



7.1.1 Cu,Zn-SOD的基因特征
7.1.1.1 hCu,Zn-SOD的基因特征

图7-1 hCu,Zn-SOD氨基酸序列及相应的核苷酸序列
Cu,Zn-SOD基因位于21号染色体长臂2区1带(21q21),基因组DNA长11kb,含有5个外显子,编码甘氨酸、赖氨酸等153个氨基酸,组成一个33~34kD的蛋白质——Cu,Zn-SOD。它属于金属酶,由两个亚基组成,每个亚基均含有一个原子Cu和一个原子Zn。它的活性不仅与蛋白质分子有关,而且还取决于金属离子。
近年来分子遗传学的研究已确认10%~15%家族性肌萎缩侧索硬化症(FALS)的发病与Cu,Zn-SOD基因突变有关。其遗传方式多数为常染色体显性遗传,少数为隐性遗传。
Cu,Zn-SOD基因突变后的产物——变异Cu,Zn-SOD,在氨基酸组成和结构上发生变化,如两个亚基的空间三维结构不对称,尤其是在铜的结合部位,而致酶的功能发生改变。在FALS中,变异Cu,Zn-SOD活性下降,清除超氧化物自由基的能力降低,导致毒性超氧化物自由基的聚集。也有可能变异Cu,Zn-SOD活性增强,导致生成过多的过氧化氢(H2 O2),而H2O2与金属离子,如铁离子反应则能形成毒性极高的羟自由基。因此,认为Cu,Zn-SOD功能改变是FALS致病机制。
不同的Cu,Zn-SOD基因突变似乎不影响FALS疾病的进程,如A4V突变平均病程为1~2年,E100G突变为4.7年,而H46R和G37R为18~20年。
近年来国内报道FALS的3个家系21个患者。1995年,宋永梅等报道一个家系共8个患者,遗传方式为常染色体显性遗传,外显率高。起病年龄为23~40岁,发病原因可能与重体力劳动有关。李洵华等报道一个家系6例患者,属常染色体显性遗传,有外显不全现象。1997年朱凯云等报道一个家系13例患者,亦属常染色体显性遗传。
7.1.1.2 粗糙链孢霉细胞溶质中Cu,Zn-SOD的基因特征
将一段编码完整的粗糙链孢霉的长4.8kb的Cu,Zn-SOD的限制性内切酶片段从基因库中提取出来并测序。该基因包含三个内含子,并且没有一个内含子和已经鉴定的人的内含子相同。对内含子的位置的分析验证了一个假说,即Cu,Zn-SOD基因的复制和融合是在附加的外显子编码的N-末端β-发夹结构和锌结合位点亚区后进行的。粗糙链孢霉基因的一个内含子对应蛋白质一段保守序列的114位氨基酸残基,而人基因也有这样的一个内含子对应氨基酸残基,但是位置不同,在118位上。这些内含子位置的不协调性说明了它们中的一个是近期插入的。粗糙链孢霉的第一个内含子包含一个序列和酵母细胞色素c的转录调节位点UAS1相似,也和酵母Mn-SOD的假设调节位点UAS相似。
一些研究发现在真菌、高等植物和动物的线粒体的膜间隙中也和细胞质中一样存在着Cu,Zn-SOD。粗糙链孢霉的线粒体和细胞质中提取的Cu,Zn-SOD具有相似的相对分子质量,但是它们的电荷不同。
