



第二节 生命的进化
对生命的产生有了初步了解后,我们对周围生命世界的探索并未停止,对复杂多样的生物世界形成的原因和机制即生物进化的过程的探索,将极大地帮助我们理解这个纷繁芜杂的世界。进化论是生物科学的核心理论,也是生物科学理论中最富生命力的理论之一。生物进化简单来说是指生物种群在一定时间内性状和遗传组成上的变化,是生物与其生存环境相互作用的过程中,其遗传系统随时间而发生的一系列不可逆改变后,导致相应的表型的改变。
现代生物进化理论一般从微观和宏观两个角度来探讨生物进化的过程和机理,即微观进化(microevolution)和宏观进化(macroevolution)。微观进化,也称小进化,是指种内的个体和种群层次上的进化改变;宏观进化,也称大进化,指种和种以上分类群的进化。微观进化是无性繁殖系或种群在遗传组成上的微小差异导致的微小变化,是进化的基础。多种微观进化汇集的结果即表现为宏观进化。
一、生物的微观进化
生物微观进化的基本单位是无性繁殖系和种群。对于原核生物和无性繁殖的真核生物而言,在不发生突变的情况下,同一克隆的个体之间无遗传差异,差异存在于不同的克隆之间;有性生殖的生物,任何一个个体都不可能把自己的基因型全部遗传给下一代,故由个体构成的种群之间必然存在差异。因此,个体不是进化的基本单位,无性繁殖生物的进化基本单位是无性繁殖系,有性繁殖生物的进化基本单位是种群。
1.分子水平的生物进化
分子水平的生物进化是以生物大分子为基础,从分子水平上研究生物进化,主要是研究核酸和蛋白质等生物大分子结构和功能的变化。从时序上来讲,分子进化是生物进化的初始阶段,是生物组织基础层次的进化,只有从分子水平研究生物的进化才能触及生物进化的本质。
(1)核酸的进化
生物进化过程中,总趋向于结构复杂、功能完善方向发展。复杂结构、功能完善的生物要维持生命并繁衍,需具备大量的基因。在整个进化过程中,从低等生物到高等生物,细胞内的DNA含量逐渐增加(图7-8)。很多基因只有高等生物才有,如血红蛋白基因、免疫球蛋白基因、结合珠蛋白基因等。原核细胞的遗传信息一般是由几个至几千个基因组成,而哺乳动物物种的一个单倍体基因组约有3×109个碱基对,约相当于300万个基因。

图7-8 不同生物细胞中DNA的含量
一般来讲,越是高等的生物,DNA含量就越高。果蝇的卫星DNA和一些啮齿类动物的对比研究表明,DNA的量是以每100万年至少1%的速度递增。但DNA含量不一定总是跟生物的复杂程度成正比,有些生物虽然在分类位置上并不高等,但其DNA含量却相当高,如一种肺鱼的单倍体基因组DNA含量几乎是哺乳动物的40倍,甚至很多两栖动物和一些植物的DNA含量都高于哺乳动物。有些亲缘关系比较近的物种之间,DNA含量也差异比较大。如蚕豆(Vicia faba)的DNA含量是大巢菜(Ricia sativa)的7倍多。这些现象产生的原因都是由于真核基因组内有很多重复的DNA引起的。在生物进化过程中,真核生物DNA有很多是靠重复获得的。
生物进化不仅带来DNA含量的变化,还引起DNA质的变化,也就是核苷酸顺序的变化。这主要是由核苷酸链中碱基的替换、插入或缺失等原因引起的。这种在遗传物质的复制和翻译过程中发生的差错,引起个体的突变,是生物进化的主要驱动因素之一。也是由于这种变化,通过不同生物的DNA序列的差异可以反映它们之间的亲缘关系的远近,亲缘关系越近的两个物种的核苷酸序列差异越小,反之亦然。比如人与猕猴的核苷酸差异比例为9.3%,而与黑猩猩的差异比例为2.5%,人与黑猩猩的亲缘关系更近,与猕猴的关系更远。
