



第二章 如何采集菌类标本、鉴定需求及分类的重要性
王向华
引 言
本章主要介绍如何采集菌类子实体标本。与产生叶、花、果等复杂器官的植物相比,菌类子实体可供分类的外观特征较少。要达到准确的鉴定和深入的分类学研究,必须借助于对一些显微特征,如孢子(颜色、大小、形状、纹饰)、表皮(包被)、囊状体等的观察,而这些组织均取自干标本。因此,与植物标本相比,好的菌类标本对于菌类的分类鉴定更为关键。有很多文献描述了菌类标本的采集、制作和保存的详细过程,包括Baxter和van der Linde(1999)的手册。该手册可通过www.spc.int/pps/SAFRINET/fung-scr.pdf下载。
标本采集与野外记录
理想的菌类标本应符合以下标准:
·一份标本内包含从不成熟到完全成熟的不同生长阶段的多个个体,但应避免采集那些过熟的、近于腐烂的标本;
·烘烤彻底、得当,最大可能地保持子实体新鲜时的形状和颜色;
·标本各部位的组织在相应的试剂中能够得到理想的恢复;
·附有对产地、生境、新鲜特征的详细记录;
·照片和孢子印也可附在标本内作为补充标本信息;
·能够在长时间保存后不生虫、不长霉。
采 集
从自然生境中获取标本
采集生于地表的子实体时,应避免用手直接将其拔出。小心地用小刀剖开子实体与基质(土壤、枯枝落叶等)接触的部位后,注意观察是否有部分子实体埋于地下或子实体与地下的某些东西相连。一些菌类如蚁巢伞属(Termitomyces)、暗金钱菌属(Phaeocollybia)、小奥德蘑属(Oudemansiella)的子实体常具假根。采集这些标本时,若不注意会将全部或部分假根遗留在土中。虫草属(Cordyceps)的子座生于死的昆虫体内或地下真菌大团囊菌(Elaphomyces spp.)上,若不小心可能会将子座与地下部分分离。采集有菌核的种类如猪苓(Polyporus umbellatus)、黑柄炭角菌(Xylaria nigripes)时,需要注意埋于地下的菌核。采集那些长在腐木或枯枝落叶层上的子实体时,最好连同子实体着生的基质一起采集,这样可以较好地保持子实体的形态,并为鉴定提供生长习性方面的参考。采集菌根菌的子实体时,一并采集共生树种(或伴生树种)的枝、叶、花、果标本,有助于在鉴定真菌标本时提供辅助信息。
在采集外生菌根菌时,事先了解共生树种对于找到真菌很有帮助。如在本书第七章提到的,在北温带,外生菌根菌多生于松科、壳斗科、桦木科、杨柳科树下;在热带或南温带,则多生于龙脑香科、金缕梅科树下;在非洲热带地区,豆科树下(如miombo woodland)也可以找到外生菌根菌。
地下真菌(hypogeous fungi)的子实体全部或部分埋生于地下,标本比较难采集。请当地熟悉这些菌类的采集者做向导会大大提高采集的效率。块菌属(Tuber)的一些种,如黑孢块菌(T.melanosporum)和印度块菌(T.indicum),在产子实体的地表,往往出现大片枯草,如火烧过一般。这一被称为菌塘(br0lé)的区域可以用来判断块菌的发生场所。地下真菌通常是一些噬齿类动物的食物,因此噬齿类动物的洞穴、掘食的痕迹以及留下的粪便,均可以作为地下菌分布的指示信号。事先了解该类菌类的共生树种也对采集很有帮助。
采集标本时,应携带不易变形、具有足够空间的容器,如竹篮、塑料篮等。较大或较硬的子实体可直接放入篮内,也可先将其放入纸袋内再放入篮内,这有助于避免挤压或不同标本间的污染。较小(菌盖<3厘米)或软而不耐挤压的子实体,可放入具分隔的塑料容器内(彩图2.1A)或独立容器内(彩图2.1B)。具黏液或易失水的子实体,可装入干净的小塑料袋,并置于不易被挤压的地方。在同一次采集中,如果采集到的菌类仅与某些树种共生,可将共生树种的标本一并采集,并与菌类标本放入同一个袋内,以免混淆。
在采集菌类的野外工作中,必须留有一定的时间用来记录标本。根据经验,如果要对标本进行详细记录,一般1小时内记录的标本不会超过4份。可据此推算需要为标本记录预留多少时间。一般的经验是,在标本较多的情况下,下午2点前结束采集并回到驻地进行记录较为合适。如傍晚时分才结束采集,晚上进行标本描述和登记可能会降低记录的质量。
从本地市场获取标本
在全世界的很多地区,特别是多民族聚居的地区,如中国、墨西哥、泰国等地,在蘑菇生长季节,有很多大大小小的的野生菌市场(Jones et al.,1994;王向华、刘培贵,2002; Pérez-Moreno et al.,2008)。从市场上不仅可以获得标本,还可以获取民间采集和利用菌类的相关知识(彩图2.2)。市场上出售的菌类繁多,来源广泛,有机会遇到稀有或珍贵的标本。故在一些分类学文献中常常发现某些菌类的模式标本(type specimen)采自市场,典型的如丛枝瑚菌(Ramaria ssp.)和牛肝菌类(boleti)(Chiu,1948; Petersen&Zang,1989)。
在市场上收集标本时,需要对出售者、采集者、收购者进行访谈,以获取关于标本的产地和生境等信息。在市场上出售的,既有菌类的新鲜材料也有干材料。由于子实体在干燥处理后很难保持原貌,因此在从市场上收集干的菌类标本时,要给予特别的注意。比如,那些看来一样的标本可能会包含多个种的子实体。采集时尽可能了解标本的产地、用途和地方名。这些信息对后期鉴定很有帮助。
