



第二节 环境中的物质循环
一、水循环
地球上的天然水约1.4×1021kg,并具有和地球几乎相同的年龄(约46亿年)。这么多的水又这样长久存在而不消散,这和水的性质、大气的化学组成,地球的质量及地球在太阳系中的位置(和太阳间距离适中)等因素相关。
在地球表面,海水约占97%,冰川水约2%,淡水略小于1%,而大气中水分远小于1%。
水在地球表层的循环,其驱动力是太阳能。组成循环的主要过程有蒸发、蒸腾、降雨与径流,多数属于相变过程。地球表面水经吸收太阳辐射热而蒸发,进入大气的水中有83%来自海洋,但通过雨雪而返回海洋的水量仅为70%左右;陆地水的情况正好和海水相反,也就是说,有一部分海水是通过河川归流入海而得到补足的。在河川水归海的过程中还有一部分渗入土壤或岩层,为植物吸收或转入地下水,缓慢流动后才归流海洋。
生物圈含水总量少得几乎不能和其他圈层相比,但构成生物体的主要成分还是水。例如活细胞中含水分约60%~90%,其他生物体含水量大约是:脊椎动物为66%、哺乳动物平均为85%、木材为60%、干的种子为10%。氧、碳、氢依次是生物圈中丰度最大的三元素,且氧与氢的比例接近于水中该两元素之比。如此看来,生物体中含有如此多量的水是可以理解的了。
水在生物圈内的循环,在许多方面具有重大意义。首先,一切有机体内含有大量水分,没有水就不可能有生命。
还可以认为,地球表层是由太阳与大海造就的一个超级规模的蒸馏水制造装置,由此制得了淡水,供大多数生物需用。生命有机体中水的主要功能是作为输送与流通体内物质(如食物、氧气、排泄物)的溶剂。此外,水有很大热容值,能帮助高等动物身体保持平均体温。水还是参与光合作用与呼吸作用的重要物质。
流动于大气中的水分有调节气温的巨大能力。大气中一般含1%~3%(体积)水蒸气,这部分水分子能强烈吸收红外线,由此影响地球热平衡;大气中水分形成云层后还有反射太阳光而降温的作用;到了晚间,覆盖在地球表层的水蒸气又起了“被褥”的作用,使地面在白天所吸收到的热量不致于大量散发到空间中去。
由于水具有溶解许多物质及具有流动的特性,因此通过其本身循环能载带与推动其他物质循环。岩石中的很多组分受水体侵蚀后随水流归入大海是最显而易见的例子;再如,海水中所溶解的盐分能以喷沫形式转入大气,从而使近海地区降雨中含有相当数量盐分也是一例。
二、碳循环
1.碳在自然环境中的循环
碳在地球各圈层中的浓度如下:
地球整体:350mg/kg
地壳:200mg/kg
海洋:无机碳为28mg/kg;有机碳为2mg/kg
大气:CO2为3.60×10-4(mg/kg);
甲烷为1.7×10-6(mg/kg)
环境中碳(特别是含碳有机物)的存在形态十分多歧,这和碳元素的如下性质有关:①碳元素具有四个共价性较强的价键,可通过各种各样的结合方式联结大量基团;②C—C键具有很大强度,可形成十分长的长链结构;③碳原子有形成双键与多重键的能力,更加增大了生成多种有机分子的可能性。
地球上的碳主要集中在岩石之中,石灰岩中的碳(约占岩石层中总碳的3/4)主要以石灰石(CaCO3)与白云石(CaCO3·MgCO3)形态存在;沉积性页岩中的碳(约占岩石层中总碳的1/4)主要以分散性有机物形态存在。散布在其他圈层(水体、大气、陆生生物、土壤腐植质、化石燃料、海生生物等)中的碳大约只占地球上总碳的0.1%。
大气中碳的主要存在形态有二氧化碳、一氧化碳、甲烷等。在地球形成初期,作为大气组分之一的甲烷,随同氢、氨与水蒸气,通过发生在大气中的辐射或放电作用,导致了氨基酸等生命原始物质的形成。二氧化碳则逐渐被海水吸收,并以碳酸钙形式沉积海底,达成了现今二氧化碳在大气-海水-沉积物之间的平衡状态。
天然水系中碳的主要存在形态有构成平衡碳酸系统的各种组分:水合二氧化碳、碳酸、碳酸氢根离子、碳酸根离子。在水质测定中,这些形态的浓度之和组成了水样的总无机碳(TIC)参数。存在于水体中的有机碳组分可划分为溶解性有机碳(DOC)与颗粒有机碳(POC)两类。一般以含碳组分能否通过0.45μm滤膜作为区分此二者的判据。水体中DOC来源主要是浮游生物排泄物中可溶成分与生物尸体细胞经过胞溶作用产生的溶出物。水中DOC可进一步作为异养微生物的营养物料,在海水中还有部分可转化为高分子腐植质。严格说来,在作水样中总有机碳(TOC)项目测定前,应先过滤除去POC组分,随之测得的结果实际上就是DOC。构成水样POC的有难分解与易分解的两部分。前者为生物尸体的骨骼碎屑,后者如生物粪便物等为是。
碳是组成生物体的主要元素,所以碳循环,特别是二氧化碳的循环对生物体有极大意义。环境中二氧化碳的行为至少和下列一些环境过程有着密切的关联:①大气与海洋的起源与演化;②地球表层岩石的形成与变异;③生命诞生;④地球表层热量的收支状况;⑤水体中碳酸平衡;⑥绿色植物等自养者的光合作用与生产力;⑦有机物氧化,包括呼吸作用,燃料燃烧、机体腐烂分解等。对这些过程中于当今环境有现实意义者,将在以后相关章节详加阐述。
2.生物体中的含碳有机化合物
前已述及,原生质是所有细胞的组成成分,也列举了原生质的化学元素组成。
以上所列举的各种化合物在细胞中的含量不同。一般情况下,这些化合物占细胞鲜重的比例是:水大约占60%~90%,无机盐约1%~1.5%、蛋白质7%~10%、脂类1%~2%,碳水化合物与其他有机物1%~1.5%。这些化合物在细胞中存在的形式与所具有的功能也各不相同。
构成生物体的有机分子实际上是通过小分子缩合反应交联而成的。在大量小分子存在下,就有可能合成诸如淀粉、纤维素、蛋白质、核酸等大分子物质。与此相反的过程是水解,即在水与催化剂存在条件下,这些大分子裂解成为各种小分子的过程。
现将生物体中几种主要的含碳有机化合物分述如下。
(1)碳水化合物
这里所说的碳水化合物即糖类,可用Cx(H2O)y表示这类化合物的分子通式。糖类可分为单糖、二糖与多糖三大类。从化学结构看,碳水化合物是多羟基醛或多羟基酮,或者是通过水解能生成多羟基醛(或酮)的化合物。多羟基醛又称醛糖,多羟基酮又称酮糖,糖类广泛地分布在动植物的体内。
