



第二节 生物催化剂——酶
一、酶的化学本质及组成
生物体内的代谢不是完全自发进行的,而是靠生物催化剂——酶来催化的。生物的生长发育、繁殖、遗传、运动、神经传导等生命活动都与酶的催化过程紧密相关,酶是推动生物体内全部代谢活动的工具。那么酶到底是什么物质呢?十九世纪后,随着对酿酒中发酵过程的深入研究,科学家才逐步揭开了酶的“面纱”。
19世纪,酿酒业在欧洲经济中占有重要地位,科学界也非常重视对发酵过程的研究。1857年,法国微生物学家巴斯德提出酿酒中的发酵与活的酵母细胞不可分割,没有活细胞的参与,糖类是不可能变成酒精的。德国化学家李比希却认为发酵是酵母中的某些化学物质作用的结果,与活细胞无关。两种观点争执不下。
1897年德国化学家毕希纳结束了这场争论。他制备了不含细胞的酵母提取液,然后在这些汁液中加入葡萄糖,一段时间后糖液居然变成了酒。这一结果说明发酵可以不依赖活的酵母细胞。那么,引起发酵的物质到底是什么呢?有些科学家推测是蛋白质,并试图将酶从酵母提取液中分离出来,得到纯酶,但由于技术上的困难都未成功。
1917年美国科学家萨姆纳从资料中得知刀豆种子中脲酶含量很高,便决定从刀豆种子中提取纯酶,在经历了一次又一次的失败后,终于用丙酮作溶剂得到了脲酶结晶。这种结晶溶于水后能够催化尿素分解成氨和二氧化碳。然后,他又用多种方法证明脲酶是蛋白质。这以后,经过科学家们的努力,有数千种酶经研究证明是蛋白质。
酶是活细胞产生的具有催化作用的有机物,其中绝大多数是蛋白质。20世纪80年代初期,科学家发现RNA也具有生物催化作用,它们被称为“核酶”(ribozyme),这一发现改变了生物体内所有的酶都是蛋白质的传统观念,被认为是生化领域最令人鼓舞的发现之一。目前,核酶应用的理论基础是核酶的专一的“剪切活性”和“剪接活性”。“剪切活性”是指核酶专一破坏RNA尤其是mRNA的性质,利用这一特性设计核酶以阻断有害基因表达和病毒RNA的复制,达到治疗疾病的目的;另外,可利用核酶的“自我剪接活性”设计核酶,对突变的mRNA分子进行修复,使之转变为具有正常功能的mRNA。