市场上出售的子实体往往残缺不全或形态发生了变化。例如,鹅膏属(Amanita)子实体基部的菌托常被人为去除,其菌幕(volval remnants)也很容易因人为触摸而脱落;牛肝菌类基部有时会被削去,从而无法观察到其基部菌丝的形态。小奥德蘑属(Oudemansiella),如鳞柄小奥德蘑(O.furfuracea),新鲜时其菌盖表面黏滑,但由于采集后放置时间较长,黏液常会变干;红菇属致密组(Russula sect.Compactae)和乌芝属(Amauroderma)一些种的子实体常因摩擦发生色变。以上这些情形均会给鉴定带来一定困难。所以,在市场上,采集完整的、状态较好的标本很重要。如果有可能,应采集多个成熟度不一致的个体作为一份标本。
在市场上做菌类资源调查时,在现场不一定能识别所有的种。对于初学者而言,对于市场上出现的分类群往往缺乏足够了解。在这种情况下,如果要准确了解市场上每种菌类出现的频度、贸易量、地方名、应用价值等,可采纳如下的程序:在市场上先对每一种菌类进行编号,然后记录每个编号所对应的菌类的详细信息,最后再将后期标本鉴定后得到的名称替换编号。在市场上进行拍照,记录照片号码样式与标本编号一一对应,这对鉴定和后期分析非常有用。由于初学者对标本特征难以做出正确的判断,如果发现某些标本间存在差别,却又无法判断该差别是否为种间差别时,应将标本分别编号。避免一开始就按照地方名的方式采集标本,这样做极有可能将属于不同种的标本混为同种,从而得出错误的结算。
记 录
与菌类标本有关的记录包括产地、生境和习性,以及标本的新鲜状态特征。其中的生境信息,如腐木的种类、共生树种的种类、海拔高度等对于分类鉴定具有重要参考价值,故在野外采集时就应记录这些信息。有条件时,可用GPS仪记录采集点的坐标信息。菌类分类在很大程度上依据的是子实体新鲜状态的特征,而这些特征在干燥后常会发生改变,因此及时记录这些特征对于准确的分类鉴定至关重要。
记录子实体的新鲜特征应在采集当天和在自然光下进行。如果在晚上记录,应使用光强度足够的白色日光灯。如果盖表质地特殊,如胶粘、水浸状等,在采集时就应记录下来。记录前,先将用铅笔写好采集号的标签系于成熟个体上。对子实体各部位尺寸进行测量时应基于成熟的个体。记录颜色可参考两本色谱: Kornerup-Wanscher色谱和Ridgway色谱。如在色谱上无法找到匹配的颜色,可以用两个色码标定的范围来表示。如果发现子实体(或其乳汁)在受损或放置后有变色的情况,应在半小时内注意变色是否会进一步发展。记录时可以尝取子实体组织的味道,但品尝后应将组织吐出并漱口(注意:初学者难以预先知道哪些是有毒的种类,因此牢记这一点非常重要)。孢子印应该在采集当天搜集。锡箔纸是搜集孢子印较好的材料。这种材料不会吸水、易于分辨颜色且容易用铅笔编号。采集的方法:取部分或全部菌盖(或其他着生产孢组织的部位,如子囊盘),将有产孢组织的一面倒扣向锡箔纸,静置12~24小时后将组织移开,观察并记录孢子印的颜色后,将锡箔纸对折装入有一定硬度的纸袋内。对于那些较薄的孢子印,可以用铅笔将其范围划定,以便以后查验(布里德森与福门,1998)。
标本的记录纸一式两份,一份由采集人保留,一份随同标本放置。在野外记录时可以用复写纸直接复印采集记录,也可回到实验室后复印或抄写。记录最好用铅笔(2B),其字迹不会因水浸或长时间保存而消退。彩图2.1C为一份乳菇属标本野外记录表。
干 燥
标本干燥应在记录完成后进行,最好是记录完毕后立即烘烤。小的或将来要提取DNA的标本最好用硅胶干燥。硅胶干燥时,将子实体用吸水纸包裹后放入装有硅胶的自封袋内。虽然阳光暴晒干燥标本的做法有时在外观上也可以达到要求,但由于温度不够,无法杀死子实体内的虫卵,保存过程中可能发生虫蛀情况,不建议使用。
与植物标本放入标本夹再烘烤不同,菌类标本需直接放在烤架上。硬而韧的标本可以整体烘烤。较大而肉质的标本,如牛肝菌类的标本,可将子实体纵剖为两半或更多瓣后再烘烤。烘烤较小标本时,需将标本放在绵纸上,或用绵纸将标本包裹起来,再放在烤架上,否则这些标本有可能从烤架的网眼中漏掉。已散放孢子的腹菌类标本应用透气的纸袋装好后再放在烤架上,以防止其孢子污染其他标本。在烤架上摆放标本时,将特征明显相异的标本摆在一起,而将相似的种类分开摆放,以免干燥后难以辨别。
干燥标本可采用烘烤水果蔬菜的带有鼓风设备的电烘箱(彩图2.1D),也可自制简易烘箱(彩图2.1E)。烘干温度50~60℃较合适,该温度不仅能最大程序的保持标本原始状态,还能杀死虫卵。自制烘箱应设计为可折叠的,以便携带。烘箱制作可参考以下尺寸及要求:高60~80厘米,横截面正方形,边长30~40厘米,内置4~5层金属筛网,筛网间距10~12厘米,标本距加热源40~80厘米,加热源为300W的电炉或煤油炉。使用时置于平整地面上,地面摆放加热源,烘箱上口覆盖布块,通过敞口的大小调节温度高低。烘烤完毕的标本迅速装袋,并置于防压、防碎的容器内。
