



四、生物降解作用的认为强化
大量研究工作表明微生物降解环境污染物的潜力远未充分开发利用,对现有降解菌的生理遗传改造,还可以进一步提高这种降解能力。生物降解作用的强化提高是生物降解中的重要研究课题。提高生物降解能力的方法从组织层次上包括:①群体降解水平的提高,如向环境投入营养物可以从总体上提高微生物的生理活性。②微生物种群降解能力的提高,通过对降解微生物的生理遗传改造,提高降解能力。这包括生理层面的驯化适应,遗传层面的遗传修饰改造及遗传工程改造。③酶工程的降解酶改造扩大酶底物范围,提高降解能力。
从种群水平的工作程序可以包括对现存降解能力的提高以及构建新的降解能力。
1.现存生物降解能力的提高
(1)自然生物降解多样性的分析和解降能力资源开发利用
自然发生的微生物活性过去是,现在也是所有生物技术应用的出发点和基础。分离具有新的代谢能力的细菌菌株,并对它们的降解途径作出生物化学和遗传学阐述,这样有利于开发自然发生的多样性和降解能力资源。主要的应用方面是可以克隆遗传基因构建新的遗传工程菌以及构建协同式菌群,构建畅通的代谢降解路线。
在芳香化合物生物降解中,各种双加氧酶的羟化作用可以增加化合物的极性,促进生物降解,因此在这类化合物降解启动有重要作用。导入的两个羟(基)氢氧基(hydroxylgroup)位于邻位或对位,完成催化过程的酶是多成分的双加氧酶,酶是由一个电子传递链和具有催化活性的α-和β-亚基组成的。这类酶的多样性和底物范围受到特别关注。鞘氨醇单胞菌(Sphingomonas sp.)RWI菌株产生的二(dioxin)双加氧酶是最先被报告的酶,其能进行一种成角(度)(angular)的使一对邻近的碳被氧化的双氧化,其中的一个碳被包含在两个芳香环之间桥的之中(one of which is involved in one of the bridge between the two aromatic rings)。编码二双加氧酶的基因也有独特的特点,一般的双加氧酶的编码基因成簇,而这种酶却非预期地分散于染色体。另一个能进行成角(度)双氧化的咔唑(carbazole)1,9α-双加氧酶(in Pseudomonas sp.strain CA10)也有其特点,其末端的加氧酶由单一蛋白质CarAα组成,和典型(classical)的由大的α和β亚基组成的加氧酶不同。CarAα的核苷和推想的氨基酸序列也是独特的,与其他的末端加氧酶(包括二双加氧酶)的大亚基仅展现出很少的相似性。对羟化作用的大量双加氧酶的研究说明环激活的双加氧酶存在着广泛的多样性,从这个角度出发,我们还可以发现具有新的特异性的基因。
细菌降解多样性不限于环激活的双加氧酶,也包括降解芳香烃化合物代谢产物的酶。芳香环断裂是卤代芳香烃降解的主要关键反应,其降解酶的多样性也广受关注。长期以来认为通过间位(meta)裂解途径代谢3-氯代儿茶酚是不可能的,这是由于反应产物将失活二醇外(extradio)双加氧酶。但恶臭假单胞菌GJ31菌株却含有一种新的氯代儿茶酚2,3-双加氧酶,这种酶能有效地打断3-氯代儿苯酚(在2,3-位置),这导致同时环断裂和脱氯,细菌能通过间位断裂途径降解氯代苯的途径已被详细研究,抗自杀失活的残基已被定位。五氯酚和γ-六氯环己胺(林丹)也以同样方式被降解。
参与各种硝化芳烃化合物降解的新的类型的间位断裂双加氧酶也有报告。环裂解的基质通常是二酚,二酚中的二个羟基不是邻位(ortho)就是对位(para)。在代表性反应中,仅一个羟基存在于环断裂基质2-氨基酚中,其经历环断裂到2-aminomuconic半醛,氨基明显替代第二个羟基功能。