单糖一般是能溶于水的无色结晶,是一类不能进一步水解成更简单的多羟基醛(或酮)的碳水化合物。按照其分子中含碳原子的数目又可分为三碳糖、四碳糖、五碳糖、六碳糖与七碳糖。在动植物的细胞中,最重要的单糖是五碳糖与六碳糖。核糖与脱氧核糖是五碳糖,它们都是组成核酸的必要物质。葡萄糖与果糖是六碳糖,它们的分子式是C6H12O6。葡萄糖是植物光合作用的产物,也是机体内的重要能量物质。
二糖一般也是易溶于水的晶体。在植物细胞中最重要的二糖是蔗糖与麦芽糖;在动物细胞中最重要的二糖是乳糖。
多糖在植物细胞里,最重要的多糖是淀粉、纤维素与半纤维素,动物细胞中最重要的多糖是糖元。淀粉的分子量范围在2万~100万,是植物细胞中储藏能量的物质,在各类粮食中富含淀粉。纤维素是地球上最丰富的一种有机化合物,分子量范围在30万~50万。植物的细胞壁几乎全部由纤维素构成,借以支撑植物本体。由于它具有复杂的分子结构与不可溶解的性质,所以不能直接作为生物的食料。半纤维素的基本组成单元有六碳糖与五碳糖,大多数天然木材中除含纤维素外,还含相当多半纤维素。糖元的分子量更大于淀粉,有更多支链,它在肝脏与肌肉中含量较多,是动物细胞中储能物质。
二糖与多糖的最重要的相关性质是它们能通过生物酶作用水解为单糖。所有二糖与多糖都能水解产生葡萄糖。
(2)脂类
对脂类难以下一个确切的定义,简单说来,它们是动植物组织中能溶解于低极性溶剂(氯仿、四氯化碳、乙醚、苯等)的组分。在很多脂类物质中,除碳、氢、氧三种元素外,还含氮、磷等元素。脂类主要包括脂肪、类脂与固醇。
脂肪是生物体内储能物质。动物与人体内脂肪还有减少身体热量散失、保持体温的作用。脂肪在室温下呈固态,纯脂肪是无色、无嗅、无味的物质,大多是丙三醇与脂肪酸相结合的酯类物质。
类脂包括磷脂与糖脂。磷脂由甘油、脂肪酸、磷酸与一种含氮化合物总合衍生而成,是组成生物膜(细胞膜、内质网膜、线粒体膜等)的主要成分。糖脂的组成和磷脂相似,它是动物细胞膜重要成分。
固醇主要包括胆固醇、性激素、肾上腺皮质激素与维生素D等,这些物质对于生物体保持正常的新陈代谢起着积极的作用。
脂类的重要化学性质有水解作用与加成作用。由于这类化合物分子中多含有不饱和双键,所以在水溶液中可能发生卤素加成作用。但因脂类大多难溶于水,所以在一般化学反应条件下,加成反应的速度很慢。
(3)蛋白质
蛋白质存在于所有机体之中,在细胞中的含量仅次于水,约占细胞干重的50%以上。蛋白质的种类多、结构复杂,但每种蛋白质都含有碳、氢、氧、氮四种元素,许多蛋白质还常常含有少量的硫,有的还含有磷、铁等元素。蛋白质的元素百分组成如下:C(51%~55%),H(6.5%~7.3%),O(20%~24%),N(15%~18%),S(0.0%~2.5%),P(0.0%~1.0%)。
蛋白质由几千甚至几十万个原子构成,分子量从几万一直到几百万以上。氨基酸是蛋白质的基本组成单位,约有20多种。实际上,每个蛋白质分子就是由不同种类的、上百成千的氨基酸按照一定的排列次序连接而成的长链高分子化合物。每种氨基酸分子至少都含有一个氨基与一个羧基,当形成蛋白质分子时,一个氨基酸分子的羧基与另一个氨基酸分子的氨基相连接,发生缩合作用而脱去一个水分子,从而形成了联结两个小分子的肽键。由甘氨酸与半胱氨酸通过肽键联结生成高分子蛋白质的过程如下所示。

蛋白质分子结构的多样性,决定了蛋白质分子具有多种重要功能。作为结构材料,由蛋白质分子构成许多非骨骼性的机体物质(肌肉、皮肤、毛发等);作为催化剂,蛋白质分子可起生物酶的作用;作为激素,能在生物体内起调节代谢过程的作用;作为抗生物质,又能抵御外来有毒物质与病菌的侵入。
(4)酶与辅酶
酶是存在于活细胞中的,分子中含有金属原子(锌、镁、锰、铁、钼、铜之类)且分子量可达10000~50000的蛋白质或其衍生物。酶在活细胞中通过和基质形成不稳定中间化合物来降低系统活化能,所以是一类能加速代谢反应的生物催化剂。这种催化作用具有效率高、速度快、专一性强等特点,往往在一个细胞中可能同时发生由不同酶控制的数千种催化反应。
首先,酶在其活性部位和基质相结合,生成ES复合物。复合物可进一步分解,得到新产物P与复原的E。ES也可能反向分解,回复为E与S。在以上各个过程中,酶的化学结构始终都不变,表现出它的催化剂本性。
控制酶反应速度的因素有基质浓度、温度、pH值与离子强度等。在有作为抑制剂的毒物存在的情况下,可能使酶失去活性。
温度也是影响生物化学反应的重要因素。一般说来,在限定的温度范围内,温度每升高10℃,反应速率约增加2~4倍。即两者关系遵循范特霍夫经验规则。影响速率另一重要因素是pH值。有些酶在低pH值能发挥最大效能,而另一些酶则需要高pH值,但大多数酶在中性溶液中最有效。此外,某一种酶能起有效作用的pH范围是十分狭窄的。
酶的分子一般是由单独存在时不具有活性的两部分所组成:①不耐热的蛋白质部分,称为载体或酶蛋白;②称为辅酶的非蛋白质部分。辅酶具有较小分子量,热稳定性与膜透过性。酶与辅酶只有同时存在、同时作用时,才能体现催化活性。
酶的种类十分繁多,但按作用部位可将酶分为外酶与内酶两类,它们分别在细胞外与细胞内起作用。按作用特性,又可粗分为能催化水解反应的水解酶与非水解性的碳链裂解酶(或呼吸酶)两大类。一般说来前者属于外酶而后者属于内酶。
已知的辅酶约有12种,其中最主要的是以下几种:
NAD烟酰胺腺嘌呤二核甙酸的略名,以前称二磷酸吡啶核甙酸(DPN),也称辅酶Ⅰ。它是一种载氢体,常和脱氢酶、还原酶、过氧化物酶配合反应。NADH是它的还原形态,二者间通过氢传递过程而发生转化的反应如下:
NADP烟酰胺腺嘌呤二核甙酸磷酸的略名,以前称三磷酸吡啶核甙酸(TPN),也称辅酶Ⅱ。它的结构与功能和NAD相似,只是在分子上含有三个磷原子而不是两个。还原形态是NADpH。
辅酶A这是一种泛酸(B族维生素的一种)的衍生物,简写为CoA或CoASH。在脂肪酸的代谢与合成中,它能作为乙酰基载体参与三羧酸循环过程。
黄素蛋白包括黄素单核甙酸(FMN)与黄素腺嘌呤二核甙酸(FAD),它们在将代谢物中的氢(或电子)传递给氧的过程中起着重要作用。
辅酶M与辅酶F420属甲烷菌专有。在甲烷发酵过程中,两者分别起甲基载体与电子载体的作用。
(5)核酸