保 存
标本在保存之前应最大限度地干燥。野外烘干过的标本,特别是那些大而肉质的标本,带回实验室后常发生返潮。所以,在入馆前最好再次检查标本的干燥程度。有的标本馆要求在入馆前将标本进行冷冻,但切记在冷冻时将标本使用密封性强的塑料袋包装。标本从冰箱内取出后,回到常温前,不能从袋内取出,否则易吸水受潮。放置菌类标本理想的容器是纸盒,可以很好地保持子实体的形状。也有一些标本馆使用纸袋或塑料袋,并将纸袋装于台纸上。先将标本放入纸袋内,再将纸袋放入塑料袋内是最好的保存方法。这种方法不仅可有效防潮,还可保证标本的组织在观察时得到较好的恢复,同时还可防止袋子与标本间摩擦产生的静电吸附标本碎片(布里德森和福门,1998)。特别小的子实体在装入标本馆规定的袋内之前,最好先装在尺寸合适的小袋内,以免丢失。
分 类
鉴 定
鉴定的目的是为标本找到正确的科学名称。然而,在真菌的鉴定中,由于研究的基础相对较弱,常常难以找到正确的名称,或者根本就不存在正确的名称(比如鉴定的对象是个尚未被描述的新种)。在这种情况下,根据现有特征和文献找到相接近的名称也可视为理想的鉴定结果。
做出准确的鉴定应具备以下条件:广博的分类学知识、标本新鲜特征的详细记录、显微特征的细致观察、大量的文献查阅。前文已对如何记录标本进行了介绍,下面将对显微观察和相关文献进行介绍。
细致的显微观察和研究
显微观察和记录对于鉴定菌类标本至关重要。现代真菌分类学研究对显微特征越来越重视(Singer,1986),对各个显微成分的测量也越来越精准。显微特征观察得越详尽、细致,参考价值越大。显微结构研究包括3个步骤:对拟观察部位的正确切片、详尽的显微观察和记录以及必要的显微绘图。图2.1是一份对乳菇属标本进行显微观察时所做的记录。
菌类标本鉴定中最常观察的对象和最有用的特征是孢子。孢子被认为是菌类的“身份证”。与孢子有关的性状包括:颜色、大小、形状、表面纹饰。在蒸馏水或KOH中,有色孢子不需染色便可直接观察,无色孢子需要用染色剂染色(如刚果红或梅试剂)。观察和测量孢子时,一般从侧面观进行。初学者对于孢子的判断常比实际的宽且圆,这是由于方位选择不当造成的。由于表皮(包被)和子实层组织中包含的信息在分类鉴定中越来越重要,显微观察中应给予足够重视。在担子菌中,子实体组织中锁状联合(clamp connections)的有无与多寡也常作为分类的指标,如在对Amanita、Ramaria、Tricholoma、Cantharellus/Craterellus、Sarcodon的标本的鉴定中(Corner,1967; Singer,1986; Petersen&Zang,1989; Stalpers,1993;杨祝良,2005)。判断有无锁状联合一般可先选取子实层进行观察,因为这一部位往往是锁状联合最易产生的地方。
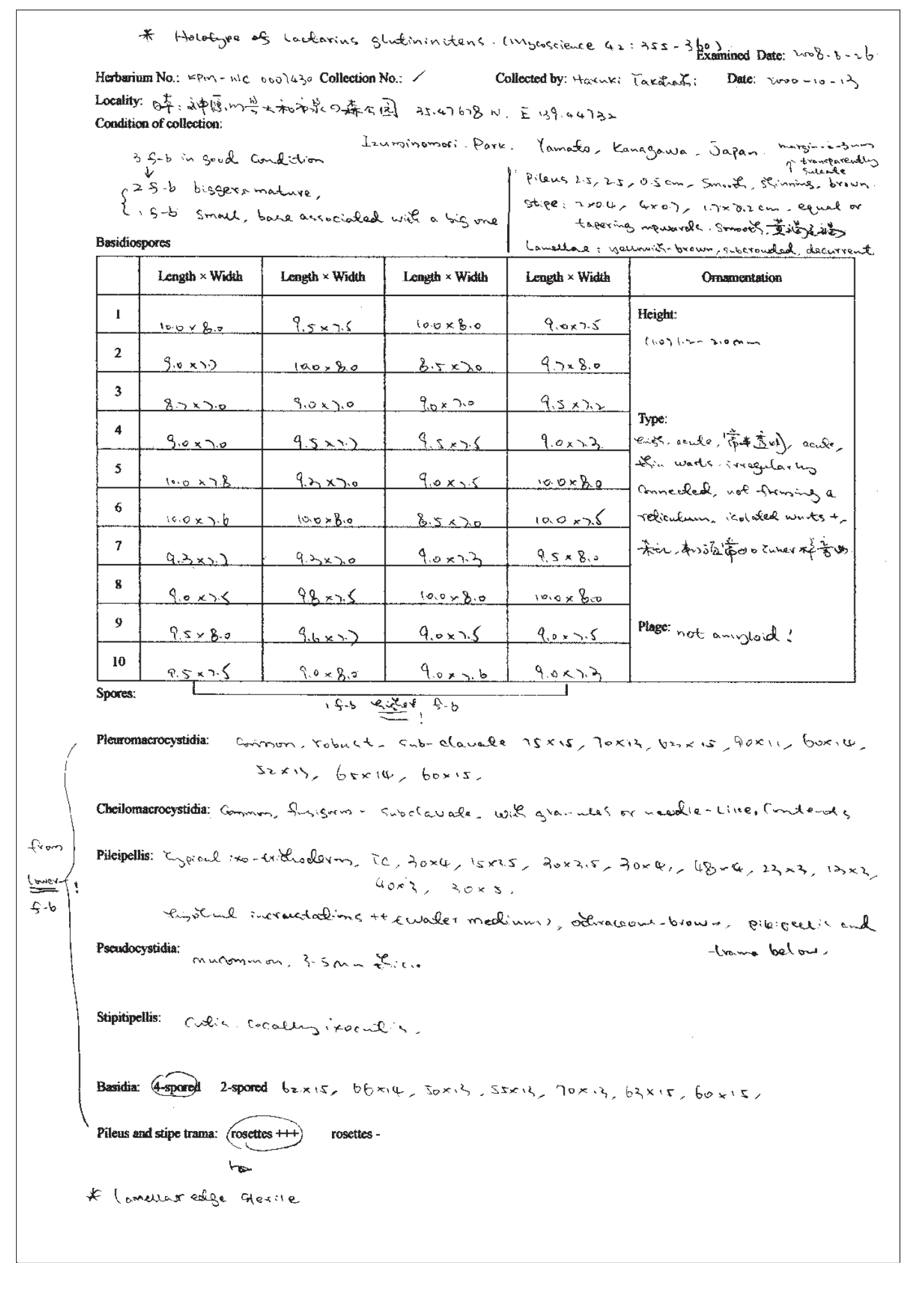
图2.1 一份乳菇属标本的部分显微观察记录
菌类的显微观察不需要复杂的切片技术,徒手切片即可满足大多数观察需求。用镊子夹取小片菌褶(或产孢组织)放在特定的试剂中即可观察孢子。对子实体表皮(如盖表皮、柄表皮或包被)进行切片时,应注意将刀片与表面保持垂直后沿径向或纵向切片。当菌丝不能较好地分开时,用镊子一头轻敲玻片有助于恢复菌丝,特别是那些皱缩膨大的菌丝(杨祝良,2005)。当遇到难以恢复的皱缩菌丝时,可将切下的子实体组织浸泡在KOH或蒸馏水中,过夜后再染色观察。无论是无色还是有色菌丝,使用KOH浸润后再加入少量的刚果红进行染色,都可有效地提高菌丝的恢复效果。
真菌菌丝细而壁薄,孢子较小[2~15(30)μm],观察时应使用100倍浸油(immersion oil)物镜。使用40倍物镜往往无法看到细节,对各个部位的测量也不准确。使用显微镜观察菌类组织时,要时刻注意显微镜光圈的大小。较大的光圈对观察表面有纹饰的孢子很有用;而对数层菌丝进行观察时,需将光圈调小。使用具相差(phase contrast)功能的显微镜对观察和寻找锁状联合较为容易。这样的显微镜容易突出反映菌丝壁。相差显微镜还适合观察透明的未经染色的孢子和胶质化菌丝。显微镜观察要求快速。长时间处于介质中的组织其菌丝轮廓会逐渐变得不清晰,最终失去观察的价值。
当一些显微特征,如孢子形状和表面纹饰、盖表皮式样和菌丝构成等,难以用语言进行准确描述时,可采用绘图来表示。绘图可通过安装于显微镜上的绘图管进行。在没有绘图管时,需要不时地通过目镜上的测微尺对画在纸上的图进行校正和核对,以免表达的图像与真实的图像间产生偏差。在绘制孢子时,画在纸上的图与原物的比例保持在3000~5000倍,即纸上的1厘米相当于实物的2~3μm较合适;对菌丝进行绘图时,比例控制在1000~2000倍较合适。
进行菌类显微观察和研究必备的仪器和材料如下:带有目镜测微尺的生物显微镜、用于切片的双面剃须刀片、一头为半圆形的尖头不锈钢镊子、用于恢复浸润干标本组织的5%~10% KOH溶液、用于判断孢子表面是否为淀粉质的梅试剂(Melzer's Reagent)[配方参考Kirk等(2001)]和用于对无色组织进行染色的刚果红(Congo Red)(水溶液,浓度以适宜观察为佳)。