(2)表达生物表面活性剂提高异生物源化合物的生物可利用性
环境中的疏水性有机化合物长时间抗生物降解的主要原因是它们的生物可利用性受到溶解性的限制。提高它们的生物可利用性,从而促进生物降解最可能的方法是应用(生物)表面活性剂。大量的研究都表明导入表面活性剂可以促进生物降解。然而表面活性剂在生物修复中的有效性的报告是混淆不清的(mixed)。生物表面活性剂的本质作用已被解释为可以增加疏水性、水不溶性生长基质的表面积,通过增加溶解度或从表面上解吸下来,以及调控微生物对表面的吸附及脱离从而增加化合物的生物可利用性。这样加入表面活性剂对生物降解的净效率将是化合物的溶解度增加和细菌对这些化合物直接吸附降低的综合结果。表面活性剂的加入会降低细菌吸附到非水液体表面,这样降低了在疏水性化合物上的生长。实际上表面活性剂的相应效应我们还了解很少,是极其复杂的不同环境中微生物和土壤、沉积物、污染物相互作用的结果,所以在使用单一表面活性剂时往往有不同的效果。这样我们对这些相互作用还有待进一步研究了解,在使用表面活性剂时要优化表面活性剂,了解生物降解和靶环境条件的相互作用。产生生物表面活性剂的具有降解能力的菌株将有助于生物降解,将是一种最好的选择。
(3)提高细胞吸收有机污染物能力,促进生物降解
许多有机污染物要进入细胞内才能被胞内酶降解,因此把有机物运输进入细胞对生物降解也是十分重要的。许多芳香化合物可以通过能量依赖迁移系统被细菌吸收。Pao等指出有三类(three families)透性酶负责酸性芳烃化合物及其代谢产物的转运。新的研究资料表明异生物源化合物也是被特异的运输系统转运,Leveau等首先报告了一种转运2,4-二氯苯氧醋酸盐(2,4-dichloro phenoxyacetate)的运输系统(transporter)。对手性化合物(chiral)的对应选择(enantioselective)吸收也已得到证实,除草剂2-(2,4-二氯苯氧基)丙酸盐就是这样的结构,其被Sphingomonas herbicidovoransMH降解时就存在三种可诱导的质子-梯度-驱动吸收系统(inducible,proton-gradient-driven uptake system)。在降解蒽的荧光假单胞菌中也存在对非电性的(noncharged)疏水性蒽分子的主动运输系统。这些研究结果说明我们为修复而设计超级生物降解菌(或称为超级生物催化剂superior biocatalysts)时也必须考虑对污染物的吸收机制。
(4)增强对有机污染物的趋化性,促进生物降解
微生物对污染物的趋化性也可以增强发生在自然环境中的生物降解。已有研究证明恶臭假单胞菌的4-羟基苯酸盐的运输系统pcak也使细菌对这种化合物的趋化性起作用。其他许多运输系统(如透性酶)也能起到决定趋化性的化学受体作用,它们和已描述的化学受体是不同的。Grimm和Harwood报告说,恶臭假单胞菌的降解质粒(NAH7)上的nahy基因编码的膜蛋白Nahy也决定着对萘的趋化性。它的羟基-末端区类似于趋化性的转换器(transducer)蛋白,因此具有对萘化学受体的功能,其也可以成为像联苯这样相关的化合物的化学受体。对趋化性遗传基础的研究将使我们能够利用趋化性的特征来增强生物降解作用。
(5)改变细胞表面特征提高适应环境和生物降解能力