虽然酶绝大多数是蛋白质,但是许多酶并非由纯蛋白构成,它们还需要非蛋白的辅助因子(cofactor)和辅酶(coenzyme)才能完成其催化作用。通常将无机金属离子称为辅助因子,将有机化合物称为辅酶。有些辅助因子或辅酶与酶蛋白的活性部位结合紧密,有的则结合疏松。大多数氧化还原酶类的辅酶是一些具有核苷酸结构的维生素。如烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD+,又称辅酶Ⅰ)、烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADP+,又称辅酶Ⅱ)、黄素腺嘌呤二核苷酸(flavin adenine dinucleotide,FAD)、黄素腺嘌呤单核苷酸(flavin adeninemononucleotide,FMN)等都是一些传递H+和电子的重要辅酶。现已知绝大多数维生素作为酶的辅酶在生物代谢中发挥重要作用。
二、酶的催化机理
酶是如何催化生物化学反应的呢?酶通过降低活化能的方式加速反应。在一个化学反应体系中,反应物分子所含的能量高低不同,每一瞬间并非全部反应物分子都能进行反应。只有那些具有较高能量的活化分子才能在分子碰撞中发生化学反应。反应物中活化分子越多,反应速率越快。活化分子比一般分子高出的能量称为活化能(activation energy)。活化能还可定义为,在一定温度下1摩尔反应物全部进入活化态所需要的自由能,单位为kJ/mol。反应所需的活化能愈高,相对活化分子数就愈少,反应速率就愈慢。而酶可以降低反应所需要的活化能,使更多的底物分子有足够的能量来形成产物,从而加快反应速率(图4-4)。
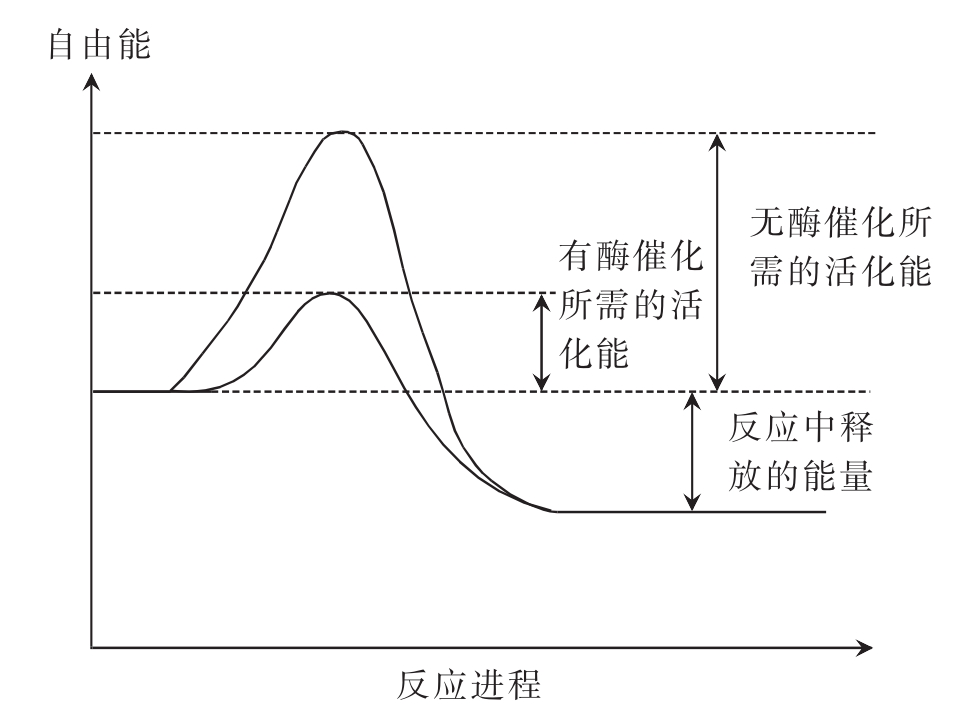
图4-4 酶降低反应活化能
在催化反应中,酶和底物结合成中间产物,经过中间反应所需要的活化能比底物直接生成产物所需的活化能小很多。
研究者在探索酶如何降低反应的活化能时付出了很大的努力,并对这个过程了解得越来越清楚。在催化反应中,酶(以E代表)首先和底物(以S代表)形成不稳定的中间产物S-E,这个中间产物进一步分解形成产物(以P代表)和酶本身:
S+ E→S-E→P+ E
经过中间反应所需要的活化能比底物直接生成产物所需的活化能小很多。这是因为当酶和底物结合时,底物分子能诱导酶分子的构象发生一定的变化,使活性中心部位与底物可以很好地结合。这种诱导契合关系使活性中心化学基团有最佳的定位,从而导致底物分子特定的化学键伸直或弯曲以及化学键的断开,启动了化学反应的发生,即打破了“能障”,降低了化学反应所需要的活化能(图4-5)。
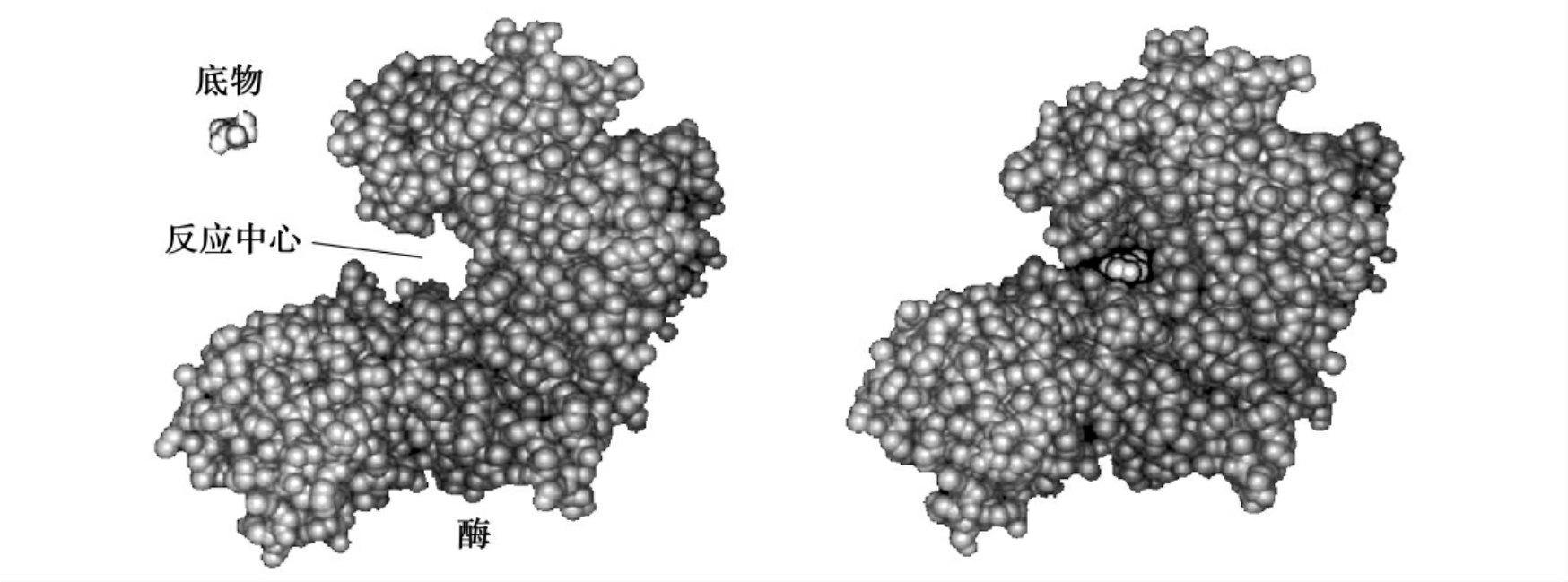
图4-5 酶与底物的诱导契合关系
当酶和底物结合时,底物分子能诱导酶分子的构象发生一定的变化,使活性中心部位与底物可以很好地结合。这种诱导契合关系使活性中心化学基团有最佳的定位,从而导致底物分子特定的化学键伸直或弯曲以及化学键的断开,启动了化学反应的发生,即打破了“能障”,降低了化学反应所需要的活化能。
酶与一般的无机催化剂相比具有催化效率高、专一性强、活性受到调节和控制等特点。酶的催化效率比无机催化剂高106~1013倍。据报道,如果在人的消化道中没有各种酶参与催化反应,那么在体温37℃的情况下,要消化一顿简单的午餐,大约需要50年,而动物吃下的肉食,在消化道内只要几小时就可完全消化分解。酶的专一性是指酶对催化的反应和反应物有严格的选择性,酶往往只能催化一种或一类反应,作用于一种或一类物质,如乳酸脱氢酶只能催化L-乳酸,不能催化D-乳酸的反应。酶活性受到调节和控制是酶区别于一般催化剂的重要特征,生命体通过激素、反馈抑制、酶的合成与降解来调节酶活性,从而维持生命活动的高度有序性。
酶大多数是蛋白质大分子,外界的许多物理和化学因素都会对蛋白质分子的结构和构象产生影响。影响酶活性的主要因素包括底物浓度、温度、pH值和抑制剂等(图4-6)。
1.底物浓度的影响
在底物浓度很低时,酶缓慢催化形成产物,因为它能接触到底物分子的机会很少。当底物浓度增加时,酶常常更多地结合底物,反应速率加快,即酶催化反应的速率随底物浓度增加而加快。然而,底物浓度的进一步增加并不会导致有更大的反应速率,因为有限的酶分子已结合底物,即酶被底物饱和。