鉴定参考文献
可供鉴定的参考文献很多,难以一一列举。这里仅罗列用于一般快速鉴定的常用文献。带有彩色图片的蘑菇图鉴:
世界菌类: Laessoe,2002; Hall et al.,2004。
日本菌类:今关六也和本乡次雄,1957,1965; Imazekiet al.,1988; Hongo&Izawa,1994。
中国菌类:中国科学院微生物研究所真菌组,1979(毒蘑菇);应建浙等,1982(食用蘑菇);应建浙等,1987(药用菌)。
美洲菌类: Arora,1986; Miller&Miller,2006。
欧洲菌类: Courtecuesse&Duhem,1995; Jordan&Wheeler,2005; Phillips,2006。
利用检索表对伞菌标本进行大致的属一级及种一级鉴定可参考Moser(1983)。
鉴定程序
鉴定标本是一个需要多次反复进行的过程。初学者可参考子实体的外观或必要的显微特征对标本进行大致归类,如归为子囊菌、担子菌、伞菌、多孔菌等。在对标本,特别是伞菌类标本进行属一级的鉴定时,孢子印颜色(或显微镜下的孢子颜色)和孢子形态是十分有用的特征。在进行属下鉴定工作时,不同的属侧重的特征不同,如红菇科(Russulaceae)的红菇属(Russula)和乳菇属(Lactarius)侧重的是子实体大小、颜色和味道、孢子印颜色、孢子纹饰、囊状体多寡和有无盖表皮及其式样;丛枝瑚属(Ramaria)侧重的是子实体颜色、分枝式样、孢子大小及表面纹饰和锁状联合的有无;丝盖伞属(Inocybe)侧重的是盖表鳞片(毛)、孢子和囊状体的形态;牛肝菌类的一些属,如牛肝菌属(Boletus)、粉孢牛肝菌属(Tylopilus)常常侧重于盖表、柄表及菌孔颜色和变色、柄表网纹及盖表式样。初学者在鉴定标本时常会发现,一些有鉴定价值的信息在前期野外记录或显微观察中未被充分注意到。平时有意地对这些疏漏进行总结和改进,将有助于快速提高对各个类群的理解。鉴定中,对于文献中使用的但前期又未观察到的显微特征,一定要进行再观察并作出判断。想当然地对这些特征进行推测常会导致错误的鉴定。
除一些常见种外,为一份标本找到合适的种名是一个较为耗时的过程。由于普遍存在种内个体间变异,在与文献中的相关描述进行对比时,常会发现或多或少的差异。为了避免对这些差异产生不合适的判断,应尽可能多地观察同一份标本内的不同个体和采自不同地点的不同标本。只有当一些差异表现稳定而连续时,才可以将这些差异作为种间差异看待。当不能找到完全合适的名称时,可暂时采用那些特征最为相近的种的名字。应尽可能地避免在不熟悉的类群内发表新种。
民间采集利用的菌类多为有一定知名度的种,先参阅一些带有彩图和简单描述的图册,对进行初步标本鉴定是一条快捷途径。在这些图册上找到相近的名称后,再参考其他带有检索表的文献做进一步核对。必要时,参阅专著类文献对标本进行更为准确的鉴定。
命 名
命名是分类学的一个重要内容。它涉及名称的使用问题,即根据现有国际植物命名法规(International Code of Botany,ICBN),决定哪一名称才是应该被采用的名称。命名的目的是给予一个分类群唯一的、稳定的名称。了解与命名相关的知识,对于理解系统分类学研究结果,比较分析不同文献对不同名称菌类的报道,避免使用过时的名称都具有实际的意义。与野生菌有关的命名学问题非常普遍。在很多时候,如Index Fungorum所展示的那样,异名(synonym)现象非常普遍。案例2.1列出了一些与真菌有关的命名学问题。
分子生物学在菌类分类学中的应用改变了一些长期使用的、传统分类群的概念和范围。在一些属、科、目级别的分类单元上发生了拆分与归并。Hawksworth(2005)认为,与这些变化密切相关的一个问题就是与菌类有关的命名问题。及时根据系统分类学的研究进展对相关的命名学知识进行更新,有助于更有效地利用菌类。
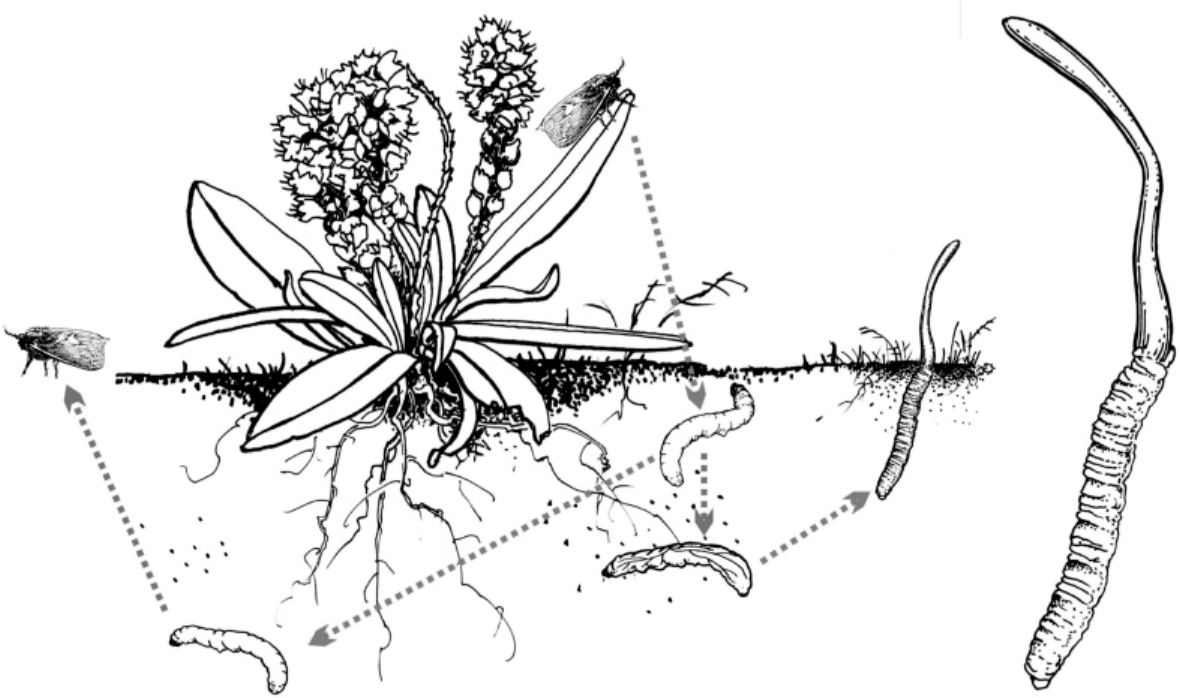
图2.2 药用菌冬虫夏草(Cordyceps sinensis)的复杂生活史
案例2.1 一些与食药用菌有关的命名学问题
冬虫夏草(Cordyceps sinensis):该种描述于19世纪,命名基于从中国的一个药材市场购得的模式标本。其模式标本子座不成熟,无法看到孢子(刘作易等,2003)。由于该种的概念不甚清晰,一些与本种相近的标本近年来被描述成了新种,如甘肃虫草(C.gansuensis)、尼泊尔虫草(C.nepalensis)、阔孢虫草(C.crassispora)等。刘作易等(2003)将多份采自青藏高原的冬虫夏草(C.sinensis)标本与这些种的模式标本进行对比后认为,这些种的特征与冬虫夏草的特征没有本质区别,都与C.sinensis为同种。因此C.gansuensis、C.nepalensis、C.crassispora这些名称均被作为C.sinensis的分类学异名。与冬虫夏草相关的另一个命名学问题:由于其生活史中除产生有性孢子——子囊孢子的有性型(telemorph)外,还具有产生无性孢子(分生孢子)的无性型(anamorph,多表现为霉菌的形式)。根据国际植物命名法规,这类真菌可以拥有两个名称,分别用于代表其有性型和无性型。如Cordyceps sinensis为冬虫夏草的有性型的名称,而Hirsutella sinensis为其无性型的名称(刘锡进等,1989; Jiang&Yao,2002)。最近的研究还产生了冬虫夏草的另一个命名法异名: Ophiocordyceps sinensis,同样用来指该种(Sung et al.2007)。