核酸最初是从细胞核中提取出来的,呈酸性,由此而得名。核酸由C、H、O、N、P等元素组成,是细胞中的另一类高分子化合物,分子量可达十万至几百万。核酸的基本组成单位是核苷酸。一个核苷酸分子由一分子含氮的芳香族碱基,一分子五碳糖与几分子磷酸所组成。每个核酸分子是由几百个到几千个核苷酸互相连接而成的长链分子。
三磷酸腺苷(ATP)与二磷酸腺苷(ADP)构成了细胞中主要的能量转移系统,它们都是核苷酸单体。通过水解,ATP脱去一个磷酸分子就形成ADP,同时放出能量。
相反的过程是ADP获得能量发生磷酸化反应,即和磷酸基结合成ATP。所得到的能量就以ATP形式贮存起来。
核酸可分为两大类:一类是含脱氧核糖的称为脱氧核糖核酸,简称DNA;另一类是含核糖的,称为核糖核酸,简称RNA。DNA主要存在于细胞核内,是细胞中的遗传物质。此外,在线粒体与叶绿体中,也含少量DNA。RNA主要存在于细胞质中。核酸对生物体的遗传性、变异性及蛋白质的生物合成具有极其重要的作用。
对于细胞作分子级的结构研究,按其深刻程度,可有三个层次。以上所述的基本上还只是第一层次,即弄清单体(单糖、氨基酸、核苷酸)是如何形成聚合链的。第二个认识层次是深入了解聚合链是如何进一步通过卷合或叠合成为更加复杂结构分子的(例如蛋白质中某些氨基酸组分中S—S原子间的键合情况)。第三个认识层次应能描述某些聚合物是如何再度集聚成三度空间结构分子的,这种集聚后的分子应具有螺旋式的结构。
3.光合作用
(1)进行光合作用的生产者
光合作用是生物圈内所发生的碳循环中的一个重要过程,对于生物体有着特别重要的意义。
生物体需要不断地从环境中获取碳源与能源。光合作用能使植物等自养生物在接受太阳光能的同时,将取自环境中的二氧化碳、水与营养物质合成为有机的含碳物质,从而也就将太阳能以该有机物所含的化学能形式贮存起来,以备自身活动、生长之需。此外,在推动碳、氢、氧、硫、氮、磷等元素的自然循环与生物循环中,光合作用也具有相当重要的意义。
地球上的绿色植物制造有机物与释放氧的规模是很大的。据估计,地球上的绿色植物每年能结合来自二氧化碳中的碳1500亿吨与来自水中的氢250亿吨,并释放出4000亿吨氧气。这个巨大的作业,只有10%是陆地上田野森林中的植物完成的,其余的90%应该归功于海洋里的单细胞植物与藻类。
人们对光合作用的机制尚有许多不清楚之处。
在标准条件下(25℃与1.013×105Pa),生产1mol的葡萄糖需要输入2880kJ的能量。按生态学术语说,那些能进行光合作用的绿色植物、藻类等都是生产者或称为自养者。光合细菌是另一类自养者,如绿硫细菌、紫硫细菌类能利用光能与二氧化碳保持自养生活。光合细菌不直接利用水,而是利用某些无机物(如硫化氢)或有机物(如醇、脂肪酸),按和光合作用相似的反应过程参与二氧化碳还原,且在反应产物中不含氧气。再有一类所谓化能自养生物,如生活在水或土壤中的硝化细菌、硫氧化细菌、氢细菌、铁细菌等,体内没有光合色素,所以不能利用光能,而是依靠生活环境介质中发生的化学反应所释出的化学能来促成碳水化合物的合成反应。
(2)光合作用的参与物
如前列方程式所示,参与光合作用的有自养生物外界环境中的光能、二氧化碳、水与某些无机营养物质。此外,还有自养生物细胞内部的光合色素、ATP——ADP、NADP——NADpH等。
对于植物来说,参与光合作用的光合色素是植物体内的叶绿素与类胡罗卜素。在植物体细胞里有一些被称为叶绿体的物质,叶绿体内部有许多伸长在壁与壁之间的称为“板层”的薄膜,板层的厚实之处称为“基粒”,而基粒中的每一个板层就像澡塘地面上铺得整整齐齐的瓷砖,它们都是可以进行光合作用的单元,里面含有250~300个叶绿素分子,光合作用主要就是通过这些叶绿素分子进行的。叶绿体中还含有另一类光合色素,称类胡萝卜素,包括胡萝卜素与叶黄素。
叶绿素,包括叶绿素a与叶绿素b,在高等植物中,它们大多浓集在绿组织细胞中。叶绿素是一类含镁的卟啉衍生物,具有卟啉的环状结构(这种环状结构存在于许多重要的生物物质之中,如血红蛋白、维生素B12等),这种环又是由四个含N的吡咯小环构成。各个吡咯小环的侧链上带有不同基团,而Mg离子则居于大环的中心。叶绿素a与叶绿素b在分子结构上的差异仅在于图中X位置上具有不同的基团。在叶绿素分子结构中,有着共轭双键系统,这对分子接受外来能量,从而引起键上电子激发是一个有利的因素。叶绿素a能吸收光能进行光反应。叶绿素b则将其吸收的光能传递给叶绿素a。类胡萝卜素除具有叶绿素b相似功能外,还能保护叶绿素a免被光氧化。现在一般认为,参与光合作用初始反应的只是位于反应中心的一小部分叶绿素a分子,其余的叶绿素a分子与其他的光合色素主要是吸收光能,并将之传递到反应中心的叶绿素a分子上。
(3)光合作用的机制
光合作用的反应机制十分复杂,其中包含着100多个反应步骤。根据已经了解的情况来看,基本上可将光合作用的过程分为光反应与暗反应两个阶段。光反应需要光合色素作媒介,将光能转化为化学能,其中包括水的光氧化分解反应与光合磷酸化作用。暗反应则主要是一些酶促反应,包括二氧化碳的固定与还原反应。
①光反应
在光合作用过程中,叶绿素吸收太阳光谱中可见光部分,并进一步引起各种光化学反应的情况。太阳光能的作用包括如下三个方面:
在光合系统Ⅱ中按下列反应式将水分解为氧气、质子与电子:

光合作用所产生的氧气就是来源于水的光化学分解;所产生的质子经由NADP-NADpH系统载带后去参与下一阶段暗反应中的化学反应,而电子将进入叶绿素分子之中,参与电子迁移反应。
在光合系统Ⅱ中,叶绿素a分子吸收小于680nm波长的光量子;在光合系统Ⅰ中,则是吸收700nm的光量子。由此,叶绿素a分子中某些基态电子受到激发,并进一步发生电离,其本身转为分子离子。从系统Ⅱ内叶绿素分子中逸出的电子由通过水分子原先分解产生的电子予以补充,此时叶绿素分子离子又复原成为分子。
引起光合磷酸化作用。由于光照引起的电子传递作用和磷酸化作用相偶联而生成ATP的过程就是光合磷酸化作用。即从光合系统Ⅱ中的叶绿素分子激发出来的高能电子在其迁移过程中将能量传递给ADP-ATP系统,从而将由光能转化过来的化学能贮存在ATP分子之中。释能之后的电子进入光合系统Ⅰ中的叶绿素分子离子,以补给该分子受700nm阳光激发后逸出的电子;后者又通过电子迁移归入NADP-NADpH系统,去参与下一阶段暗反应中的化学反应。由光合细菌发生的光合作用情况和上述略有不同。光合磷酸化作用是环式的,即细菌光合色素分子受光激发后,逸出的电子沿铁氧化还原蛋白所组成的光能链传递,中途释放能量以供磷酸化反应(→─ADP ATP)之用,最后又返回到光合色素分子的基态位置。