许多高度疏水性的环境污染物(如甲苯等)对微生物具有很强的毒性,其可以积累在细胞膜上,干扰细胞膜,使细胞失活,从而就会阻碍所希望的生物降解。具有降解能力的微生物也会因细胞膜受损而失去其降解功能。现在已经发现对有毒污染物的耐受能力是它们具有降解能力的重要因素。
耐受能力的原理主要有三种。
①细胞膜组成成分中脂肪酸从顺式(cis)转变成反式(trans)的异构化可以提高耐受能力。反式脂肪酸较高的刚性使膜结构对有机溶剂的干扰敏感性较小。编码使脂肪酸从顺式异构化成反式的酶的基因已被克隆。使溶剂耐受性增强的微生物磷脂生物合成量增加的现象已被观察到。
②通过修饰外膜蛋白和脂多糖来降低细胞表面的疏水性,从而减少有机溶剂在细胞膜上的积累,这从另一个角度提高耐受性。
③微生物存在溶剂泵出系统,从而提高耐受性。
许多分离出来的溶剂耐受细菌已被证明能降解矿化有机溶剂,它们的代谢能力已被用于工程化的生物修复。在许多芳烃严重污染的地方,溶剂耐受菌成为最先的定殖菌,成为这些污染物去除的优势菌。使用有降解能力的溶剂耐受菌可以成为生物修复有前途的方法。
对有机溶剂的这种耐受性我们实际上可以理解为一种适应,一种对环境的适应。适应环境是生物降解的前提条件,有人已构建出重组子耐放射异常球菌(Deinococcus radiodurans),其能在高辐射环境下氧化甲苯、氯苯等污染物;用于极端环境修复的极端降解菌已得到应用,例如在北极和南极地区,降解PCB的耐冷细菌已分离出来。细胞表面的疏水性的改变不仅代表一种对有机溶剂的防御机制,同时也对细菌表面吸附产生影响。在生物修复中,我们希望外源接种细胞不是堵塞在周围而是迁移到所希望的地方。黏附缺陷型菌株在运动迁移上具有优势,现在能在土壤中快速迁移的突变株已被筛选,并已得到实际应用。
2.发展新的生物降解能力
(1)构建降解型基因工程菌
构建组合式、互补式降解能力的新菌株可以大大提高微生物的降解能力。构建超级生物催化能力的一种策略是把源于不同生物的降解片段合理组合并转移到一种受体菌株,从而在对异生物原物质生物降解中避免形成截止式产物或毒性更强的代谢产物,因而达到完全的降解过程。这种策略已被成功用于降解高毒性化合物三卤代丙烷(trihalopropanes)。例如Bosma T等先构建一个广宿主范围质粒(broad host-range plasmids),这种质粒含有编码卤代(键)烷烃脱卤酶的基因(来自Rhodococcus sp.Strain M15-3),可在不同的异源启动子控制下实现表达,这种酶能有效地把三卤丙烷转化成二卤丙醇。
有研究者在氯苯酸盐降解途径上结合一段氧化途径,这种途径能把(氯)二苯转化(由bph基因编码酶进行)成(氯)苯酸盐。实现这种组合的方法是通过细胞的接合融合得杂合菌株,或把bph基因转入氯苯酸盐降解菌。Hrywna Y等克隆出氯苯酸盐邻位和对位脱氯酶的编码基因,并在降解二苯和共代谢氯代二苯菌株(睾丸酮丛毛单胞菌Comamonas testosteroni strain VP44)中表达。结果表明构建的工程菌株能生长并对2-和4-氯二苯脱氯降解(构建基因工程菌见本书第十五章)。
(2)构建转基因植物,提高植物的净化能力