2.温度的影响
一般来说,在一定范围内,酶的活性随温度升高而增加,超过一定的温度界限,活性即下降。酶活性随温度变化的曲线是钟形曲线,有一个最高点,即最适温度。适当地升温促进了分子的运动,提高了底物与酶活性位点的结合速度,但超过一定的温度界限,由于酶蛋白肽链间的氢键、离子键或其他更弱的作用力被破坏,导致蛋白质变性,使酶活性很快降低。温血动物的酶最适温度是35~40℃,植物的酶最适温度是40~50℃。一般情况下酶在60℃以上变性,少数酶可耐高温,如某些生长在温泉中的细菌所含的酶的最适温度高达70~80℃。
3.pH的影响
大部分酶的pH-酶活性曲线是钟形曲线。每一种酶只在一定的pH范围内具有较高的活性,而且有一个最适pH值。一般酶的最适pH在6~8,少数酶需偏酸或偏碱性条件,如胃蛋白酶最适pH为1.9,而胰蛋白酶的最适pH为8.1。pH对酶活性的影响主要是因为pH影响酶的构象,也会影响酶或底物的解离。
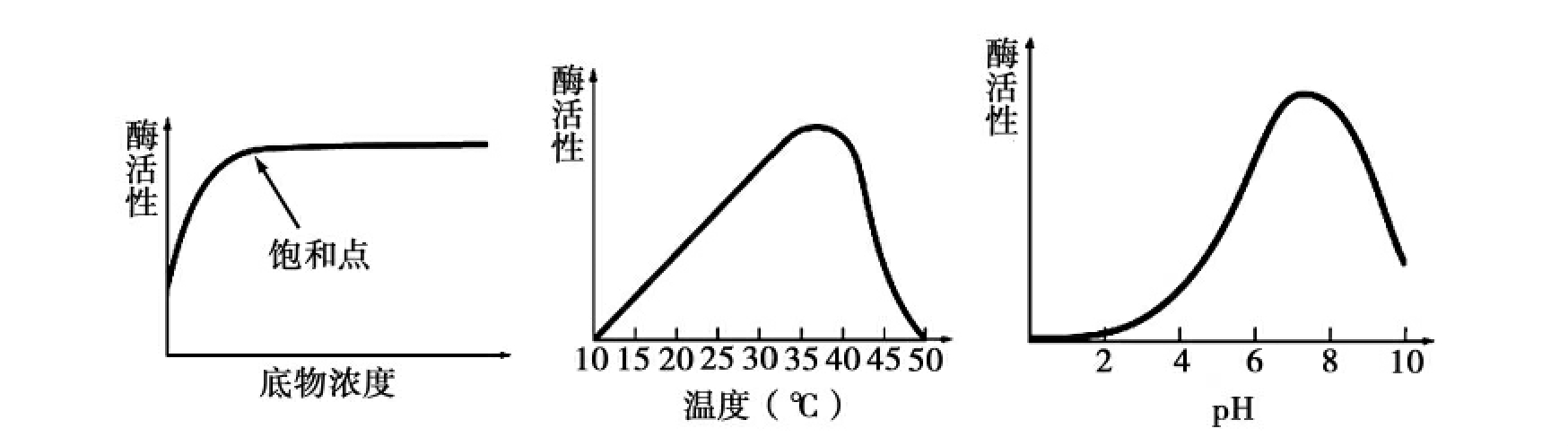
图4-6 底物浓度、温度和pH对酶活性的影响
酶催化反应的速率随底物浓度增加而加快,然而底物浓度增加到一定程度,反应速率不再增加,因为酶被底物饱和。在一定范围内,酶的活性随温度升高而增加,超过一定温度界限,活性即下降,即酶活性随温度变化的曲线是钟形曲线。酶活性和pH的关系也有一个相似的曲线,在pH太大或太低时,酶都会产生变性,只有在一个合适的pH值时,酶才能发挥其最大效力。
4.抑制剂的影响
使酶活力下降,但不引起酶蛋白变性的作用称为抑制作用。能引起抑制作用的物质叫做酶的抑制剂。根据抑制作用是否可逆分为两类:不可逆抑制(irreversible inhibition)与可逆抑制(reversible inhibition)。
(1)不可逆抑制
此类抑制剂通常以共价键与酶结合,不能用透析、超滤等物理方法除去。由于被抑制的酶分子受到不同程度的化学修饰,故不可逆抑制也就是酶的修饰抑制。常见的不可逆抑制剂有:有机磷化合物、有机砷和有机汞化合物、氰化物、重金属盐、烷化剂等。
(2)可逆抑制