还应指出,在光合细菌中只有光合系统Ⅰ,没有光合系统Ⅱ;植物进行光合作用时,以水作还原剂,并放出氧,而细菌则用其他无机物(氢气、硫化氢、硫、硫代硫酸根离子等)或简单有机物(如琥珀酸)作还原剂,不产生氧。
②暗反应
暗反应可以在黑暗中进行,但需要多种酶参加催化才能正常进行。
绿叶从外界吸收到的二氧化碳,不能直接转化为有机物。在暗反应阶段,二氧化碳首先和植物体内的一种五碳化合物二磷酸核酮糖相结合,形成一种六碳化合物,这个过程就叫做二氧化碳的固定。所生成的六碳化合物很快水解裂成两半,形成三碳化合物磷酸甘油酸。在接受了由ATP与NADpH转来的能量与质子H+、电子e-后,磷酸甘油酸又转化为磷酸甘油醛,并释出作为暗反应另一产物的水。一分子磷酸甘油醛经过酶异构化而生成二羟丙酮磷酸酯后,与另一个分子磷酸甘油醛两相结合,就产生六碳化合物1,6-二磷酸己糖。多余的磷酸甘油醛又转化为暗反应的起始物(五碳化合物),由此形成了一个暗反应循环。
三、氧循环
1.氧在自然环境中的循环
氧在各地球圈层中的浓度如下(括号内的数字为其丰度排序):
地球整体:28.5%(2)
地壳:46.6%(1)
海洋:总量85.8%(1)
溶解氧量:15℃时为6mg/kg(13)
大气:23.2%(2)
所有元素中,唯有氧是同时在地壳、大气、水圈与生物圈中都有着极大丰度的元素。因此,在生物界与非生物界,元素氧都有着极端重要的地位。
按照化合物形态,氧的列于首位的无机氧主要是指岩石硅酸盐中所含氧。
在地壳中,形成岩石的矿物质中约95%是硅酸盐,其主要结构单元是四面体的 。其余5%的组分也大多含有氧元素,如石灰岩中碳酸盐(碳酸根离子)、蒸发岩中硫酸盐(硫酸根离子)、磷酸盐岩石中的磷酸盐(磷酸根离子)等。氧的离子半径是140pm(1pm=10-12 m),除钙(100pm)、钠(102pm)、钾(138pm)外,地壳岩石中其他主要元素的离子半径都小于80pm。正因为氧具有特别大的离子半径,所以以体积计的地壳元素组成中,氧占了极大的比例。当硅酸根离子这类含氧基团在岩石发生风化碎裂时,通常仍能以不变的原形进入地球化学循环,即随水流迁移到海洋,进入海底沉积物,甚至重新返回陆地。因此,地壳中存在的氧可看成是化学惰性的。
。其余5%的组分也大多含有氧元素,如石灰岩中碳酸盐(碳酸根离子)、蒸发岩中硫酸盐(硫酸根离子)、磷酸盐岩石中的磷酸盐(磷酸根离子)等。氧的离子半径是140pm(1pm=10-12 m),除钙(100pm)、钠(102pm)、钾(138pm)外,地壳岩石中其他主要元素的离子半径都小于80pm。正因为氧具有特别大的离子半径,所以以体积计的地壳元素组成中,氧占了极大的比例。当硅酸根离子这类含氧基团在岩石发生风化碎裂时,通常仍能以不变的原形进入地球化学循环,即随水流迁移到海洋,进入海底沉积物,甚至重新返回陆地。因此,地壳中存在的氧可看成是化学惰性的。
大气中的氧主要以双原子分子O2形态存在,并且表现出很强的化学活性。这种化学活性足以影响能和氧生成各种化合物的其他元素(如碳、氢、氮、硫、铁等)的地球化学循环。大气中的氧气多数来源于光合作用,还有少量系产生于高层大气中水分子和太阳紫外线之间的光致离解作用。
在此反应中同时产生氢气逸散到大气空间。
在紫外光作用下,大气中氧能转变为三原子分子臭氧。第一步是氧分子通过光解反应生成氧原子:

随后,氧原子与氧分子结合生成臭氧分子:
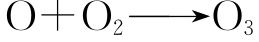
通过以上反应,在距地面约10~40km的大气层上空形成了臭氧层,正常情况下,臭氧分子的形成过程与随后的分解过程在臭氧层中达到平衡,所以,臭氧层中的臭氧具有大体恒定的浓度;又由于臭氧的生成与分解都需要吸收紫外光,所以臭氧层成为地球上各种生物抵御来自太阳过强紫外光辐射的天然屏障。臭氧层对于地球生物,有着生死攸关的作用。
在构成水圈的大量水中,氧是主要组成元素;在水体中还有各种形式的大量含氧阴离子以及相当数量的溶解氧,它们无不对水圈或整个生物圈中的生物有着极为重要的意义。
各种含氧化合物在氧循环中发生迁移与转化的情况。在转化的各种过程中,许多别的元素也随同氧元素一起进行着循环。
在生物光合作用与呼吸作用的过程中,参与氧循环的物质有二氧化碳、水等。化石燃料的燃烧与有机物腐烂分解过程则是和呼吸作用具有类似情况的一类氧化反应。对于呼吸作用与燃烧过程将分别在下一节与以后的相关章节中予以详述。
由于火山爆发或有机体腐烂产生硫化氢,能在大气中进一步被氧化为含氧化合物二氧化硫,化石燃料燃烧及从含硫矿石中提取金属的过程中也都能产生二氧化硫,这些二氧化硫在大气中被氧化为硫酸根离子,然后通过酸雨形式返转地面。相似地,由微生物或人类活动产生的各种氮氧化合物最终也被氧化为硝酸根离子,然后通过酸雨形式返回地面。
大部分金属通过氧化过程转化为不溶性氧化物,也有一些还原性的非金属可能被氧化为溶解性更大的化合物,这种情况大多发生在陆地上,也少量地发生在海洋中。如:

大气中的氧与水体中的溶解氧之间存在着溶解平衡关系。当由于某种外来原因引起平衡破坏时,该水-气体系还具有一定的自动调节、恢复平衡的功能。例如当水体受有机物污染后,水体中的细菌当即降解有机物并耗用水中溶解氧,被消耗的溶解氧就由大气中的氧通过气-水界面予以补给。
反之,当大气中氧的平衡浓度由于某种原因(例如岩石风化加剧)低于正常浓度时,则水体中溶解氧浓度也相应低落。由此,水体中有机物耗氧降解作用缓慢下来,相反地促进了水生生物的光合作用(增氧过程),这样就会进一步引起表面水中溶解氧浓度逐渐提高到呈过饱和状态而逸散到大气中去。
2.呼吸作用
(1)意义
生命的一个重要特征是它和环境的物理过程正好相反,环境趋向于从高度有序状态逐渐变为无序状态,而生命以高度有序状态为特征。它在产生、生长、壮大、死亡的过程中力图保持这种高度的有序性。因此,生命的连续存在取决于能量不断供应,以修复环境所引起的破坏。尽管能量的最根本来源是太阳的辐射能,但对于生物,能量要在呼吸过程中才能成为可供利用的形式。