植物已被广泛用于污染环境的生物修复。把来源于细菌的遗传信息转入植物将更有利于提高植物的净化能力。转基因植物白杨(poplar plantlets)能表达细菌的汞还原酶,已经证明能在离子汞表现出毒性的水平下萌发和生长,并能释放出元素汞,因此能有效地把土壤中的结合汞排出。同样转基因植物能表达修饰的有机汞裂解酶,它们能旺盛生长在较高浓度的高毒性有机汞条件下,有机汞裂解形成的离子汞会积累在可任意利用的植物组织中。阴沟肠杆菌PB2(Enterobacter cloacae PB2)能以TNT作为N源生长,其表达的季戊四醇四硝酸盐(pentaerythritol tetranitrate)还原酶能还原TNT的芳香环,并释放出亚硝酸盐。把细菌这种酶的基因转入烟草,其种子也能表达这种还原酶,也能在丙三氧(基)三硝酸盐或TNT毒性浓度条件下萌发和生长,而此时野生型种不能萌发和生长,其幼苗也比野生型幼苗对丙三氧(基)三硝酸盐有更快、更完全的脱硝能力。实际上转基因植物在生物修复中可以作为细菌的替代物。
(3)降解酶的定向改造和进化,提高降解能力
酶是生物化学反应过程的核心,各种降解酶也是生物降解过程中的关键因素。研究酶基因的克隆和表达、酶蛋白的结构和功能的关系以及对酶进行再设计和定向加工的基因工程、蛋白质工程方法和技术的发展及进步为发展更优良的新酶或新功能酶提供了广阔的前景,这也为降解酶的发展提供了新的技术手段。
酶分子本身蕴藏着很大的进化潜力,许多功能有待开发。目前酶工程主要采用基因定点突变(site directed mutagenesis)和体外分子定向进化(in vitro molecular directed evolution)两种方式对天然酶分子进行改造。
1)基因定点突变。基因定点突变的基因突变是在了解酶蛋白的三维空间结构及编码序列,搞清结构与功能关系的基础上,根据蛋白质的空间结构知识来设计突变位点,然后通过点突变使已知的DNA序列中一定长度的核苷酸片段发生替换、插入或缺失。而改变蛋白质结构中特定位置(如活性中心)的氨基酸残基,从而改变酶的特性,最后通过筛选选出有益的突变从而提高酶的活性或开发了新的酶。目前已利用定点突变技术改进天然酶蛋白的催化活性、抗氧化性、底物特异性、热稳定性及拓宽酶反应的底物范围,改进酶的别构效应(allosteric effect)。这种方法与使用化学因素,自然因素导致突变的方法相比,具有突变效率高,简单易行,重复性好的特点。
点突变的方法及技术也可以用于降解酶的改进,并且已显示重要的应用前景。卤代烷烃(haloalkane)脱卤酶是异生物源化学物降解中最先受到关注的酶,为了详细了解不同卤烷烃脱卤酶的专一性,蛋白质序列和酶的三维结构模式已被进行了比较,功能上重要的氨基酸被指出,这些氨基酸可以作为将来定位诱变实验的目标。根据已知的三维结构,Vollmer等构建了一个黏康酸盐异构酶(muconate cycloisomerase)(参与自然芳香化合物的降解)的变异株,其结合穴(binding cavity)中的氨基酸也见于氯(代)黏康酸盐异构酶(chloromuconate cycloisomerases)(参与氯化芳香烃的降解)的相同位置,并能增加某些氯代黏康酸盐的特异性常数(specificity constants),然而在许多其他方面,突变体酶保留了野生型的特点,这说明不论结合穴是简单改变,还是更复杂的改变都可以造成酶的可见差异。