樟芝(Taiwanofungus camphoratus):名贵药用菌,特产于中国台湾。臧穆、苏庆华(1990)将采自台湾的樟芝标本作为模式发表新种Ganoderma camphoratum。Chang&Chou(1995)发表了新种Antrodia cinnamomea T.T.Chang&W.N.Chou,同指樟芝。Wu et al.(1997)研究了G.camphoratum的模式标本,发现该模式标本事实上是受到了Ganoderma sp.的孢子的污染,其分类学位置应在Antrodia内,又成立了新组合Antrodia camphorata,且将Antrodia cinnamomea作为A.camphorata的分类学异名。后来Wu et al.(2004)又根据基于DNA序列的系统学研究结果,以G.camphoratum为基名将樟芝归入一个新属Taiwanofungus。这样,T.camphoratus成了樟芝的最新名称。在以上这些名称中,Ganoderma camphoratum为A.camphorata的命名法异名,因为它们有着共同的模式。尽管A.camphorata的基名G.camphoratum早于A.cinnamomea,但当属名为Antrodia时,A.cinnamomea为樟芝的正确名称,因为该名称发表的日期(1995年)早于A.camphorata被合格发表的日期(1997年)。但若采用最新的系统学处理,则T.camphoratus为唯一的正确的名字,G.camphorata和A.camphorata均为其命名法异名,而A.cinnamomea则成为其分类学异名。虽然樟芝生于牛樟树Cinnamomea kanehirai而不是香樟树C.camphora上,T.camphoratus并不因其种加词“camphoratus”不合适而不能被采用。
松茸(Tricholoma matsutake):该种为一类重要的商品真菌,T.matsutake这一名称在全球,尤其在东亚被广泛使用。研究发现,该种与T.nauseosum(A.Blytt) Kyt9v.为同种(即为后者的分类学异名),前者的基原异名Armillaria matsutake S.Ito&S.Imai(1925)较后者的基原异名A.nauseosum A.Blytt(1909)晚。A.nauseosum这一名称与前者相比,远不如前者使用广泛。为了名称的稳定,有必要对后来的名称进行保留。Ryman et al.(2000)基于此提出了保留T.matsutake这一名称的建议,该建议被最新版维也纳法规采纳(Appendix IV)。
分类学的应用
按照形态特征以及彼此之间可能的亲缘关系对菌类进行归类的工作称为科学分类。科学分类采用双名法的拉丁学名,分类学本身是科学研究的重要内容。同时,分类学知识在指引人们对菌类的利用中也发挥着重要的作用。最典型的就是帮助区分有毒与可食的菌类。在食用了菌类而致死的案例中,绝大多数是因食用了剧毒鹅膏种类引起的(杨祝良,2005)。如果能正确识别鹅膏属(Amanita),就可以在很大程度上避免死亡事件的发生,尽管由此也会错过该属中一些可食的种,如恺撒鹅膏(A.caesaria)、隐花青鹅膏(A.manginiana)。再如裸盖菇属(Psilocybe)的250个种中具有致幻作用的就多达150个(Guzmán,2008)。具备了识别Psilocybe的知识就可以在很大程度上避免采食这类致幻蘑菇(hallucinogenic mushrooms)。同时,相近类群的真菌常具有相似的化学成分(刘吉开,2004),如在Psilocybe中,含有裸盖菇素Psilocybin具致幻作用的种类与不具裸盖菇素的种类可以在系统树上区分开来(Moncalvo et al.,2002; Maruyama et al.,2006)。故菌类之间的亲缘关系可以指导我们对菌类的化学成分进行研究。
分类学知识有助于我们了解真菌的生态习性。菌类中有很多科属包含的种类具有共同的生态习性,如蚁巢伞属(Termitomyces)的种类均与白蚁共生,虫草属(Cordyceps)的绝大多数种类生于昆虫体上(图2.2),红菇科(Russulaceae)、牛肝菌科(Boletaceae)、鸡油菌科(Cantharellaceae)、块菌科(Tuberaceae)的种类均为外生菌根菌,香菇科(Lentinaceae)、多孔菌科(Polyporaceae)(狭义)的种类均为腐生菌。如果具备了足够的分类学知识,就意味着同时具备了这些菌类的生态学知识。这些知识可以帮助快速地判断一些菌类人工驯化的难易程度及可能的驯化方式,同时也可以帮助理解这些菌类在各种陆地生态系统中的功能。