呼吸作用是生物体参与氧循环的一个重要过程,和光合作用把简单的无机物合成为复杂的有机物并且储藏能量的过程相反,呼吸作用是分解复杂的有机物,并且释放出能量。但是,不能把呼吸作用看成是光合作用的简单逆转。事实上,这两种作用都是生物体内生命活动中的复杂过程。
呼吸作用在生物的生命活动中有三方面意义:①为生物的生命活动提供能量;②在呼吸作用中产生各种中间产物,可作为合成核酸、蛋白质、酶、色素与生物激素等必须物质的原料,因此呼吸作用可以说是生物体内各种有机物相互转换的枢纽;③能增强生物抗病能力。当有毒物质进入生物体时,生物通过强烈的呼吸作用,可以氧化分解这些有毒物质。呼吸作用对于环境的重要意义在于:各类呼吸过程是碳、氧、氮、硫等元素参与生物地球化学循环的重要环节;细菌呼吸关系到水体自净,污水治理等过程。
生物的呼吸有两种类型:好氧呼吸与厌氧呼吸,这两种类型的呼吸各有不同的过程。陆栖动物靠空气中的氧气进行呼吸;水栖动物则通常摄取水中的溶解氧,它们都是好氧呼吸的生物。还有少数厌氧生物须要在没有氧的地方才能生存,如昆虫的幼虫、软体动物,贫毛动物、原生动物及厌氧性细菌皆在此列。
此外,兼气性细菌在有氧或无氧条件下都可生活。也还有这样的情况,即好氧呼吸的生物,在特定条件下可以进行厌氧或欠氧呼吸,这是生物体适应环境的体现。在此我们将先着重介绍好氧呼吸过程。
关于好氧呼吸作用的最一般概念是:生物体从环境摄取氧气,然后由体液将氧气输送给组织并为细胞所利用,最终再由组织除去二氧化碳的物理与化学的过程。植物、动物、微生物体内所进行呼吸作用的过程各有其异同之处。
(2)线粒体及其中的催化物质
普通的线粒体像个橄榄球,是细胞在有氧条件下进行呼吸的主要场所。一个普通细胞可能含有几百到一千个线粒体,很大的细胞可含有几十万个,而厌氧菌里一个也没有。
线粒体内含有多种酶、辅酶与细胞色素,这些物质能催化在线粒体内进行着呼吸作用的三个主要过程,即三羧酸循环、电子传递与氧化磷酸化作用。线粒体具有很精致的结构,它由一层内膜与一层外膜构成,内膜表面充分扩展,折成很多皱褶或嵴,以扩增它的面积。这些嵴陷入整个线粒体的内部,嵴和嵴之间,内膜和外膜之间是结构不定的物质,称为基质。嵴的表面上有很多小球体,可能就是能催化生物电子传递与磷酸化作用的酶分子复合体。在外膜上松散地连接着三羧酸循环过程所需的酶与很多能催化辅助反应的酶。
呼吸酶是内酶,主要包括对氧化还原反应有催化能力的氧化酶、脱氢酶、过氧化氢酶以及能在基团迁移反应中起催化作用的氨基转移酶、磷酸基转移酶等。
此外,在线粒体内膜上还附有各种细胞色素,它们是以铁卟啉复合体为辅基的血色素蛋白。细胞色素具有电子载体的功能,这种功能是借助于其中铁离子能经受可逆的价态变化来实现的。细胞色素主要有四种,定名为a、b、c与d。在生物氧化链索中占最重要地位的是细胞色素c,其辅基和蛋白部分肽链中两个半胱氨酸分子相连。细胞色素c是一种可溶物,氧化型为黄色,还原型为红色,耐热而不易变形,含铁约0.43%,分子量约13000。
生物氧化过程之所以需要多种催化剂交替作用来完成,主要是为了组成某种氧化还原电位梯度,反应逐级进行,可使过程顺利完成,放出大量能量。
3.好氧呼吸作用进行的过程
好氧呼吸作用就是在氧气参与下,将生物体内的能量物质氧化分解为最简单产物(二氧化碳与水)的过程。在这一过程中,同时放出一定能量。呼吸过程总的反应如下:
C6H12O6+6O2─→6CO2+6H2O+2880kJ
由上式可见,从外界环境吸入的氧参加了这个氧化分解过程,而放出的二氧化碳是过程的产物。在呼吸过程中,氧与二氧化碳的交换,是最明显与最易被研究的外部特征。但实际上整个呼吸作用是一个十分复杂的过程。
好氧呼吸作用(包括三羧酸循环、电子转移、氧化磷酸化作用)及其先期能量物质的消化作用,是保持好氧生物的生命所必须的两种功能。消化能量物质的功能很像一个代谢的“磨石”,能把大分子氧化成小分子并进一步发生代谢;如果生物不能对其有所利用,就将它们排泄掉。呼吸作用的功能恰如一个“发动机”,保持着细胞的活动,并保证细胞进行各种活动时能提供足够量的可利用的化学能。
能量物质指的就是淀粉、脂肪与蛋白质。以哺乳动物为例,在消化道内各种消化酶作用下,将能量物质消化分解为可以吸收的小分子有机物。淀粉分解为葡萄糖,脂肪分解为甘油与脂肪酸,蛋白质分解为氨基酸。在分解大分子化合物的同时,将它们所含的大量化学能释放出来,供生命活动利用。对细菌来说,则是通过向介质(如水)分泌外酶,将存在于介质中的能量物质水解为小分子有机物,然后摄入体内,再进行内呼吸作用。
糖类是上述三类有机物中的主要能量物质。下面就以葡萄糖为例,叙述该能量物质如何在呼吸过程中最后转化为二氧化碳与水这两种产物的。
4.葡萄糖酵解生成丙酮酸的过程
所谓酵解指的是在无氧条件下进行分解的过程,这还只能算是有氧呼吸之前的一个先行过程。葡萄糖酵解过程包括很多步骤,最后的产物是丙酮酸。丙酮酸再经过氧化,并进一步和辅酶A结合,生成作为呼吸作用第一个过程(三羧酸循环)的引发物乙酰辅酶A。
葡萄糖酵解过程计有4个步骤:
(1)酵解的第一步是葡萄糖的酯化,即葡萄糖在酶与ATP存在下转变为葡萄糖-6-磷酸酯。
(2)葡萄糖-6-磷酸酯受到酶的作用而转变成果糖-6-磷酸酯。

果糖-6-磷酸酯在ATP存在时,受酶的作用而形成果糖-1,6-二磷酸酯。
(3)果糖-1,6-二磷酸酯经过酶的催化作用而分裂成2个丙糖,即二羟丙酮磷酸酯及3-磷酸甘油醛。后二者互为异构体,且能相互转化。所以,为后续反应所需而逐渐消耗的3-磷甘油醛,可通过异构体转换反应不断得到补充。
(4)3-磷酸甘油醛又通过一系列的酶促反应最终生成丙酮酸。
葡萄糖酵解反应全过程可用下列方程总结:
C6H12O6─→2CH3COCOOH+4H
过程中所放出的净能量用于氧化磷酸化反应以合成ATP。
细胞中氧气充足条件下,丙酮酸可进一步氧化脱去羧基生成乙醛与二氧化碳,乙醛与辅酶A反应生成乙酰辅酶A,是三羧酸循环的引发物。
三羧酸循环又名柠檬酸循环。因为首先由德国学者Krebs阐明,所以也可称为克雷布斯循环。