在不能取得结构信息时,对酶之间的氨基酸序列所作的对比分析能找出对催化活性有重要作用的氨基酸残基。通过具有不同底物专一性或催化特点的相关酶的杂合体(hybrids)的分析,可以得到关于酶决定基质专一性的酶区域(regions)的信息。这种方法已被用于查明联苯双加氧酶底物专一性差异的相应的残基。Parales等的研究表明2-硝基甲苯2,3-二加氧酶的α-大亚基的C-末端区域决定的酶的专一性。BeilS等研究甲苯和氯苯双加氧酶产生的杂合酶(hybrid enxyme),结果证明甲苯双加氧酶活性中心(active site)附近一个氨基酸从Met220(甲硫氨酸)转换成Ala(丙氨酸)能使酶对1,2,4,5-四氯苯转化和脱氯。
有研究结果说明二苯双加氧酶(伯克霍尔德氏菌CB400菌株)的底物专一性是由活性中心的三个区域决定的。ZielinskiM等进一步研究这些区域中的23个氨基酸的效应。结果证明替代这些氨基酸会直接影响与底物的相互作用,另一方面也证明不与底物相接触的许多氨基酸(Ile 243,Ile 326,Phe 332,Pro 334,and Trp 392)也强烈改变中心双加氧作用的表现。这说明预测中没有影响的氨基酸残基也可以对专一性起关键作用。这样应用定位突变可以使一种单一的酶扩大其生物降解基质的范围。
2)体外分子定向进化。酶的体外分子定向进化是在人工模拟自然进化过程的条件下,通过容错PCR、DNA改组、交错延伸、随机引物引导重组和递增截短等方法对编码酶的基因进行随机突变和体外重组,经高通量筛选获得性能更优良或全新的酶。
酶定向进化通常分三步进行:第一,通过随机突变和(或)基因体外重组创造基因多样性。第二,导入适当载体后构建突变文库。第三,通过灵敏的筛选方法,选择阳性突变子。这个过程可重复循环,直至得到预期性状的酶。其中获取多样性基因是整个工作的基础,是酶的定向进化成功的关键。
①基因随机突变。容错PCR(error-prone PCR)技术、化学诱变剂介导的突变、致突变剂产生随机突变和随机寡核苷酸突变可以造成基因随机突变。容错PCR技术是一种相对简单、快速廉价的随机突变方法,通过改变PCR反应条件,使扩增的基因出现少量碱基错配,从而导致目的基因的随机突变。化学诱变剂(如羟胺)直接处理带有目的基因片段的质粒也可产生随机突变,然后用限制性内切酶切下突变的基因片段,克隆到一定的表达载体中进行功能筛选。致突变株体内的DNA突变率比野生型高出数千倍,将带有拟突变基因的质粒转化到致突变株内培养,也可以产生随机突变,频率一般为1/2000。
②基因体外重组。体外重组有同源基因重组(homology-dependent gene recombination)和非同源基因重组(homology-independent gene recombination)。同源基因重组在体外重组中占有重要地位。
a.同源基因重组。同源基因重组的主要方法包括DNA改组、家族DNA改组(family DNA shuffling)、交错延伸过程(staggered extension process,StEP)、随机引导重组(random priming recombination,RPR)等方法。DNA改组系将一群密切相关的序列,如多种同源而有差异的基因(或一组突变的基因文库),在DNaseⅠ的作用下随机切成小片段,这些小片段可通过自身引导PCR(self priming PCR)延伸并重新组装成全长的基因。这些重排产物的集合又称为嵌合文库(突变文库)。再对嵌合文库进行筛选,选择改良的突变体。家族DNA改组(也被称为自然发生的同源序列的shuffling或DNA shuffling)采用一系列天然存在的同源性较高的基因作为起始基因进行DNA改组操作。