基因的进化更是一个十分复杂的生物化学过程,如编码区与非编码区的进化、启动子的进化等。以内含子的起源与进化为例,可以更好地理解基因的起源和进化。内含子是在20世纪70年代被发现,而对于其起源仍没定论,目前主要有两类观点:“后起源论”(intron-late)和“先起源论”(intron-early)。后起源论认为内含子都是作为间隔序列插入到连续编码的基因序列中而形成的,内含子在较高级的功能基因或真核生物出现后才产生。先起源论认为早期的内含子具有自我催化、自我复制等能力,它们是原始基因和基因组的组织与复制所必不可少的。目前支持先起源论的证据很多,但也不能完全排除内含子的后起源产生方式。
基因新功能获得可以通过突变、产生重叠基因、选择性剪接和基因共享等途径来实现。重叠基因是在同一条DNA片段上有不同的可读框构成的有相互重叠的基因,这种情况在病毒、细菌和细胞器中较常见。基因共享即基因及其产物在保持原有功能的情况下新获得一种或多种功能,简而言之,是一基因多用。如眼晶体蛋白除用于维持眼睛晶体状的透光度和折光性,鸟类和鳄鱼的ε-眼晶体蛋白还具有乳酸脱氢酶活性,分析发现这两种蛋白来源于相同基因翻译的同种蛋白质。新基因的形成与基因功能的进化关系十分密切。
新基因形成的途径除突变和产生重叠基因外,还有基因重复、基因延长、外显子改组等。基因重复是一个基因中的部分区域或整个基因倍增,其中重复部分可发生突变形成新基因。基因延长是通过基因中部分区域重复等使基因长度的增加,从而使其产物活性位点和稳定性增加,进而增强原基因功能或获得新功能。内含子还能重组使外显子在不同基因之间进行交换组合,从而使外显子改组,形成杂合基因。
生物进化是种群或无性繁殖系在长时期内遗传组成上的变化,这种变化主要体现在基因频率(gene frequency)和基因型频率(genotype frequency)的改变。基因频率是指群体中某一等位基因在该位点上可能出现占基因总数的比例,基因型频率是指某种基因型的个体在群体中所占的比例。由突变、选择、迁移以及偶然因素能引起种群基因频率变化是微观进化的主要因素。如果没有这些因素,则一个有性种群的遗传组成保持相对恒定。换句话说,在种群内不发生突变、种群成员没有迁入迁出、没有自然选择作用、没有任何其他因素作用的情况下,有性生殖过程不会改变种群基因库的基因频率。这就是遗传上的哈迪-温伯格定律发生的理想条件。
(2)蛋白质的进化
蛋白质由基因编码,蛋白质组是与基因组相对应的动态实体,基因和基因组的进化也需要蛋白质参与。因此,蛋白质和蛋白质组的进化与基因和基因组的进化是密切相关的。一般从两个方面来研究蛋白质进化的一般规律:不同物种中同一蛋白质结构的比较,同一物种中不同蛋白质结构的比较。
不同物种中同一种蛋白质结构研究比较清楚的是不同生物的细胞色素C的氨基酸序列。它由一条多肽链组成,脊椎动物的细胞色素C一般有104个氨基酸残基,无脊椎动物、植物和真菌的细胞色素CN末端含有4~8个氨基酸残基。通过细胞色素C的序列对比,可以比较不同生物之间的亲缘关系,序列差异越小关系越近。如人与黑猩猩的差异为0,而与酵母菌的差异为44,说明人与黑猩猩的亲缘关系更近,而与酵母菌的亲缘关系更远。通过比较20多种生物的细胞色素C的氨基酸序列,发现有35个位点的氨基酸相同,是保守位点。这些保守位点有11个是连在一起的,这一部位是其形成正确三维结构和发挥功能所必需的。