由葡萄糖酵解产生的丙酮酸,一方面可生成乙酰辅酶A,另一方面也可能与二氧化碳作用产生草酰乙酸。乙酰辅酶A极为活泼,可和草酰乙酸缩合为柠檬酸,同时再生出辅酶A,从柠檬酸出发,经历许多种中间产物,完成循环后的最终产物又是草酰乙酸。
在三羧酸循环过程中:①丙酮酸、柠檬酸与草酰乙酸可在代谢程序上互相衔接,成为一种循环机构,这种循环即为三羧酸循环;②从丙酮酸开始,以草酰乙酸为终结,整个循环中放出3个分子二氧化碳,作为有氧呼吸的废弃物;③在整个过程中脱出的氢原子被NAD与FAD所接收,由此形成NADH、FADH2与H+;转入呼吸链去参加还原O2的反应;④在整个循环中所释出的能量,用于将ADP分子磷酸化为ATP分子。
氧化还原反应包括质子与电子迁移以及氧化磷酸化反应,由此组成了一个呼吸链。
有氧氧化时,有机物往往不直接将氢原子交给分子氧,而是通过一系列载氢体来传递。这些载氢体先后是NAD、FAD、CoQ(辅酶Q)。由CoQH2释出H+,同时又将电子传递给细胞色素,使电子再沿细胞色素循环往下传递。最后,质子、电子与氧分子汇合生成水分子,从而完成氧化反应。
5.厌氧呼吸
大多数动植物与微生物在有氧条件下进行好氧呼吸。但在欠氧或无氧条件下,这些生也能适应环境条件,相应地进行厌氧呼吸,例如人在激烈运动时的呼吸,根部入水的水稻兼氧微生物所进行的呼吸即是。此外,厌氧性微生物本来就需要在无氧条件下才能生活。
有机物厌氧分解过程有一定的限度,即不能最终达到无机化程度,这样的过程称为发酵。
糖类发酵的产物可能有酒精、乳酸与其他低碳一元羧酸。这类过程被称为酒精发酵与酸性发酵。过程实质是以葡萄糖酵解产物丙酮酸代替氧气作受氢体。过程中所产生的酸可使介质pH值降低,进一步引起细菌生命活动中断而停止降解过程。但如果条件适宜就可能发生另一类发酵过程,使有机醇、酸继续实行无氧氧化,最终产生甲烷气体,这称为甲烷发酵。
甲烷发酵是由专门的甲烷细菌促成的,这类细菌是绝对厌氧菌,在污水与污泥中广泛存在,可以分解低级的醇、酸以及醋酸。这些反应的实质是以二氧化碳或有机物分子本身作为受氢体的无氧氧化过程,可表示为:
2CH3CH2OH+CO2─→2CH3COOH+CH4
2CH3(CH2)2COOH+CO2+2H2O─→4CH3COOH+CH4
CH3COOH─→CO2+CH4
甲烷发酵是在无氧条件下有机物降解的最终阶段,但它只能在适宜的条件下发生。无氧、有一定的菌种、一定的pH值与温度范围等都是必要的条件。甲烷细菌要求的最优pH值为6.4~7.2,低于6或高于8则会停止生长繁殖。最优温度则在32℃~70℃。
以上各种发酵过程也是在各种酶与辅酶作用下完成的,过程具有相当的复杂性。由于发酵产物仍是小分子有机物,所以过程中释放出来的能量是不高的。如上述乙醇发酵与乳酸发酵时产生的能量分别是112.5kJ/mol与97.5kJ/mol(作为对比,葡萄糖彻底氧化释出能量为2880kJ/mol)。
总的说来,有机物厌氧性降解大多发生在富营养湖及污染河流底部,这种过程在不同种类的细菌作用下,按水解、酸化、甲烷化的三个阶段进行,前两阶段合称液化阶段,后一阶段称汽化阶段。
四、氮循环
1.氮在自然环境中的循环
氮在地球各圈层中的浓度如下(括号内数字为其丰度次序):
地壳:20mg/kg(31)
海洋:溶解氮气,15.5mg/kg(11)
可溶性氮化合物:0.7mg/kg(17)
大气:75.53%(1)
陆地、大气与水系中氮的存在形态分布,存在于陆地上的氮主要是分布在生物体内的有机氮,水系与大气中则都以分子氮为其主要存在形态。
大气中分子态氮不像氧气那样具有高度的化学反应性,因此就不可能直接参与生物体内的元素循环。对于大多数需氮而又欠氮的作物,需要得到外来的含氮肥料才有可能正常生长。
氮分子之所以有化学惰性,是因为构成它的两个氮原子以三个共价键维系在一起:N≡N。要形成新的氮化物则需要外加905kJ/mol的能量,以使这三个键都发生断裂。对很多微生物说,它们却有能力越过这样高的能垒,使形成氮化合物的过程得以进行。所以在大多数情况下,只要具备其他必须条件(如充足的水分),则自然环境中发生的氮循环就能以足够大的速率来推动植物生长。
各种氮化合物的相对热力学稳定性可用生成自由焓值来度量。在氮循环中,由生物物质引起的反应起了主导作用,这些反应发生的次序为:

相逆的过程,即返转到氮气的反应,也可以在微生物存在下顺利地发生。
一般环境条件下,即便是无机氮化合物也不可能通过大气中氮经过自发反应而生成。只是在高温条件下(例如天空打雷闪电,高温燃烧炉或汽车发动机内)才能生成各种氮氧化合物,它们在大气中又逐渐转变为硝酸盐,作为微生物生产硝酸盐的补充。
1914年,德国人哈柏发明了合成氨的方法,从此开创了大规模氮肥工业。据估计,世界范围内通过微生物作用全年固氮量约是9.0×107吨,而世界氮肥产量可能已达8.0×107吨。由此可见,氮肥生产对世界粮食的逐年增产有着决定性意义。由于氮肥的使用,使这类化合物逐年散失在环境中,参与氮元素的生物地球化学循环。
2.生物固氮作用
许多微生物与藻类都能将空气中的氮气还原成氨,这就是生物固氮作用。生物固氮对保持自然界的氮循环与供应植物生长所需要的氮源都起着重要作用。
生物在固氮过程中起作用的是其体内的固氮酶。这种酶是两种蛋白质(铁蛋白与铁钼蛋白)的复合体。
能固氮的生物大体上可分为共生固氮生物与自生固氮生物两大类。共生固氮生物(如根瘤菌)的特点是它们独立生活时,没有固氮作用,当它们侵入宿主植物后形成根瘤,从宿主植物汲取碳源与能源进行固氮作用,并供给宿主以氮源。
自生固氮生物是指能独立固定氮气的生物。它们能利用土壤中的有机物或通过光合作用来合成各种有机成分,然后耗用这些碳水化合物并将分子氮转变成氨态氮。自生固氮生物又包括好氧与厌氧的细菌、光能自养细菌、烃氧化细菌、蓝藻等。
生物体氧化碳水化合物时生成二氧化碳,同时释出电子。
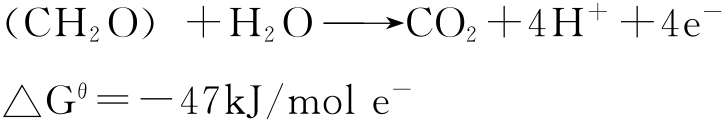
固氮反应正好需要让氮气获得电子,使N(0)还原到N(-3)。

在后一个半反应中,氮气每得到一摩尔电子需要27kJ能量,从能量角度看这个半反应难以发生,但如果以上两个半反应相结合,则可得到总自由焓变发生负变化的结果。
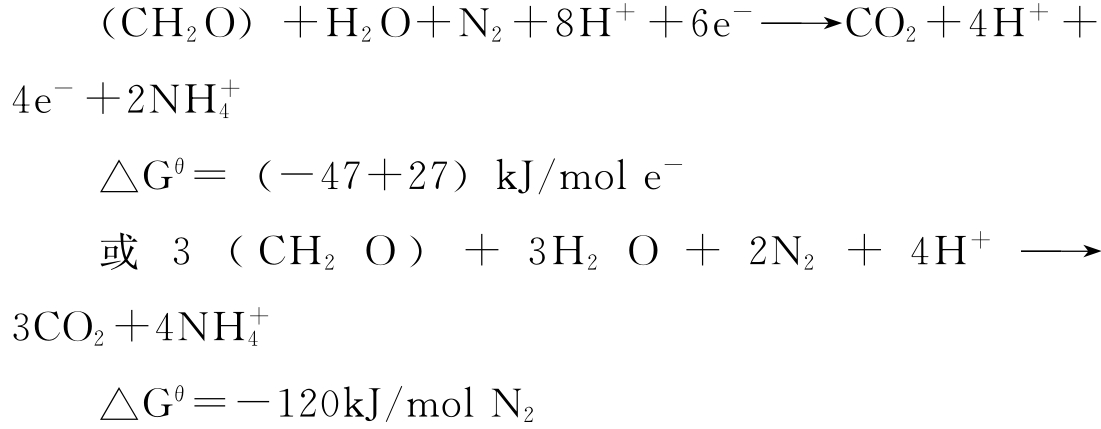