此法存在重组子产率低的问题,解决这一问题是减少亲本背景,人们进行了两项改进,以单链DNA(ssDNA)替代双链DNA(ds-DNA)或用限制性内切酶取代DNaseⅠ。由此形成的DNA片段没有交错重复,从而减少了同源双链(home duplex)的形成,提高重组频率。这种方法已在降解酶定向进化中得到应用。交错延伸过程是在DNA改组的基础上发展起来的简化的DNA改组方法。这种方法将含有不同点突变的模板混合,短暂地进行退火(annealing)及延伸反应;在每一轮中,那些部分延伸的片段根据序列的互补性与不同模板退火并进一步延伸,反复进行,直到获得全长基因片段。随机引导重组利用随机序列引物产生大量互补于不同部分模板序列的DNA短片段,由于碱基的错误掺入或错误引导,这些DNA片段中也含有少量的点突变,在随后的PCR反应中,DNA小片段可相互同源引导和扩增成全长基因,然后克隆到适当的载体上表达并通过适当的筛选系统加以选择。
b.非同源基因重组。非同源基因重组不要求酶基因序列的同源性。主要的方法是递增截短法(incremental truncation),这种方法的核心是以核酸外切酶Ⅲ代替DNaseⅠ对靶序列进行切割,由于其5'-3'外切核酸酶的作用,因此得到的递增截短片段库(incremental truncation libraries,ITLs),理论上包括了靶序列DNA单碱基对删除的各种情况,使得在较低的复性温度下,可实现非同源序列间发生重组。依靠这种方法可获得不依赖DNA序列间同源性的杂合酶。
c.选择和筛选。当突变体酶赋予细胞生长或存活优势,赋予寄主对药物的抗性或可满足营养缺陷型菌株的生长需求时,可容易地从有106以上的酶蛋白突变体的文库中筛选出所需要的酶。
d.家族DNA改组在降解酶定向进化中的应用。联苯双加氧酶(BphDOX)催化联苯和相关化合物开始的氧化。这种酶是一种多组分酶(multicomponent enxyme),其中的一个大亚基(由bphA1基因编码)决定基质的专一性。Suenaqa A等以类产碱假单胞菌(Pseudomaonas pseudoalcaligenes KF707)KF707菌株和洋葱伯克霍尔德氏菌LB400菌株(Barkholderia cepacia LB400)的bphA1作为同源基因进行DNA shuffling,得到许多定向进化的Bph DOX,其中一个大肠杆菌克隆表现出对苯、甲苯和烷基苯极强的降解能力。这种酶中的4个氨基酸残基(H255Q,V2581,G268H和F277y)从KF707改变成LB400酶。随后的定点突变说明这些氨基酸决定单环芳香烃的降解。
Barriault D等以来源于伯克霍尔德氏菌菌株LB400、睾丸酮丛毛单胞菌B-356(Comanonas tostosteroni B-356)和圆红球菌P6(Rhodococcus globerulus p6)的bphA基因的关键片段作为同源基因进行家族DNA改组,以儿茶酚代谢物作为检测手段所得到的几种BPhA突变体表现出更强的降解PCBs的能力,也显示出对2,2'-,3,3'-和4,4'-一氯二苯的更强的降解活性。
(4)未能培养微生物生物降解信息资源的开发利用
目前得到充分利用的生物降解信息资源主要来自可培养微生物,但未能培养微生物也含有丰富的降解信息资源,这部分资源的开发利用对生物降解的强化可以起重要作用。