不同蛋白质分子的进化速率不一:血纤维蛋白肽的改变氨基酸序列的10%需1.1百万年,血红蛋白改变氨基酸序列的1%需5.8万年,而组蛋白H4改变氨基酸序列1%需要600百万年。凡是对生物生存制约性大的进化速度慢,反之则进化速度快。
宏观水平上,生态关系密切的生物之间存在协进化(coevolution)。在分子水平上,协进化现象同样存在,主要发生在密切联系的相互作用的生物大分子之间。如细胞因子与其受体和配体,蛋白酶多肽抑制剂与其受体之间等。这种协同进化首先体现在序列进化速率的相关性上,而不同蛋白质分子之间的协进化,主要是因为具有相互作用的蛋白质分子之间存在功能上的相互制约。
(3)分子进化与中性学说
广义的分子进化有两层含义:一是原始生命出现之前的进化,二是原始生命产生之后在进化发展过程中,生物大分子结构变化以及这些变化与生物进化的关系。我们通常所说的是后者,也即狭义的分子进化。分子进化的速率是恒定的,因此可以为生物进化提供一个时间表。分子进化速率是通过比较不同生物同源核酸和蛋白质等生物大分子在进化过程中碱基或氨基酸发生替换的频度,来测定生物大分子进化快慢的尺度,以年为单位。如松田对8种脊椎动物血红蛋白α链进行比较研究,推断8种脊椎动物的起源时间和分歧时间的长短。
分子进化的中性理论是由日本的木村资生提出,其主要观点是分子水平上的进化大都不是通过达尔文的自然选择,而是由选择中性或近中性突变基因的随机固定实现的。所以,在分子进化过程中,突变压和随机固定起着重要作用。只有进一步导致形态和生理上的差异,自然选择才能发挥作用。
2.物种形成与进化
在进化论产生之前,物种是可依据表型特征识别和区分的基本单位。如林奈认为物种是由形态相似的个体组成,同种个体可以自由交配,能产生后代,他肯定了物种的客观性和稳定性,同时也认为物种不变。进化论产生后,对物种的概念,经历了长时间的讨论。达尔文认为一个物种可以进化为另一个物种,物种之间存在不同程度的亲缘关系。他认识到物种可变,但对于物种的稳定性没有很好的解释。现代生物学对什么是物种进行了深入的研究,以迈尔(Mayr E)的定义最为简明清楚:物种是种群所组成的生殖单元(与其他单元在生殖上隔离),它在自然界中占有一定的生境地位。我国生态学家陈世骧对这一定义进行了补充:在宗谱线上代表一定的分支。从而形成了一个简明、完整的关于物种的定义。
根据物种的定义,物种的形成包括三个步骤:谱系分裂;不同谱系间产生生殖隔离;当间隔足够大而使生殖隔离机制完善时,新物种即形成。自然选择是促成物种形成的主要因素,如Dodd将培养在同一培养基下的果蝇(Drosophila pseudoobscura)分成两个种群,分别放入仅装有麦芽糖和淀粉的培养基中进行培养。经过8代后,取两个种群中不同性别的果蝇进行交配实验,发现实验瓶中的果蝇在选择配偶上有明显的倾向性:淀粉培养基上培养的雌果蝇倾向于选择相同培养基上培养的果蝇,麦芽糖培养基上培养的雌果蝇也更倾向于选择麦芽糖培养基上培养的果蝇。自然界中也观察到类似的结果,如生活在美国内华达州死亡谷(death valley)中小湖泊内的数十种内华达鳉鱼(Cyprinodon spp.),它们分别生活在以前连续而现在分隔的有限的几个水体中,它们都是由共同的祖先进化而来,由于干旱的原因将原先联结在一起的种群分隔成数个小种群并分别进化,形成生殖隔离而成现今不同个种群。而生殖隔离的形成一般需要多久的时间呢?一般隔离机制形成平均需要1.5百年~3.5百年,也可以更快。而隔离机制的产生需要多少基因参加呢?如果突变的基因存在上位效应的话,就可能产生隔离而形成不同的物种。根据Coyne的研究发现,只要X染色体上有一个基因改变就可以导致不育,而常染色体至少要有4个基因的参与,才能产生隔离机制。