建立在准确鉴定基础上的分类学研究结果,在评估、比较不同社区的居民对菌类的利用状况时也发挥重要作用。分类学研究将世界各地的居民对菌类的应用联系为一个整体。例如,比较中国云南和墨西哥两地市场上出售的菌类可以发现,两地市场共有的菌类有23个属,分别占中国云南、墨西哥两地市场已知菌类属数的33.3%和62.2%,反映了两地在应用野生菌方面的共同知识。
人为分类和命名
世界各民族在利用野生真菌的过程中,为方便识别与交流,产生了民间的分类和命名,即地方名或俗名。民间对菌类的分类命名大致可分为以下几类:
(1)以子实体形态进行的分类和命名:为民间最常用的分类命名方法。根据形状,如在中国和墨西哥,Ramaria的种类均被称作“扫帚(把)菌”(王向华等,2004; Pérez-Moreno et al.,2008);根据颜色,如Cantharellus被称为“鸡油菌”,因为这些菌类的子实体的颜色如鸡油般的黄色; Boletus的种类在墨西哥被称为“鹿肝菌”或“肝菌”(deer liver,liver),在中国被称为“牛肝菌”,指其子实体的质地。
(2)以生态因子进行的分类和命名:红菇属的一些种类在中国云南被称为“栎树青”,表明它们生于壳斗科植物下;名贵药用菌Taiwanofungus camphoratus被称为“牛樟芝”,指其生于牛樟树上(Cinnamomum kanehirai)(Wu等,1997);口蘑属(Tricholoma)的一些种在中国云南被称为“松毛菌”,指它们生于松林下,同样如墨西哥的Hebeloma spp.,也称为松针菌(pine needle fungus); Cordyceps的种类被称为“虫草”,指该类真菌生于虫体上。
(3)以发现方式进行的分类和命名:在四川的一些地区,块菌属(Tuber)的一些种类被称为“猪拱菌”,这是因为该类真菌为地下真菌(hypogeous fungi),子囊果埋生于地下,被猪拱后可被发现。
(4)以出菇时间进行的分类和命名:乳菇属色奶乳菇组(Lactarius sect.Deliciosi)的种类由于在稻谷成熟的时候出菇,在云南被称为“谷熟菌”。玉蕈离褶伞(Lyophyllum shimeji)在云南景东被称为“九月菇”,指该菌于9月出菇(刘培贵,1993)。
科学分类命名与民间分类命名的关系
民间分类与科学分类间存在一定的联系和对应。这是因为民间分类的基础与科学分类的基础有相通之处,即均根据形态上的相似性。对民间分类和科学分类之间的关系进行总结,可以帮助理解民间在利用真菌资源方面的宝贵知识,有助于更好地识别、利用这些菌类。
民间分类命名中尽管存在很多同名异物和同物异名(案例2.2),但在一些基于地方名的分类中也可以找到与科学分类间的巧合。一个典型例子来自于乳菇属。该属的菌类在民间常被称为“奶浆菌”,意即子实体内含有“奶浆”(乳汁)。由于乳菇属是伞菌中为数不多的具有乳汁的类群,这一民间分类事实上也反映了这些菌类的亲缘关系。有意思的是,同样是乳菇属的种类,有一些具有稀少乳汁的种,如松乳菇(L.deliciosus)和红汁乳菇(L.hatsudake),在中国云南被称为“谷熟菌”,即谷熟时出菇的菌类。在科学分类中,“谷熟菌”刚好与松乳菇组(L.sect.Deliciosi)的范围相吻合。还有一个例子是牛肝菌类(boleti)的分类命名。在中国云南,牛肝菌属(Boletus)的一些种被称为“见手青”。这些种几乎都为B.subsect.Calopodes和B.subsect.Luridi的种类,对于那些子实体破损后不变色的Boletus sect.Boletus的种,均不被称为“见手青”。
民间命名分类除可提供有价值的系统分类信息外,还可提供其他如生境方面的信息,如斗菇属(Lentinus)的一个种L.strigellus在中国云南被称为“竹根菌”,即生于竹子根部;又如细柄丝膜菌(Cortinarius tenuipes)在中国云南南部被称为“黄栎窝”,表明这种菌丛生于壳斗科树下。掌握不同民族的菌类分类命名知识,还可以在标本缺乏的情况下帮助判断一些种类。
案例2.2 食用菌地方名中的同物异名与同名异物
相对于科学名称,民间的地方名称明显较少,这使得一些地方名包含数个类群。同物异名与同名异物现象在民间非常普遍。
同名异物的典型例子如鸡 。这是一类中国西南常用的名称,与此名称有关的至少有5个属的真菌:①鸡
。这是一类中国西南常用的名称,与此名称有关的至少有5个属的真菌:①鸡 :包括蚁巢菌属(Termitomyces)的所有种。使用时常在这一名称前加上修饰词,如黄皮鸡
:包括蚁巢菌属(Termitomyces)的所有种。使用时常在这一名称前加上修饰词,如黄皮鸡 、大果鸡
、大果鸡 等。②(露)水鸡
等。②(露)水鸡 :指小奥德蘑属(Oudemansiella)的鳞柄小奥德蘑O.furfuracea。之所以被赋予该俗名,可能是因为该种子实体基部具假根,与鸡
