



4.6 Cu,Zn-SOD的外显子基因与Cu,Zn-SOD分子结构的进化机制假说
基于外显子和基因的复制的理论基础,提出一种Cu,Zn-SOD的进化机制。这种进化机制是在对牛的Cu,Zn-SOD的晶体结构分析中得出来的,每个亚基都含有两个内部折叠的对称轴,关于它的α-碳位点对称,铜离子位于对称轴的中心。锌的结合位点亚区域在稳定和催化中起作用。包括N-末端发夹结构的两个子域不是对称结构的一部分,它可能编码蛋白质折叠的一个核化位点。这些亚区域在人的Cu,Zn-SOD基因中是由外显子编码的,这支持了一个假说,即它们是在基因复制后加上去的。这个假说的一个困难是外显子编码的锌离子的结合区域也编码对称结构域,这与锌离子结合区域是在基因复制融合后加上去相矛盾。粗糙链孢霉对应64位氨基酸的内含子的发现解决了这一难题,因为它形成了一个新的外显子exonⅤ,几乎和锌离子的结合位点完全匹配而且它不属于结构对称区域。应该注意的是,exonⅤ只编码四个氨基酸中的一个,这个氨基酸是连接目前结构的锌离子的,这就说明更为广泛的氨基酸置换在提出附加的exonⅤ时就已经发生了。
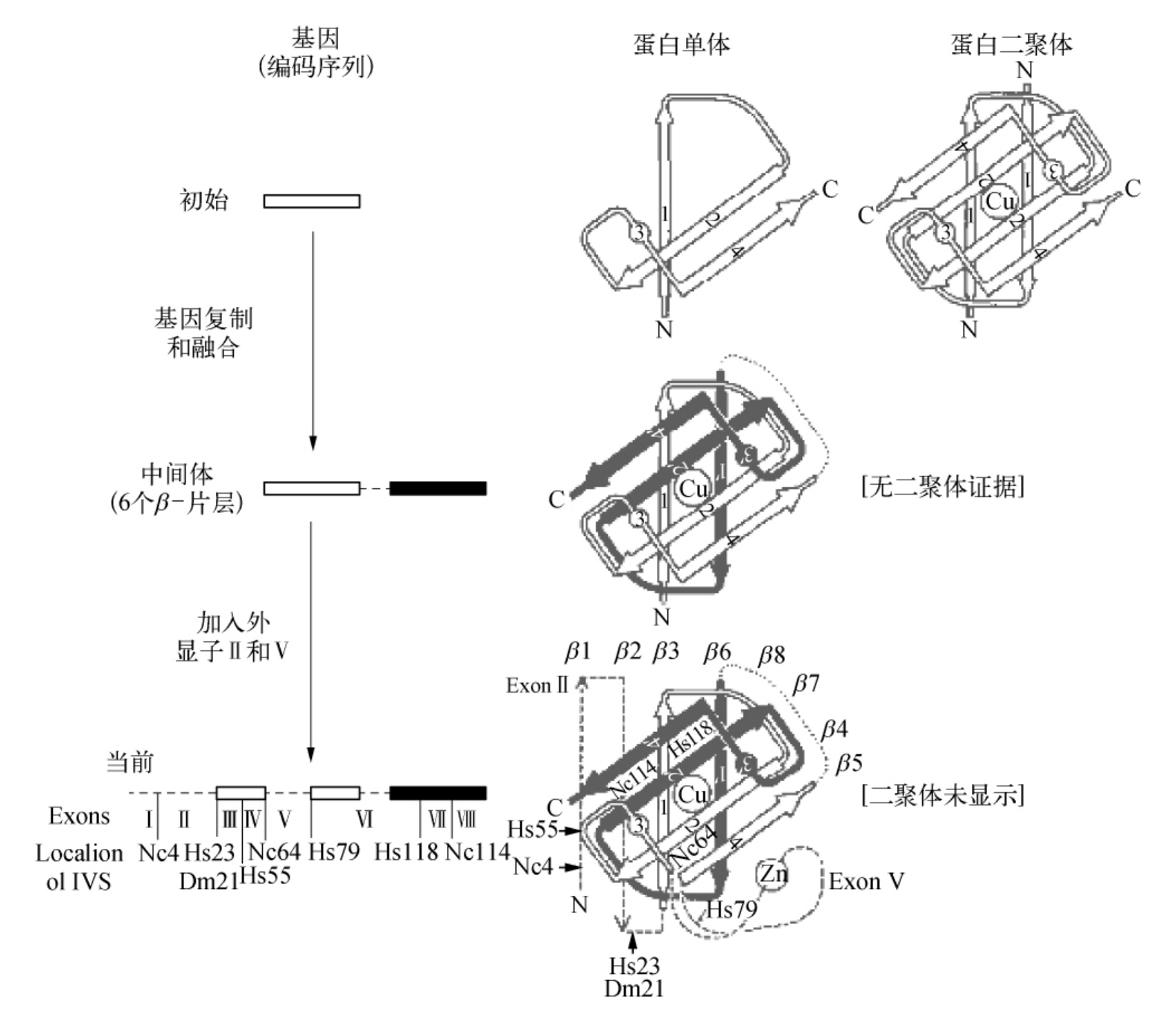
图4-7 Cu,Zn-SOD的进化机制假说
(摘自DO Natvig,et al.J.Biol.Chem.,1990)
蛋白质的结构简略地用箭头式结构表示,组成希腊钥匙β折叠桶的β片层。N和C的末端比显示的要近些,是因为分子前面的β片层的扭曲所致。在现在的结构中,锌离子和环与铜离子距离更近些,而环包含对称的3和3′比图示的要大。在结构上对称的1和1′,2和2′,4和4′对应相应的β片层,而3和3′仅包含环状残基。对称的2倍轴垂直于纸平面并且穿过铜离子,外显子Ⅱ和Ⅴ更像是最近的结构附加物,都添加到N-末端的对称区域上。为简单起见,虽然一些现在的内含子可能来自原始的结构,但原始的和过渡基因并没有表示出内含子。
粗糙链孢霉的这三个内含子没有一个与人和果蝇基因的相符合。粗糙链孢霉内含子虽然很小,但是与那些高等有机体的相似处是它们的5′和3′界面都需要GT和AG序列。Cu,Zn-SOD基因总共含有7个内含子,将蛋白质分割成8个平均长度为19个氨基酸的外显子。随着从植物和其他生物体提取基因序列变为可能,外显子的数目可能会增加。虽然外显子在蛋白质中是对应蛋白质的功能和结构区域或者亚区域,但是外显子Ⅰ和Ⅶ却因为太小而不能胜任这个角色。
有一个粗糙链孢酶的内含子对应氨基酸残基114,而人的内含子中只有4个残基对应118位氨基酸。看起来这些不协调的内含子编码的外显子使对应一个结构域和功能域不太可能,因为它非常小而且位于β链上。同样这些内含子从相同的遗传的内含子经过变化位置得到也不太可能,因为人和粗糙链孢霉的这段序列是高度保守的。另外,这两个内含子都对应蛋白质结构的包埋区域。假设包埋的内含子通常对应原始结构的表面,随着蛋白质的进化而包埋起来。至于Cu,Zn-SOD,如果上述的两种内含子在原始的β桶结构的进化后产生,那么它们的标准部件必须是小的氨基酸序列。可以选择的是,一个或者两个内含子可能是在进化的最近阶段插入的。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