物种形成方式基本上有两种:渐变式的物种形成和骤变式的物种形成。在物种分布区内,先由地形、地貌等外界物理因素阻止种群间的基因交流,从而促进种群内遗传差异逐渐、缓慢地增长,通过若干中间阶段,最后达到种群间完全的生殖隔离和新种的产生,即为渐变式的种形成。种群内少部分个体,由于遗传机制或随机因素(如显著的突变、遗传漂变等)而相对快速地获得生殖隔离形成新种,即谓之骤变式的物种形成,它是通过个体变异实现的。
渐变式的种形成过程是缓慢的,同时也具备完整的中间过程,这也是物种形成的主要方式。一般包括三种不同的演化模式:异域种形成、邻域种形成和同域种形成。如果初始种群足够大,那么这一种群可能被一些后来形成的自然的地理障碍(如河流改道、山峰隆起、火山喷发等)隔离成两个或多个小种群,这些小种群间不能进行基因交流。如果隔离的时间足够长,各小种群适应自己的生活环境而进化,即可能形成生殖隔离,而分别进化成新种,这就是异域种形成的机制,也称为分布区不重叠的种形成,这也是最早提出的物种形成的模式。而在同一地区内,由共同的初始种群分化成两个或更多的新种群的情况就是同域种形成。这一物种形成模式没有明显的地理隔离,这种模式下基因交流的阻断方式主要有:生态位异化、多倍体化两种途径。生态、行为上的歧化选择可形成生态或行为隔离,从而形成生殖隔离,如澳洲Lord Howe岛的两种棕榈树(Howea spp.)在距今690万年前为同一种群,在开花时间和土壤偏好上分开,使其分别进化成两个物种。多倍体化是促进植物进化的重要力量,蕨类植物中多倍体种类可能占到97%,被子植物中的多倍体频率为30%~35%,如亚婆罗门参属的Tragopogon dubius和Tragopogon mirus,后一种是四倍体,可能是在不到100年的时间进化而来。另外一种模式就是领域种形成模式,这一模式是由Bush提出的,指在没有地理障碍的情况下,初始种群的地理分布区相邻接而不完全隔开,种群间个体在边界区有某种程度的基因交流,这种情况下的物种形成即被称为“邻域种形成”。
进化并非总是匀速、缓慢、渐变的,快速、跳跃式的进化也同时存在,称为量子进化。这种骤变式的种形成的一种可能途径是通过遗传系统中特殊的遗传机制,如转座子在同种或一种个体之间的转移,个体发育调控基因的突变,杂交、染色体结构变异,以及染色体组增加或减少等途径实现。另一种途径涉及随机因素和环境隔离因素,即小种群在遗传上快速偏离其母种群。在有一定程度环境隔离的小种群中,由于遗传漂变和自然选择的效应,比较容易发生遗传组成上快速偏离母种群,发展为新的物种。
当环境发生剧烈变化,而物种本身缺乏合适变异来适应变化的环境时,可导致物种的绝灭。据估计,地球上存在过的物种中约有99%已经绝灭。生物的绝灭可以分为两类:常规绝灭(normal extinction)和集群绝灭(mass extinction)。常规绝灭如同种的形成一样,是进化的正常过程,它以一定的规模在各个时期经常发生,表现为各分类群中部分物种的替代,即新种的产生和某些老种的消失。其产生的原因主要是:a.物种内在的原因,物种自身结构在进化过程中高度特化大大限制了其自身的进一步发展,使后代不能适应新的环境变化,而自然绝灭; b.物种间的生存斗争,但由此导致的绝灭事件不常发生;c.