此类抑制剂与酶的结合是可逆的,可用透析法除去,恢复酶活性。根据抑制剂与底物的关系,可逆抑制分为:竞争性抑制(competitive inhibition)和非竞争性抑制(noncompetitive inhibition)。
竞争性抑制:抑制剂结构与底物类似,与底物竞争酶的活性中心,从而阻止底物与酶的结合。最典型的例子是丙二酸对琥珀酸脱氢酶的抑制。琥珀酸脱氢酶在三羧酸循环中催化琥珀酸氧化生成延胡索酸,由于丙二酸非常类似于该酶正常底物琥珀酸,所以它是琥珀酸脱氢酶有效的竞争性抑制剂(图4-7)。竞争性抑制剂在医药领域占有重要地位。例如,磺胺类药物(如对氨基苯磺酰胺)就是竞争性抑制剂,它与对氨基苯甲酸的结构相似,可抑制细菌二氢叶酸合成酶,从而抑制细菌生长繁殖。人体可利用食物中的叶酸,而细菌不能利用外源的叶酸,所以细菌对此类药物敏感。
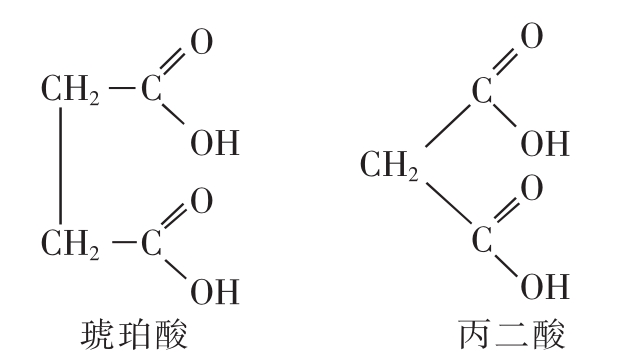
图4-7 琥珀酸和竞争性抑制剂丙二酸的比较
非竞争性抑制:非竞争性抑制剂结合在酶的非活性中心部位,导致酶分子形状发生改变,不能与底物分子相匹配和结合(图4-8)。
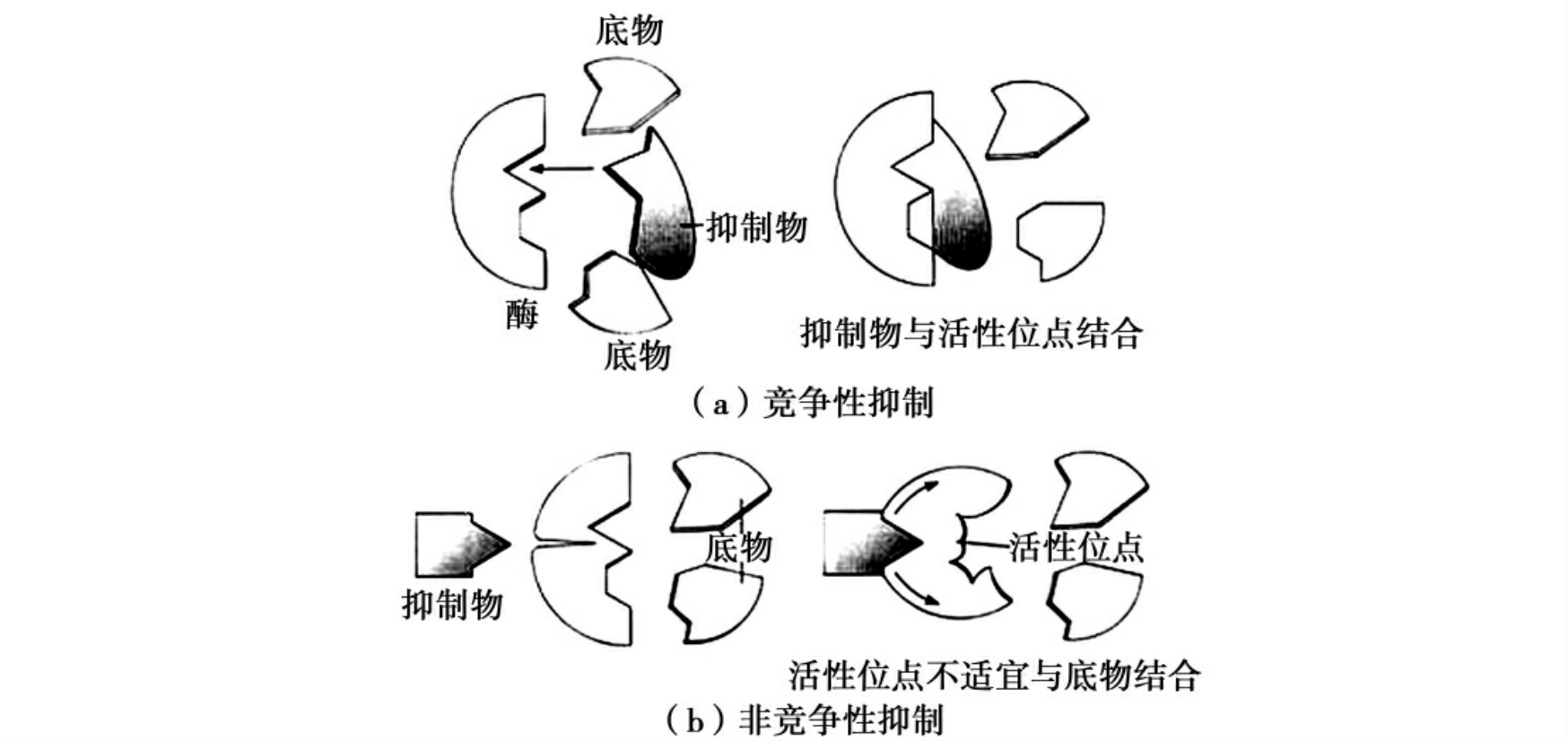
图4-8 酶的竞争性抑制和非竞争性抑制
抑制剂结构与底物类似,与底物竞争酶的活性中心,从而阻止底物与酶的结合。非竞争性抑制剂结合在酶的非活性中心部位,导致酶分子形状发生改变,不能与底物分子相匹配和结合。
前面已经提到代谢过程几乎都是酶促有机反应,而生物体的有机反应大体可归纳为以下几类:(1)氧化—还原反应;(2)基团转移反应;(3)碳—碳键的形成或断裂反应;(4)水解反应;(5)消除、异构化和重排反应。其中氧化—还原反应是细胞内最重要的一类生物代谢反应。
氧化—还原反应实质是电子的得失反应:失去电子的过程是氧化反应,获得电子的过程是还原反应,失去电子的物质是还原剂,得到电子的物质是氧化剂。细胞中氢及其电子从一个化合物向另一个化合物转移时发生氧化—还原反应,被转移的电子所携带的能量便储存在新的化学键中。例如:
NAD++H++2e→NADH