:指小奥德蘑属(Oudemansiella)的鳞柄小奥德蘑O.furfuracea。之所以被赋予该俗名,可能是因为该种子实体基部具假根,与鸡 菌属种类相似,但又因其盖表多具黏液,故被称为水鸡
菌属种类相似,但又因其盖表多具黏液,故被称为水鸡 。③油鸡
。③油鸡 :同指小奥德蘑属(Oudemansiella)的长根小奥德蘑鳞柄变种(O.radicata var.furfuracea),由于盖表具黏液,看上去有油脂感而得此俗名。④草鸡
:同指小奥德蘑属(Oudemansiella)的长根小奥德蘑鳞柄变种(O.radicata var.furfuracea),由于盖表具黏液,看上去有油脂感而得此俗名。④草鸡 :冠以此俗名的有两类:一类是鹅膏菌属(Amanita)的隐花青鹅膏(A.manginiana)和假褐云斑鹅膏(A.pseudoporphyria),这两种均可食,市场上的草鸡
:冠以此俗名的有两类:一类是鹅膏菌属(Amanita)的隐花青鹅膏(A.manginiana)和假褐云斑鹅膏(A.pseudoporphyria),这两种均可食,市场上的草鸡 主要指以上两种。另一类为丝盖伞属(Inocybe)的黄丝盖伞(I.fastigiata)和黄褶丝盖伞(I.flavobrunnea),又称“草帽菌”或“鸡
主要指以上两种。另一类为丝盖伞属(Inocybe)的黄丝盖伞(I.fastigiata)和黄褶丝盖伞(I.flavobrunnea),又称“草帽菌”或“鸡 花”,为有毒种类,曾见于中国云南中部市场;⑤麻姆鸡(
花”,为有毒种类,曾见于中国云南中部市场;⑤麻姆鸡( ):为鹅膏属的中华鹅膏(A.sinensis),见于中国云南省普洱市市场。⑥剥皮鸡
):为鹅膏属的中华鹅膏(A.sinensis),见于中国云南省普洱市市场。⑥剥皮鸡 :指丝膜菌属(Cortinarius)的近似多色丝膜菌(Cortinarius cf.variecolor)。
:指丝膜菌属(Cortinarius)的近似多色丝膜菌(Cortinarius cf.variecolor)。
同物异名的典型例子如块菌属的印度块菌(T.indicum)。在中国西南的川滇地区,该种的名称较多,分别为“松毛茯苓”,表明该种生于松林下;“无娘藤”,表明其子囊果与其周围的生物找不到直接的联系;“猪拱菌”表明经猪拱之后可被发现的菌;“煤黑”指子囊果的颜色为似煤般的黑色(陈娟,2007)。
结论:真菌分类学的未来之路
真菌分类学研究需要长期积累。如前所述,菌类标本的新鲜特征是分类鉴定的重要参考。真菌分类学家必须经历长期的野外工作才能全面掌握众多类群的新鲜特征。同时,从事真菌分类学研究的人还需要熟练运用显微镜,才能对不同的显微特征做出正确的判断和分析。只有经过长期积累才能正确把握并针对不同的类群记录、观察、分析不同的特征。
与植物分类学研究成果相比,真菌分类学无论是研究广度还是深度都相对不足(Hawksworth,2001)。绝大多数的真菌分类群缺乏全球性专著。在世界的很多地区,特别是菌类物种最丰富、最独特的热带地区,真菌分类学的研究依然很少且分散(Hawksworth,1998)。对这些地区的标本进行鉴定时会发现,很多标本无法找到合适的名称,因为很多种类尚处于未被描述的状态。因此在很多情况下便只能采纳相近种的名称甚至错误的名称。在估计的150万种真菌中,已描述的仅7.4万~12万种,而平均每年新描述的种仅为1100个(Hawksworth,2003)。尽管分子生物学研究手段为真菌的鉴定和分类提供了便利,要对庞杂的真菌种类进行准确的认识,还需依靠全球真菌学家的通力合作和长期努力(Hawksworth,2003)。
参考文献
Arora D.1986.Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi(2nd edition).Berkeley CA: Ten Speed Press.
Baxter,A.P and van der Linde,E.(1999) Collecting and preserving fungi: Amanual for mycology,SAFRINET/BioNet-International,National Collection of Fungi,Pretoria,South Africa,www.spc.int/pps/SAFRINET/fung-scr.pdf,accessed 10 June 2010.
Chang TT,Chou WN.1995.Antrodia cinnamomea sp.nov.on Cinnamomum kanehirai on Taiwan.Mycological Research 99(6): 756-758.
Chiu WF.1948.The Boletes of Yunnan.Mycologia 40(2): 199-231.
Corner EJH.1967.A monograph of Cantharelloid Fungi.New York: Oxford University Press.