隔离在相对较小的分布区内,物种长期在相似的环境条件下生存,缺乏竞争,缺乏应对环境变化的能力,也容易导致绝灭。另外,在生命史上还发生过多次非正常的大规模的绝灭事件,在相对较短的地质时间内,大量物种消失,即为集群绝灭。在整个地质历史上,共有5次大的集群绝灭事件发生:发生在晚奥陶纪的绝灭事件大概有22%的海洋动物科消失;发生在晚泥盆纪的绝灭事件大概有21%的海洋动物科消失;发生在晚二叠纪的绝灭事件最为宏大,大概一半的海洋动物科消失;晚三叠纪的绝灭事件使20%左右的海洋动物科消失;而发生在最近的晚白垩纪的绝灭事件大概有15%的海洋动物科消失,我们熟知的恐龙就是在这一绝灭事件中消失的。集群绝灭的原因可能比较复杂,每次绝灭的原因也不尽相同。比较流行的是灾变论,认为主要是宇宙和地球演化过程中出现的一系列诸如超新星爆发、外星球撞击、地球磁极倒转、大规模火山爆发等灾变事件引起生物的集群绝灭,如晚白垩纪的小行星撞地球后引起的热浪、尘埃、降温等一系列环境后果导致恐龙等动植物的消失。
物种的形成在生物进化中具有重要的意义:
(1)物种形成是生物对不同生存环境适应的结果。生物生存的环境具有变化性和异质性,环境随时间变化导致生物的适应进化,环境在空间上的异质性导致生物的分异,分异的结果是物种形成。不同的物种适应不同的局部环境,不能设想有能够适应各种不同环境的一种生物。各种生物在进化过程中不断分化、歧异产生更多的物种意味着生物能够占领更多的生存环境,生物的不连续性是生物对环境的不连续性的适应对策。物种形成不仅表示生物类型的增加,而且为新生物类型提供新的进化起点。
(2)物种间的生殖隔离保证了生物类型的稳定性。物种间生殖隔离的存在,保持了种群基因库的相对稳定性。没有物种间的生殖隔离就不能通过进化获得新的适应,杂交会使新的适应因融化而丢失。所以,物种的存在使生物既保持遗传的相对稳定,又使进化不致停滞,成为进化的基本途径。
(3)物种是生物进化的基本单位。物种具有可变性以适应环境的变化,但这只是相对的,一个物种不能永远适应变化的环境。当环境变化的速度和范围超出旧物种适应能力,旧物种的灭绝就会发生,这时新的环境也有待新物种去占领。生态系统也要适应环境的变化,物种的更替和种间生态关系的改变,可以使生态系统适应变化的环境,生物与环境之间从不平衡又达到新的平衡,从而推动整个生物界的进化。
(4)物种是生态系统中的功能单位。不同的物种因其不同的适应特征而在生态系统中占有不同的生态位。物种是生态系统中物质与能量转移和转换的环节,是维持生态系统能流、物流和信息流的关键。
二、生物的宏观进化
宏观进化在20世纪40年代由美国遗传学家戈德斯密特(Goldschmidt R)和古生物学家欣德沃尔夫(Schindewolf O)提出,其研究的是长时间尺度的进化,对于整个生命史来说,具有决定性意义。
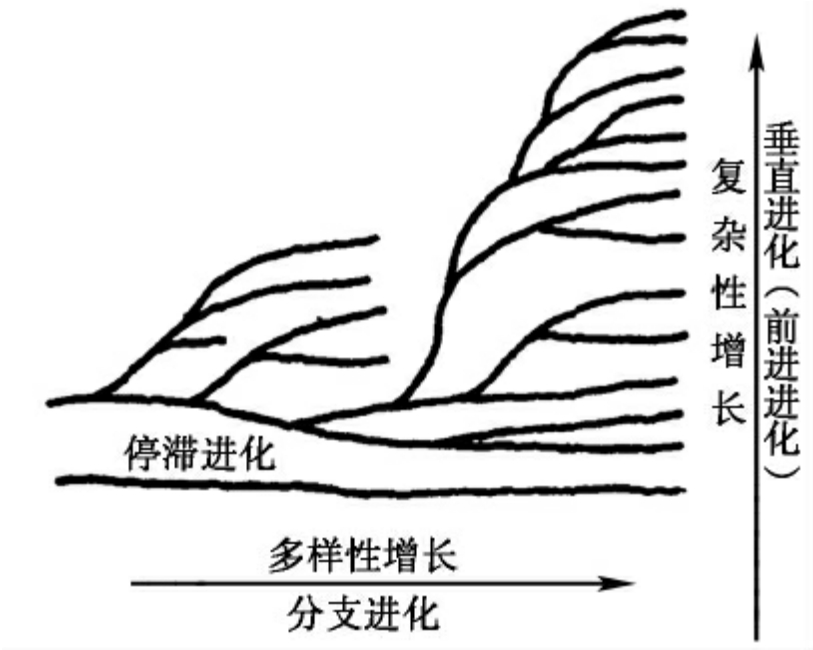
图7-9 水平进化与垂直进化图解
1.生物宏观进化的型式与趋势
按照进化的方向,可以将宏观进化分为垂直进化(vertical evolution)和水平进化(horizontal evolution)(图7-9)。