总的反应中释出能量,所以从热力学角度看,反应是容易进行的,微生物也就能由此获取能量。就这样,将氮循环与碳循环联系起来,在一个半循环中释出能量,生成较稳定的化合物;在另一个半循环中获取相当的能量,而生成不太稳定的化合物,整个循环能量得以平衡,而循环得以正常进行。
固氮酶中含有两个大的蛋白质分子。由于生物样品来源不同,人们所能获取的固氮酶其分子量及其中金属的含量也有差异。一般地说,铁钼蛋白中含2个钼原子,24~32个铁原子、24~30个不稳定的硫原子、分子量约220000;而铁蛋白中只含4个铁原子、4个不稳定的硫原子、不含钼原子,分子量约68000。这两种蛋白合并起来才有固氮作用,单独存在时没有活力,且已知,微量元素钼在固氮作用中具有关键性意义。氮和铁钼蛋白中的钼键合,还原并生成氨。
由此看来,氮气还原成氨气不是一个简单过程,而是需要一步一步地克服一个一个的小的能垒才能逐步到达终点。在过程中所需能量通过MgATP水解产生后,再传送到固氮酶:
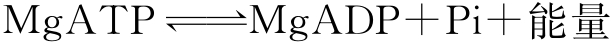
其中还发生电子转移,系通过Fe或Mo的氧化态变化而实现的

不管是固氮反应所需的能量也好,质子、电子也好,终究说来,它们都源于碳水化合物的氧化。从这一意义上看,我们在考虑问题时,应将碳、氢、氧、硫、磷、镁、铁、钼等元素在生物体内循环和氮元素的循环联贯于一体。此外,从以上叙述也可看出,很多微量元素在生物体中起有不可缺少的作用。
3.生物同化作用
由固氮作用产生的氨可在生物体内进一步转化为氨基酸与蛋白质。我们将这样的过程称作生物同化作用(广义地说,固氮也是一种同化过程)。
在水体中含有多种化学形态含氮化合物,其中氨成为水中藻类择先汲取的营养物,当氨进入其体内后,即按以上反应过程转化为蛋白质。所以水体中如含高浓度氨,将会促使藻类过度繁殖,以致发生“富营养”环境问题。
4.含氮有机物的降解
有机氮化合物降解的最终产物是氨,所以这个过程也被称作氨化作用(或无机化作用)。在自然环境中,这类作用大多属于有微生物介入的生物降解过程。
含氮有机物包括蛋白质、氨基酸、尿素、胺类、硝基化合物等等。它们的元素组成,除氮外还含有碳、氢、氧、硫、磷等。一般说来,含氮有机物在环境中生物降解较不含氮有机物更难,其产物污染性强;同时,它的降解产物和不含氮有机物的降解产物会发生相互作用,影响整个降解过程。
以上属水解反应,大多发生在微生物体外。氨基酸产物被微生物摄取后,在体内以脱氨、脱羧两种基本方式继续被降解,由此得到作为降解产物的氨。
高等动物尿中含有尿素,排入环境后在以尿素酶为媒体的反应中迅速水解生成碳酸氨。在偏碱性的介质中,此产物因稳定性不足而进一步分解,生成产物氨。
5.硝化作用
由固氮作用生成的氨或由蛋白质等高分子含氮化合物经过降解作用后产生的氨,都可能在有氧条件下,经过细菌作用而硝化硝化作用对土壤中的植物有特殊意义,因为一般植物容易吸收硝酸根离子形态的氮,不易吸收氨气或氨根离子形态的氮。但从另一方面看,硝酸盐有极大可溶性,不易为土壤所阻留,这又是对植物不利的。能引起硝化反应的细菌主要是亚硝酸菌类与硝酸菌类,合称硝化细菌。它们是无机营养型细菌,即化能自养菌,可利用硝化反应中释出的能量来合成自身所需的营养物质,或组建自身的细胞物质。
在硝化过程中,后一步反应的速度比前一步反应快得多,所以不会发生亚硝酸根离子在环境中大量累积的情况。此外,反应专一性很强,即两步反应必须由两类细菌分别承担,彼此不可替代。
硝化过程还具有很复杂的反应机理,氨氧化为亚硝酸根离子、硝酸根离子的过程中还有很多中间产物,如羟胺、硝酰基、一氧化二氮等。
6.反硝化作用
又称脱氮作用。在土壤中以及在水体的底泥或中间水层环境介质中都可能发生这种作用。兼气性反硝化细菌在欠氧条件下进行呼吸时,将其所汲取的能量物质施行脱氢氧化,这时因为周围环境中缺乏氧气,所以就以硝酸盐代替氧气作为受氢体,致使硝酸盐被逐步还原,这就是反硝化作用。
反硝化过程可简单地表示如下:
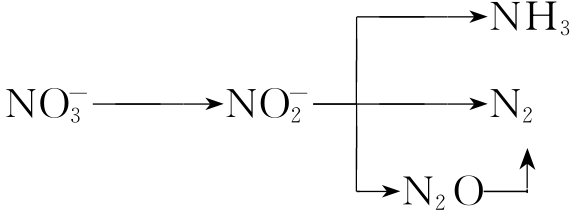
其中氮气与一氧化二氮是反硝化作用的主要产物,但一般情况下,在生成过量一氧化二氮条件下才能产生一氧化二氮。
葡萄糖在反硝化过程中被氧化的反应方程如下:

在这一反应中,硝酸根离子作为呼吸过程中的最终电子受体。当每摩尔葡萄糖完全氧化时,可产生的能量约2.51×103kJ,达到有氧氧化时的90%左右,因而是容易发生的。
海水中有机物在反硝化过程中被氧化的反应方程式如下:

反应式左侧的那个复杂结构化合物代表海中藻类、浮草等的化学组成。在反应式中还没有将反应中间产物亚硝酸根离子表示出来。此外,作为反应产物之一的氨气,还可能继续被氧化:

五、硫循环
硫在地球各圈层中的浓度如下(括号内数字为其丰度次序):
地球整体:1.8%~2.9%(5或7)
地壳:260mg/kg(16)
海洋:905mg/kg(6)
大气:0.6×10-9(V/V)(15)
地球上的硫可能大部分贮存在地幔与地核中,但人们对此详情知之甚少。近代,由于工业革命的推动,使燃料的用量大为增多,冶金工业与肥料工业迅速发展,由此促进了硫及其化合物在地球表面的生物地球化学循环。
将硫循环与氮循环作比较,两者间有很多相似之处,这显示在:在自然循环中,参与循环的生物介质中的硫与氮大多数是还原态化合物;微生物的作用很大程度上影响着这两种元素的生物地球化学循环进程;通过循环进入大气中的少量硫化合物与氮化合物,在某些情况下会引起很大的环境污染问题。这两个循环的主要相异处则在于:硫的主要储层是地壳,而氮的主要储层是大气。
在好氧条件下,存在于水体、土壤中的还原态硫化物可借助微生物作用氧化为高价态化合物。而在大气中,单靠化学因素也能实现同样过程。
在某些情况下,单质硫也可能不会生成,所以用括号括起,以示和其他生成物有所区别。
在无氧条件下,硫化氢可通过硫细菌进行光合作用而被氧化,这时的硫化氢相当于绿色植物光合作用中的水。
生物还具有将无机状态的硫(主要是水中溶解的硫酸盐)加工成为自身体内有机硫化物的能力。此时,需将硫酸根离子中的六价硫转化为RS—H中的二价硫,这是一个得到8个电子的生物还原过程,情况十分复杂,包含许多中间步骤。许多细菌、藻类与植物都可以完成这一过程。