Courtecuesse R,Duhem B.1995.Mushrooms&Toadstools of Britain&Europe.Italy: Harper-Collins,406-407.
Guzmán G.2008.Hallucinogenic mushrooms in Mexico: an overview Economic Botany 62(3): 404-412.
Hall I,Stephenson SL,Buchanan PK,Wang Y,Cole ALJ.2004.Edible and Poisonous Mushrooms of theWorld.Christchurch: New Zealand Institute for Crop&Food Research Limited.
Hawksworth DL.2005.Reflections on changing names and related nomenclatural issues in edible and medicinalmushrooms.International Journal of Medicinal Mushrooms 7: 29-38.
Hawksworth DL.1998.Getting to grips with fungal biodiversity in the tropics.In Chou CH,Shao KT(eds).Frontiers in Biology: The Challenges of Biodiversity,Biotechnology and Sustainable Agriculture.Academia Sinica.Taipei.
Hawksworth DL.2001.Themagnitude of fungal diversity: the 1.5 million species estimate revisited.Mycological Research 105(12): 1422-1432.
Hawks worth DL.2003.Monitoring and safeguarding fungal resources world wide: the need for an international collaborative Mycoaction Plan.Fungal Diversity 13: 29-45.
Hongo T,Izawa M.1994.Yama-kei Field Books Fungi.Tokyo: Yama-kei Publishers.
Imazeki R,Otani Y,Hongo T.1988.Fungi of Japan.Tokyo: Yama-kei Publishers Co.Ltd.
Jiang Y,Yao YJ.2002.Names related to Cordyceps sinensis anamorph.Mycotaxon 84: 245-254.
Jones EBG,Whalley AJS,Hywel-Jones NL.1994.A fungus foray to Chiang Maimarket in
Northern Thailand.Mycologist8(2): 87-90.
Jordan P,Wheeler S.2005.The practical Mushroom encyclopedia.London: Hermes House.
Kirk PM,Cannon PF,David JC,Stalpers JA.2001.Dictionary of the Fungi 9th edition.Wallingford: CABI Publishing.
Laessφe R.2002.Mushrooms.New York: DK Publishing.
Maruyama T,Kawahara N,Yokoyama K,Makino Y,Fukiharu T,Goda Y.2006.Phylogenetic relationship of psychoactive fungibased on rRNA gene for a large subunit and their identification using the TaqMan assay(II).Forensic Science International 163(1-2): 51-58.
Miller OK,Miller HH.2006.North American Mushrooms.Guilford: Globe Pequot Press.
Moncalvo JM,Vilgalys R,Redhead SA,Johnson JE,James TY,Aime MC,Hofstetter V,Verduin SJW,Larsson E,Baroni TJ,Thorn RG,Jacobsson S,Clemencon H,Miller OK.2002.One hundred and seventeen clades of euagarics.Molecular Phylogenet Evolution 23: 357-400.
Moser M.1983.Keys to Agarics and Boleti.London: Roger Phillips.
Pérez-Moreno J,Martínez-Reyes M,Yescas-Pérez A,Delgado-Alvarado A,Xoconostle-Cázares B.2008.Wild Mushroom Markets in Central Mexico and a Case Study at Ozumba.Economic Botany 62(3): 425-436.
Petersen R H,Zang M.1989.Ramaria Subgenera Ramaria and Laeticolor a in Yunnan.Acta Botanica Yunnanica 11(4): 363-396.
Phillips R.2006.Mushrooms.London: Pan Macmillan.
Ryman S,Bergius N,Danell E.2000.Proposal to conserve the name Armillariamatsutake against Armillaria nauseosa(Fungi,Basidiomycotina,Tricholomataceae).Taxon 49: 555-556.
Singer R.1986.The Agaricales in Modern Taxonomy.New York: Hafner Publishing Co.
Stalpers JA.1993.Keys to the species of the Thelephorales.Studies in Mycology.No.35.
Sung GH,Sung JM,Hywel-Jones NL,Luangsa-ard JJ,Shrestha B,Spatafora JW.2007.Phylogenetic classification of Cordyceps and the clavicipitaceous fungi.Studies in Mycology 57: 5-59.
Wu SH,Ryvarden L,Chang TT.1997.Antrodia camphorata(“niu-chang-chih”),a new combination ofamedicinal fungus in Taiwan.Botanial Bulletin of Acadia Sinica 38: 273-275.
Wu SH,Yu ZH,Dai YC,Chen CT,Su CH,Chen LC,Hsu WC,Hwang GY.2004.Taiwanofungus,a polypore new genus.Fungal Science 19(3-4): 109-116.
布里得森.黛安,福门.伦纳.1998.标本馆手册.克佑:克佑皇家植物园.
陈娟.2007.中国块菌属的系统分类学研究.中国科学院研究生院博士学位论文.
今关六也,本乡次雄,1957.原色日本菌类图鉴(I).Osaka:保育社.
今关六也,本乡次雄,1965.原色日本菌类图鉴(II).Osaka:保育社.
刘吉开.2004.高等真菌化学.北京:中国科学技术出版社.
刘培贵.1993.介绍一种滇产珍贵食用菌——玉蕈离褶伞.中国食用菌12(2): 29.
刘作易,梁宗琦,辛智海.2003.冬虫夏草显微结构再观察和子囊孢子发育研究.贵州科学21(1-2): 51-68.
刘锡进,郭英兰,佘永信,等.1989.多虫夏草无性阶段的分离和鉴定.真菌学报,8(1): 35~40.
王向华,刘培贵.2002.云南野生贸易真菌资源调查及研究.生物多样性10(3): 318-325.
王向华,于富强,刘培贵.2004.云南野生商品真菌图鉴.昆明:云南科技出版社.
杨祝良,2005.中国真菌志第二十七卷鹅膏科.北京:科学出版社.
应建浙,赵继鼎,卯晓岚等.1982.食用蘑菇.北京:科学出版社.
应建浙,卯晓岚,马启明,宗毓臣,文华安.1987.中国药用真菌图鉴.北京:科学出版社.
臧穆,苏庆华.1990.我国台湾产灵芝属一新种——樟芝.云南植物研究12(4): 395-396.
中国科学院微生物研究所真菌组,1979.毒蘑菇(第二版).北京:科学出版社.
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