垂直进化是使生物结构变复杂的进化过程,也称为前进进化(anagenesis),是随着时间的推移,生物个体结构的复杂程度呈增长趋势的进化。一般通过三个途径来实现:叠加组合、渐进的适应变化、旧器官改造。两个或多个独立起源的生物结构通过共生组合而逐渐发展成新的更复杂的新结构,即为叠加组合。这种进化模式往往出现在生物进化的重要时期,如真核细胞的起源可能是通过若干个原核细胞内共生而产生的,而蛋白质和核酸可能是分别独立起源的并通过叠加组合形成复杂的生物分子系统。多细胞生物的起源可能是相对独立的单细胞个体聚集成群,或通过细胞间的共生组合途径逐渐发展为复杂的个体结构,这也可以看成是叠加组合。新器官出现之初,其功能是不完善的,其结构是相对简单的,器官在适应进化过程中其功能逐渐完善的同时,形态结构趋于复杂,即渐进的适应变化。同源器官的比较生物学和解剖学研究也提供了大量的实例,如心脏的进化,从鱼类的单心室、心房,到哺乳类和鸟类的左右心室、心房完全分隔,心脏的结构也趋于复杂化。适应某种功能的器官在一定条件下朝着适应于新功能的方向特化,最终改变为相对完善的适应新功能的复杂器官,即旧器官改造。如陆生脊椎动物的肺可能是从类似于鱼类的鳔进化而来,在适应从空气中获取氧气的呼吸过程中,将鳔改造及复杂化而形成肺这样的器官。
水平进化是导致生物分类学多样性增长的进化过程,即分支进化。在进化随着时间推移使生物体复杂性增加的同时,生物体在形态、生理上产生多方向的分化,以适应各种不同的生活环境,从而使区域或全球的生物多样性增加。一般包括:趋同(convergence)、趋异(divergence)、平行(parallelism)和重复(iteration)等形式。
亲缘关系较远的物种,由于长期适应相似的生存环境,具有相似的生活方式,而各自独立进化出相似的表型的现象称为趋同。如哺乳类的蝙蝠、爬行类的翼龙以及鸟类,为适应于空中的飞行,分别进化出相似的翼的结构。来源于一个共同祖先的物种,由于适应不同的生存环境,具有不同的适应方向,而在一些性状上产生歧化,从而能够占领不同的生态位的现象称为趋异。如第四纪更新世的一次大冰川将一群棕熊祖种群分隔到北极严寒环境中,从而是棕熊祖种分别进化成现代棕熊(Ursus arctos)和适应北极生活的北极熊(Ursusmaririmus)。如果某一类群的趋异向着辐射状的多种不同进化方向发展,适应于各种多样不同的环境条件,这种多方向的趋异又称为辐射(radiation),也称适应辐射。如哺乳类发生过三次适应辐射,最近一次是新生代的由原始食虫类分化出包括灵长类在内的胎盘哺乳类,有胎类的祖先是一种食虫的短腿五趾古兽,从这个祖先种演化出丰富多样的胎盘哺乳类,如善于奔跑的鹿和狼、适于林中生活的松鼠类和灵长类、能飞翔的蝙蝠、适于水生生活的鲸鱼和海豚、善于营洞穴生活的鼠类和鼩鼱。适应辐射产生的原因可能是:a.大规模的物种灭绝后,种间竞争压力减少,空的生态位出现,使得快速的种形成;b.某一物种在进化中获得“关键性的更新”而进入一个新的适应域;c.一个物种迁入一个分散的、隔离的环境,或迁入地形复杂的环境,从而隔离成许多小种群,这些种群分别进化而没有基因的交流。趋异使得相关物种间具有来自共同祖先的同源器官或同源特征,而趋同进化的成员之间不具有共同的祖征。
从同一祖先演化而来的物种,开始生活在不同的环境条件而发生趋异,之后又遇到相同环境而发展出对相似环境的相同适应,这种情况下可产生平行进化。如澳大利亚的有袋类和其他大陆的真兽类在形态和生活习性等方面非常相似,袋狼(Thylacinus cynocephalus)与狼(Canis lupus)相似,袋飞鼠(Petaurus spp.)与鼯鼠(Petaurista spp.)相似,小袋鼠与老鼠相似。而重复进化是指一个演化支系的历史中,在不同时间重复衍生出形态相似的分支,在每个重复分支之间常有上百万年的间隔。这种进化形式只有在化石记录完整、地理分布广泛、高精度采集样品的情况下建立,如有孔虫的进化。
