



第三节 趋化因子受体的结构
趋化因子通过其在细胞膜上的受体起作用。迄今所发现的趋化因子受体都是G蛋白偶联受体(G-protein-coupled receptors,GPCR)超家族成员,是一组具有7次跨膜结构(seven-transmembrane domain,7TM)的受体。以CCR1为例,其结构特征如图7-3所示。
趋化因子受体广泛表达于各类淋巴细胞、内皮细胞、神经元及其他一些在信号传递方面起作用的细胞类型,并引起此类细胞的脱颗粒(信号分子的释放)、淋巴细胞归巢、T淋巴细胞激活、有丝分裂效应、血细胞生成及趋化反应。某些趋化因子受体正是HIV和猴免疫缺陷病毒(simianimmunodeficiency virus,SIV)等病毒进入细胞的辅助受体。
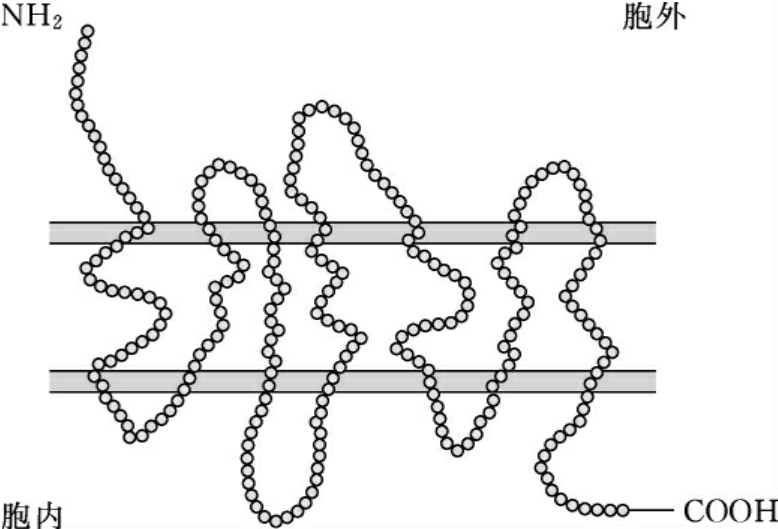
图7-3 趋化因子受体结构模式
迄今为止,已有5种CXC趋化因子受体被克隆。研究发现存在2种IL-8受体,并且都对IL-8表现出高亲和力。所有含ELR序列的CXC趋化因子都能结合1种以上的受体,但CX-CR1(IL-8Ra)只与IL-8结合,而CXCR2(IL-8Rb)可与多种趋化因子相结合。
CC趋化因子受体则更加复杂,除Eotaxin只能与CCR3结合以外,所有的CC趋化蛋白都表现出多种交叉反应性,如CCR1可与MIP-1α、RANTES及MCP3以高亲和力结合。CCR2在mRNA阶段,通过不同的剪切方式,产生了两种形式的蛋白分子,即CCR2a和CCR2b,两者只是在其胞内尾肽上存在差异。而到目前为止,还没有发现CCR2a和CCR2b在配体结合的特异性上有什么不同,两者均能与MCP-1、MCP-3和MCP-5结合。CCR2a与CCR2b在单个核细胞中的mRNA表达水平基本一致,CCR2b的胞质内尾肽富含丝氨酸和苏氨酸,且与CCR1之胞内段同源。
CCR3是从嗜酸性粒细胞中分离得到的一个高水平表达的趋化因子受体。CCR3能有效地与嗜酸性粒细胞的趋化蛋白,如Eotaxin、RANTES、MCP-2及MCP-3结合。除CCR3以外,嗜酸性粒细胞还表达CCR1,这是MIP-1α的一个受体,这就可以解释为什么MIP-1α也能趋化嗜酸性粒细胞。由于MCP-4可以阻断Eotaxin对嗜酸性粒细胞的作用,因此认为,CCR3也可与MCP-4结合。相类似地,CCR4是从嗜碱性粒细胞中克隆出的,它可与多种趋化因子如MIP-1α、PANTES、MCP-1及TARC结合。虽然CCR5与CCR2的一级序列很相似,但CCR5能以更高的亲和力与MIP-1α、MIP-1β及RANTES结合。
虽然趋化因子与其受体间的相互作用非常复杂,但目前尚未发现不同家族的趋化因子与趋化因子受体间的相互交叉作用,或者说CC或CXC家族的成员只能在其家族内部行使功能。但也有例外,如红细胞表面的Duffy抗原,它能与趋化因子结合,因此被称为DARC(duffy antigen receptor for chemokine)。DARC可结合若干个趋化因子,包括CXC与CC家族成员。由于还未发现DARC与趋化因子结合能介导信号传导,因此对DARC的生理功能尚不清楚。
趋化因子受体的另一个有趣的现象是某些病毒能产生趋化因子受体样分子。US28是CMV编码的一个分子,其结构与CCR1相似,发现US28能与CX3C趋化因子Fractalkint相结合。此外,US28还能与一些CC趋化因子结合。疱疹病毒产生的一个膜结合蛋白,结构与CXCR1、CXCR2相似。表达该重组蛋白后发现,它能像CXCR2一样与IL-8、GRO-α及NAP-2结合介导信号传导。从Kaposi’s肉瘤中分离得到的Kaposi’s肉瘤相关疱疹病毒也编码一趋化因子受体样分子。现在还不清楚病毒为什么编码这些分子。有可能通过模拟机体免疫系统的效应分子来改变机体抗病毒反应能力,或者调节病毒感染细胞的生长能力,从而达到自我保护或促进机体病变的作用。Kaposi’s肉瘤相关病毒编码的趋化因子受体样分子,能以较高的亲和力与IL-8结合。将其转染COS细胞后发现,该分子表现为持续性的生长刺激活性,能够促进感染的COS细胞的自发增生。因此,此病毒产生物在Kaposi’s肉瘤发病过程中可能具有重要作用。
趋化因子与其受体结合后,其刺激信号通过与其偶联的G蛋白传入细胞内,从而激活靶细胞的运动、黏附与去黏附、细胞骨架重排、吞噬异物、释放各种水解酶、抗微生物质以及激活NADPH氧化酶产生超氧化物等效应。趋化因子受体介导的信号传导对于细胞生理、病理功能具有重要意义。任何异常都将对机体产生不利影响,如慢性肉芽肿、中性粒细胞黏附缺陷等疾病都与趋化因子受体功能异常有关。与其他细胞因子一样,趋化因子诱导其受体的二聚体化也与其对后者的激活密切相关。
当趋化因子刺激其受体时,后者通过与之偶联的G蛋白进行信号传导,导致特征性的胞内钙离子的流动及沿趋化因子浓度梯度的细胞迁移(chemotaxis)。趋化因子和趋化因子配体偶联后细胞的一般变化如下:趋化因子与趋化因子配体结合后引起分子构象的改变,激活偶联的G蛋白形成Gα-ATP复合物并与βγ亚单位分离,通过ras/MAPK途径、N-甲酰多肽激活的Ser/Thr蛋白激酶途径和磷脂酰肌醇3-激酶途径等,分别激活一系列激酶包括磷脂酶(PLC)、磷酸肌醇三激酶(PI3K)以及酪氨酸激酶,产生1,4,5-三磷酸肌醇(IP3),促使胞外Ca2+内流和细胞储存Ca2+释放,致胞质内Ca2+浓度增高,从而激活淋巴细胞内的肌凝蛋白Ⅰ、Ⅱ,与肌动蛋白结合,导致肌动蛋白丝聚合、收缩,在多种细胞因子、黏附分子和酶等的协同作用下,细胞定向迁移、聚集。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