在欠氧条件下,各种硫酸盐还原菌可以进行反硫化作用,将硫酸根离子作为受氢体并还原为硫化氢。因此,硫化氢成为污染的明显标志之一,它可以发生臭味,使水呈黑色。在同一环境介质中,一般需要前述反硝化作用完成,即硝酸盐耗尽后,才开始发生反硫化作用。在此写出海水中可能发生的一种反硫化过程反应式,以和反硝化过程式相比较。

以上反应在水中溶解氧浓度[DO]≤0.11mg/L与细菌参与下发生。
作为污染物的硫氧化物在影响硫的正常循环与影响大气质量方面有着非常重大的意义。
六、磷循环
磷在地球各圈层中的浓度如下(括号内数字为其丰度次序):
地球整体:0.2%(10)
地壳:1050mg/kg(11)
海洋:88μg/kg(19)
自然存在的磷化合物都具有很低的溶解度与挥发性,所以磷元素的生物地球化学循环的通量很小,且取决于河川中含磷悬浮颗粒物往海洋迁移的速率与含磷尘粒在大气中的迁移能力。
磷在自然界或生物体中几乎完全以无机磷酸盐或含磷酸根离子有机物形态存在,也就是磷的完全氧化态,价态为(+5)。
最常见的含磷的矿石主要有羟基磷灰石[Ca5(PO4)3(OH)]、磷酸钙[Ca3(PO4)2]、磷酸铝[AlPO4]、磷酸铁[FePO4]。磷酸盐矿的实际组成远比以上分子式所示的复杂,因为在矿石中的阳离子(钙离子、铝离子、亚铁离子、铁离子等)间普遍地发生同晶或类晶置换,而且矿石中还含有种类与数量不一的各种微量元素。
天然水中的磷是通过矿石风化侵蚀、淋溶、细菌的同化与异化作用等自然因素引入的。
作为人为来源,主要是含于城市污水中的合成洗涤剂含磷组分排入水体。和含氮肥料易从土壤流失进入水体的情况不同,土壤中磷肥的溶解度很小,经水流作用而迁移的能力也很小。溶解态磷中的正磷酸盐部分(磷酸根离子、磷酸氢根离子、磷酸二氢根离子)可作为营养物质被水中藻类多量摄取,所以这种形态的磷具有很大环境意义,且在正常水体中浓度很低(如前所述,这类盐的溶解度也是很小的)。聚合磷酸盐是合成洗涤剂组分之一,其作用是络合水中钙离子、镁离子等离子,使之不以碳酸盐形态沉积下来。
水体中聚合磷酸盐的形态主要有过磷酸根离子、三聚磷酸根离子、三聚磷酸氢根离子、过磷酸钙等。可溶性有机磷酸物主要有葡萄糖-6-磷酸、2-磷酸甘油酸、磷肌酸等形态。
在生物的体内,磷是组成能量转移分子ATP、ADP、AMP以及具有遗传等功能分子RNA、DNA的基本组分。这显示出磷是生命体的必须元素,也是土壤与水系内生态系统中的营养要素。
磷在环境中的整体循环可被划分为三个子循环,即一个无机循环与两个生物循环。无机循环的速度很慢,其周期以109年计。
火成岩受风化作用后,其中磷化合物迁移的主要途径是由岩石经过土壤、河流转入海洋。由于海水偏碱性且含多量钙离子,所以多数磷又以磷酸钙形式沉积于海底。
在深海沉积物中,这些磷酸盐几乎被永久封闭而不易复出。
只有通过人类采掘活动或海底鱼类的食用(再经过食物链)才有可能少量地重返陆地。但沉积在内陆海或大陆架中的磷酸盐则可通过地面隆起等地质过程再次成为新陆地的组成部分。
水体中磷的生物循环起始于藻类对水中可溶性正磷酸盐的汲取;陆地上磷的生物循环起始于植物根部从土壤水中摄取磷酸盐。这两种生物循环速度很快(分别以天计或以年计)相关方面研究工作的内容十分广泛,现今还处于较浅薄的认识阶段。
七、污染物的迁移与分布
1.污染源排污特点
为了建立起一个能描述污染物环境归宿的模型,需要具备相关污染物在环境中随空间与时间分布的各种数据与资料。在此我们先从污染源出发,阐述它们在空间与时间方面的排污特点。按空间特点可将污染源分为点源、线源、面源与区域源。
工厂烟囱是典型的点污染源,其特性可用地理位置、烟囱高度与排烟量等予以确定;工厂废水排污口也可看成点源。公路汽车排气与工厂区排气可分别视为线源与面源。区域源的例子较少,典型的是布满某一地区空间的汽车排气,可将它看成是二次污染物光化学烟雾的区域源。
按时间分布特点可将污染源分为连续的、间歇的、周期的、事故性的与无规则的几类。发电站烟囱排烟在时间方面有一定程度连续性与规则性;如敞开的炼焦炉、化工厂车间排气等则通常是间歇性的,但具有可预测的规则性。连续而又具有周期性的例子如汽车在一天中的排气;含污降水则往往是无规则的。
排污率一般用千克/秒(kg/s)(点源)、千克/秒·米(kg/s·m)(线源)、千克/秒·米2(kg/s·m2)(面源)表示。对由事故引发的一次性污染源的排污率通常要用千克/(一次事故)表示。
2.化学污染物在环境中的迁移
凡排放到环境中去的化学物质都将进入大气、土壤、水系或生物体内。离开排污源的化学物质可以发生化学形态转化或维持原有状态,它的转化产物或其本身可停留在排污源附近或迁移到别的相邻的圈层中去。只要化学物质对环境因素的作用有足够的对抗能力,它就可在整个生态系统的各环节间连续循环或相对牢固地滞留在某个环节上。
关于影响化学物质迁移过程的主要因素,有该化学物质的各种物理性质、化学性质以及环境介质的各种性质。这两方面的因素比较起来,一般前一类因素着重影响该物质在各个圈层间所发生的迁移,后一类因素则对其在某一圈层内的迁移具有相对重要意义。
3.化学污染物在环境中的分布
对包括迁移与分布在内的化学污染物的各种环境行为很难作出可靠的预测,而现场实测工作又难以控制条件且耗资巨大,所以一般是先提出一种迁移模型,然后通过实验室工作,对每个单一过程进行研究,并获得必要的参数,然后再反过来验证原先模型的准确性。
现今已提出过许多种可用于阐明化学污染物环境分布的模型。现以某非极性有机污染物为例,介绍一种处理问题的最简模型。建立这种模型仅需具备污染物在各圈层间的相间分配系数数据,而这些系数可通过实验测定或从污染物的基本物理化学性质参数予以估算。虽然这种模型过于简化,但却可作为进行动力学模型研究的先导性工作。
在解决污染物于各圈层分布问题时先提出如下几个假设:①每一圈层中的介质是均匀的,污染物在其中分布也是均匀的;②污染物一旦进入环境,即迅速迁移,随后达到分配平衡;③污染物在迁移过程中来不及发生降解,只是在达到分配平衡之后方始发生降解。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