生物不仅有由简单向复杂的进化,也有由复杂向简单的进化方式,称为简化式进化,也称为退化。形态和生理上的退化表象为生物的大多数器官退化、个别器官比较发达,一般是生物对寄生和固着生活方式的一种适应,如寄生于体内的蛔虫、绦虫、钩虫等。
纵观生物进化史,不难看出生物进化的趋势表现为由少到多、由低级到高级、由简单到复杂、由一般到分化的趋势。对进化趋势的认识,随着进化生物学的发展,也经历了一个发展过程。拉马克认为进化是有方向的,而决定生物进化方向的因素是在生物体内部。达尔文虽不同意进化都是“定向”的,但也不否认进化有一定的定向性,且他认为进化的一般趋势是适应局部环境,环境通过自然选择控制进化方向。现代进化生物学也提出了不少解释进化一般趋势的原因的理论:a.由于生物本身发育的抑制,使得某些变化比别的变化更可能发生;b.环境在时间上经历着定向的改变,因而当一个生物谱系适应于环境发展时,定向进化就发生了,后者环境稳定,但存在一个对环境的最适表型,也促进定向进化的发生;c.采用计算机模拟进化的随机趋势表明:在小的分支内,随机的种形成和随机绝灭也会造成谱系的偏斜。
2.生物圈的进化
最初的生态系统是从一个缺氧的无生机状态下开始的。Cloud叙述了生物圈早期进化的8个主要阶段(表7-2):大约38亿年前,地球上生命起源开始;几亿年后,形成了原始生命;大概又过了8亿年左右,出现了蓝菌,蓝菌的光合自养使空气中的氧气含量不断增加;大约20亿年前,蓝菌是地球的优势物种,超高能途径的氧化代谢作用出现,并且这时地球上出现了第一批红层,意味着一个永久含氧大气圈产生了;大约20亿年至14亿年前,真核细胞出现,这标志着三种特征进化:有丝分裂、减数分裂、遗传重组的高级进化作用;6亿7千万年前至5亿5千万年前之间,真核细胞继续繁殖,多细胞生物出现,标志所有生命都在地区上存在的显生宙的开始;至5亿5千万年前,出现了寒武纪动物群这一世界范围内的古生物群,这一时期大气中的含氧量已达到10%;又过了大约1亿5千万年,即大约4亿年前,海洋里大型鱼类、陆地的动物植物也蓬勃发展,此时大气氧含量接近21%,也意味着生物圈进化到了现代水平。
表7-2 生物圈早期进化的主要阶段

关于生物圈的进化,英国科学家拉伍洛克(Love lock JE)还提出了著名的盖亚假说(Gaia hypothesis)也称为大地女神假说。该假说对于理解生态系统形成、发展和稳定性有着极为重要的意义。该假说认为:当地球的大气组成、地表温度、酸碱度和海水的盐分含量等受到自然条件变化的干扰,如太阳辐射、地震、火山爆发、大陆漂移等,地球上所有生命的总体就会通过其生长和代谢对这些变化作出反应。换句话说,生物和其环境组成了一个自我调节的反馈系统,以此对抗不适应生物生存的环境变化。其主要理论论点如下:a.地球上所有生物都起着调控作用,生物对其环境起着不断的主动地调控作用;b.尽管地球受到频繁的干扰和破坏,但地球生态系统依然保持表现一定的稳定性;c.盖亚假说是一种进化系统,也就是地球的进化系统是由地球上所有的生物及其物质环境构成,生物与环境的协同进化使生物圈向前发展;d.盖亚假说是一种整体论,生物与环境的联系是如此密切,它们是一个整体。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。