一般认为环境样品中的微生物中仅有3%可被培养,而其余的97%未能培养,前者称为可培养微生物(culturable microorganisms),而后者称为未能培养微生物(unculturable microorganisms),也被称为活的未能培养微生物(viable butnonculturablemicroorganisms)。从遗传学上说未能培养微生物中的一部分有与可培养微生物同样或相似的系统发育水平,但因对其生理、营养需求知识缺乏而不能应用现行培养技术培养出来,或因它们某些生理学原因抗拒现行培养方法而不能培养。而另一部分则代表新的完全与可培养微生物不同的新谱系(novel lineages),它们根本不能用标准的方法加以培养。
现在所进行的研究工作说明污染环境中不但可培养微生物携带有降解污染物的遗传信息,未能培养微生物也携带有贡献于生物降解的遗传信息。
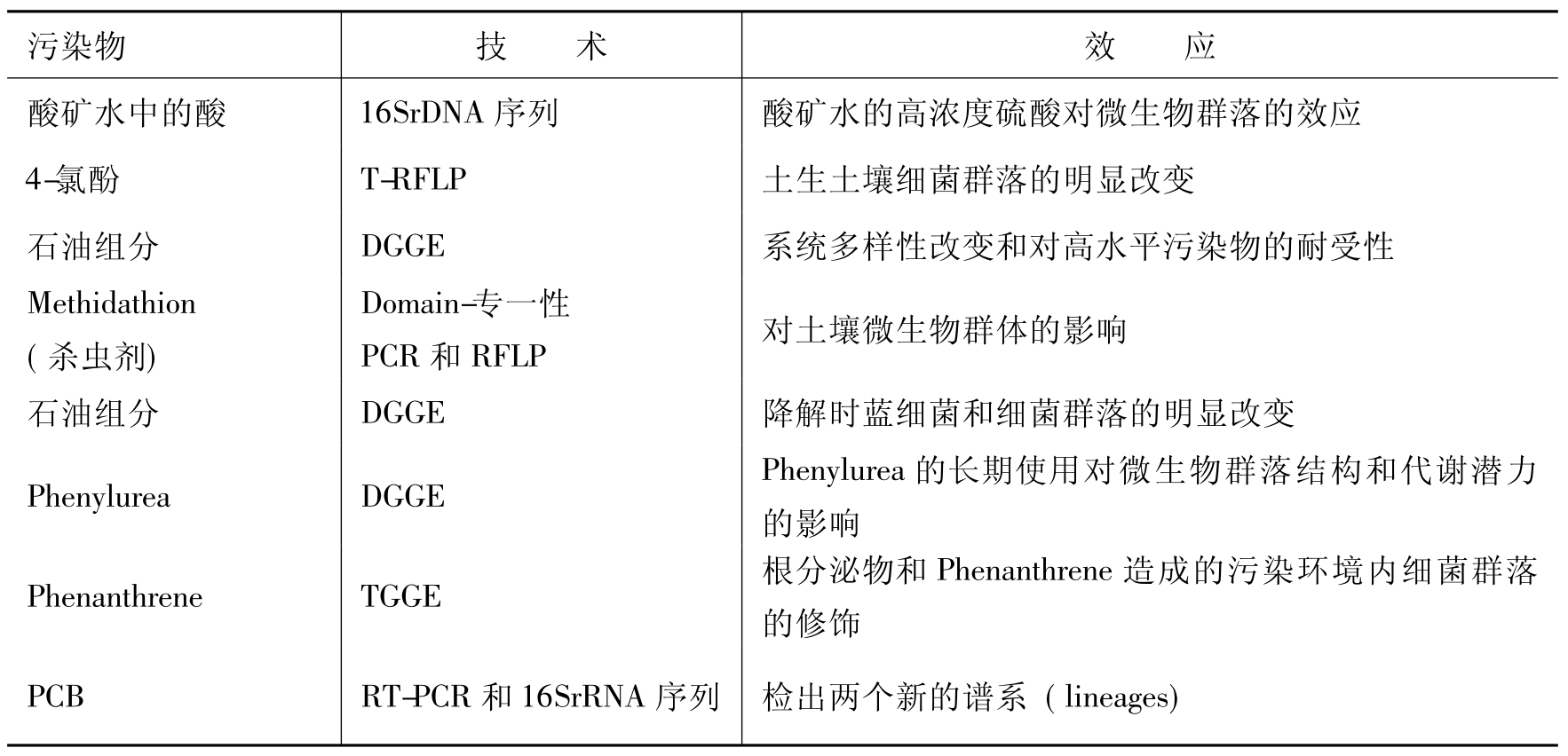
传统上的微生物多样性依赖于微生物培养技术,而现在评估多样性的分子生物学方法已经跨越传统的培养的方法,成功地用于评价和解释各种环境上的分类多样性,这也包括各种污染地点的多样性(表8-1)。最近,分子技术已发展了特征化和监测各种环境的功能多样性,图8-3是其中的流程图表。Abed RM等应用这种方法研究受石油烃严重污染地在污染物降解后的微生物聚丛及其群落的变化,受污染地生态系统微生物多样性的分析表明这种系统多样性包括培养和未能培养微生物的多样性,两者的多样性都贡献于生物降解。Marchesi J R和Weightman AJ比较研究可培养的α-卤代羧酸(α-halocarboxylic acid)降解菌的脱卤酸基因库和环境样品的总基因组(metagenome)(宏基因组)直接分离的核酸(direct isolation of nucleic acids),结果表明革兰氏阳性细菌占有总多样性的显著部分,然而由于培养的障碍,建立在富集技术基础上的研究至今尚未报告有革兰氏阳性的α-卤代羧酸降解菌。Mac-Naughton S J等用磷脂脂肪酸和变性梯度凝胶电泳分析方法对照研究了石油污染地和未受污染地的微生物群体差异,发现对照点缺乏α-变形细菌(α-proteobacteria),而实验点却含有范围广泛的该类细菌。这说明有受污染地中未能培养的微生物参与石油烃的生物降解。

图8-3 最新生物技术研究污染环境的分类和降解多样性
注:BAC:细菌的人工染色体;FISH:荧光原位杂交;RNA-SIP:RNA稳定同位素探针。
表8-1 分子生物学指标指示污染地中污染物对群落结构的效应
从环境样品中提取的总基因组(基因片段)是巨大的可利用资源,它们可以被克隆到细菌的人工染色体,fosmids或cosmid载体,进行序列基础和表达基础的筛选,或者对整体的基因组序列(以功能序列为基础)直接进行筛选,从而分离和筛选出降解污染物的操纵子。同时利用这种资源可以建立宏基因组的巨大数据库,进一步推动宏基因组的开发利用,这将促进更加有效的“设计型生物催化,促进生物修复。”
设计超级生物催化剂能力的策略最需要的是增大降解生物在环境条件下的可靠表达,从这个角度说理解异生物源化合物和生物之间相互作用的研究以及微生物在环境的归宿、存活和活性,同时还和生物化学和遗传工程研究相交织在一起,这样一种交互(cross feeding)将提供成功干涉环境过程的基础。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