NADP++H++2e→NADPH
FAD+2H++2e→FADH2
其中NADH、NADPH、FADH2这些还原型产物还可将所接受的电子和氢传递给其他传递体。根据热力学原理,电子的传递必须由低氧化—还原电位(E0)物质向高氧化—还原物质传递。氧化—还原电位是以氢电极为标准并以氢原子氧化—还原体系的E0值为对照来反映还原剂失去电子能力大小的电位差值。E0值高的体系较E0值低的体系容易接受电子,其本身是较强的氧化剂;E0值低的体系较E0值高的体系容易失去电子,其本身是较强的还原剂。生物体内一些重要物质的氧化—还原电位已经测出,如表4-1所示。
表4-1 生物体部分重要氧化—还原体系的氧化—还原电位

氧化—还原体系对生物体之所以重要,不只是因为生物体内的许多重要反应属于氧化—还原反应,更重要的是因为生物体所需的能量来源于体内所进行的氧化—还原反应。
除了氧化—还原反应,常见的生物反应还有基团转移反应,碳—碳键的形成或断裂反应,水解反应,消除、异构化和重排反应。所谓基团转移反应是指某些功能基团从一个分子转移至另一分子,最常见的基团转移反应包括酰基转移反应、磷酰基转移反应和葡糖基转移反应。
分解代谢与合成代谢实质就是以碳—碳键的断裂或形成为基础的反应过程。葡萄糖经过5次断裂反应成为CO2,葡萄糖的生物合成则是碳—碳键的形成过程。此类反应可以举出3种类型:羟醛缩合反应、克莱森酯缩合反应和β—酮酸的氧化脱羧反应。
许多生物大分子分解成小分子单体都涉及水解反应,例如淀粉水解成葡萄糖、多肽水解成氨基酸、DNA或RNA分子水解成许多单核苷酸等等。ATP水解成ADP是另一类直接放出能量的水解反应。另外,苹果酸脱去一分子水生成延胡索酸的反应属于消除反应。生物化学中的异构化反应是指一个氢原子在分子内迁移,即质子从一个碳原子脱离,转移到另一个碳原子上,在代谢中,存在最多的异构化反应是醛糖—酮糖互变反应。重排是C—C键断裂,又重新形成的反应,葡萄糖-1-磷酸转变成葡萄糖-6-磷酸属于重排反应。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










