达足以诱导细胞生长抑制并启动细胞分化成脂肪细胞。但目前对PPARγ在脂肪细胞系的决定过程中怎样作用仍不清楚。
(三)C/EBPα和PPARγ在脂肪细胞分化过程中的协同作用

图2-14 转录因子对脂肪细胞基因表达以及细胞分化的调控
在脂肪细胞诱导分化过程中,C/EBPα和PPARγ相互协调作用,促进脂肪细胞基因的转录与表达。如图2-14所示,C/EBPβ和C/EBPδ在激素诱导后迅速产生,而C/EBPβ的产生能激活PPARγ和C/EBPα的产生。在C/EBPα促进PPARγ产生的过程中,C/EBPβ极有可能是初始的诱发剂,因为C/EBPβ在细胞分化过程中的产生在C/EBPα之前,而且C/EBPα基因的调控区含有C/EBPβ结合位点,C/EBPβ因而可通过此结合位点激活C/EBPα基因的表达。一旦C/EBPα基因的转录被启动,C/EBPα则可通过自身的转录激活而继续高水平表达,从而维持脂肪细胞基因的表达并最终促使细胞分化。
C/EBPα和PPARγ能相互激活转录并具有协同作用,触发脂肪细胞的最终分化。在NIH3T3成纤维细胞中,C/EBPα和PPARγ单独的转基因表达仅是促使细胞部分分化成脂肪细胞,而两者同时表达则促使了细胞完全分化成脂肪细胞。而对成肌细胞的转基因实验表明,C/EBPα和PPARγ单独的作用都不能诱导成肌细胞分化成为脂肪细胞。但两者的基因同时转入成肌细胞则能使细胞自发分化成为脂肪细胞。
在脂肪细胞的分化过程中,基因表达的调控主要是由C/EBP家族和PPAR家族的转录调控因子协调作用的。而在这两个家族的成员中,C/EBPα和PPARγ是两个最重要的转录调控因子。
(李 希 汤其群)
1.宋后燕,朱运松主编.医用分子遗传学.上海:复旦大学出版社,2003
2.孙乃恩,孙东旭,朱德煦.分子遗传学.南京:南京大学出版社,1996
3.Robert F.Weaver.Molecular biology.McGraw-Hill Companies,Inc,1999
4.Lodish H,Berk A,Zipursky S.L,et al.Molecular cell biology.4th ed.New York:W.H.Freeman and Company,2000
5.Gregoire FM,Smas CM,Sul HS.Understanding adipocyte differentiation.Physiol Rev,1998,78:783
6.Macdaugald OA,Lane MD.Transcriptional regulation of gene expression during adipocyte differentiation.Annu Rev Biochem,1995,64:345
7.Rosen ED,Sarraf P,Troy AE,et al.PPARγis required for the differentiation of adipose tissue in vitro and in vivo.Mol Cell,1999,4:611
8.Tang Qi-Qun,Otto TC,Lane MD.Commitment of C3H10T1/2pluripotent stem cells to the adipocyte lin-eage.Proc Natl Acad Sci USA,2004,101(26):9607
9.Tang Qi-Qun,Tamara CO,Lane MD.Mitotic clonal expansion:A synchronous process required for adipo-genesis.Proc Natl Acad Sci USA,2003,100:44
10.Tang Qi-Qun,Lane MD.Role of C/EBP homologous protein(CHOP-10)in the programmed activation of CCAAT/enhancer-binding protein-βduring adipogenesis.Proc Natl Acad Sci USA,2000,97:12446
11.Lane MD,Qi-Qun Tang,Jiang MS.Role of the CCAAT enhancer binding proteins(C/EBPs)in adipocyte differentiation.Biochem Biophys Res Comnmun,1999,266(3):677
12.Tang Qi-Qun,Lane MD.Activation and centromeric localization of C/EBP’S during the mitotic clonal ex-pansion of adipocyte differentiation.Genes Development,1999,13(17):2231
13.Tang Qi-Qun,Liang MS,Lane MD.Repressive effect of Sp1on the C/EBPαGene promoter:Role in adi-pocyte.Mol Cell Biol,1999,19(7):4855
14.Lane MD,Tang Qi-Qun,Jiang MS.The adipocyte differentiation program:Repression/derepression of C/EBPαgene.J Animal Science,1999,77,(Suppl.3),23
15.Lane MD,Jiang MS,Tang Qi-Qun.Role of C/EBPαin adipocyte differentiation.In:Bray G and Ryan D,Proceding of penningtion symposium on nutrition genetics and obesity.Ed.by LSU Press,Balton Rouge,LA.1998,459
16.Tang Qi-Qun,Jiang MS,Lane MD.Repression of transcription mediated by dural elements in CCAAT/enhancer binding protein a gene.Proc Nalt Acad Sci USA,1997,94:13571
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