克隆的超氧化物歧化酶对阐述超氧化物歧化酶的作用和进化机制有着重要的意义。对从粗糙链孢霉的细胞质中提取的Cu,Zn-SOD的分子克隆,核苷酸序列,基因中内含子的结构作了报道,并且讨论Cu,Zn-SOD在蛋白质进化中的作用以及粗糙链孢霉Cu,Zn-SOD基因多样性。
A.粗糙链孢霉Cu,Zn-SOD的基因序列、内含子、外显子
Cu,Zn-SOD的序列测定(如图7-1)——粗糙链孢霉中插入到pCN24质粒的完整的基因序列包括1023碱基对。这个基因编码的氨基酸和报道过的细胞质中的Cu,Zn-SOD的153个氨基酸有152个相匹配。在基因序列中第69位的是苏氨酸,但是在蛋白质序列中却是丙氨酸。产生这种现象的原因可能是试验用菌株的多态性决定的。在蛋白质进化中丙氨酸和苏氨酸经常互相替换,它们的三联体密码在单体的转换型突变中存在差异。在酵母和其他大部分真核生物中这个位点被赖氨酸占据。核苷酸序列也揭示了甲硫氨酸的起始密码子在氨基酸1的密码子缬氨酸的上游。这个位置在人、酵母和玉米中都是相同的。
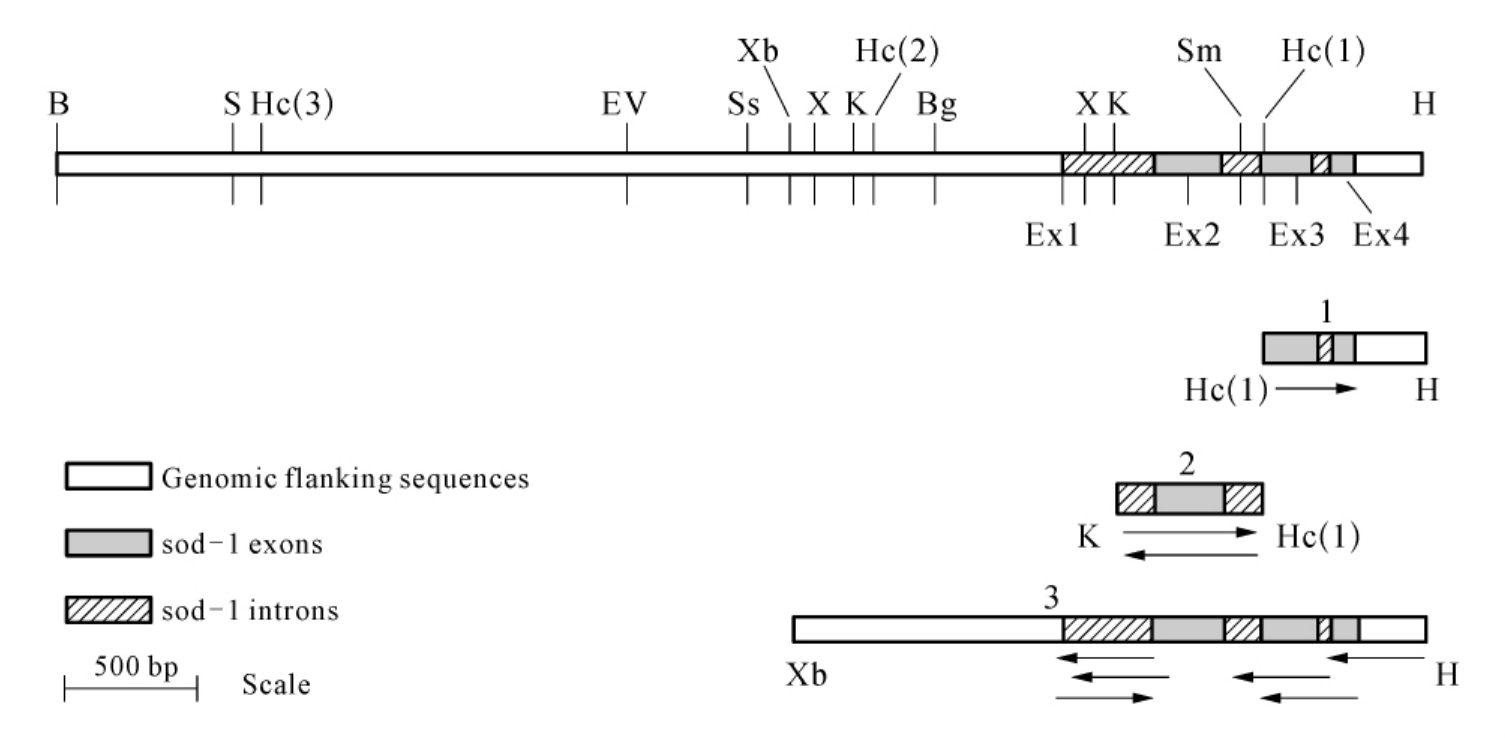
图7-2 pCN24的sod-1区的限制性图谱以及Cu,Zn-SOD基因的测序策略
(摘自DO Natvig,et al.J.Biol.Chem.,1990)
区域所示DNA插入在pBR325的BamHI和HindⅢ位点之间,片段1~3克隆到M13mp19中测序,箭头显示测序的长度和方向,限制性内切酶位点:B,BamHI;S,SalⅠ;Hc,HincⅡ;H,HindⅢ;Ev,EcoRV;Ss,SstⅡ;Xb,XbaⅠ;X,XhoⅠ;K,KpnⅠ;Bg,BglⅡ;Sm,SmaⅠ。
编码序列被三个已知的内含子所阻断,它们长度分别是287,100和57个核苷酸,并且所有的5′,3′端都被标记出来,具有粗糙链孢霉典型的内部专一序列。第一个内含子是目前报道的粗糙链孢霉最长的一个。这三个内含子将外显子隔开,分别在13,180,152和117个碱基处。
靠近第一个内含子的序列和酵母的转录调节位点UAS1的一部分相似。细胞色素c和酵母中Mn-SOD基因的UAS很相似。有一个证明就是HAP1的调节蛋白能够在有氧的条件下结合UAS1来诱导CYC1的表达,它还能调节酵母中Mn-SOD和过氧化氢酶基因的表达。举例来说,酵母中Mn-SOD基因上游序列和细胞色素c的UAS1很相似,据报道HAP1突变体中Mn-SOD缺乏氧的诱导性。研究这段序列在粗糙链孢霉细胞溶质Cu,Zn-SOD转录调节中的作用。文献报道酿酒酵母和粗糙链孢霉的调控元件是保守的,换句话说调节蛋白的结合位点与氨基酸的生物合成途径有关。
对10种不同的粗糙链孢霉sod-1的240区域上游和内含子的同源性进行了调查。一个10碱基序列ACCTCTTTGG包含UAS1相似区域的一部分并且和粗糙链孢霉2号内含子con-10基因高度匹配。这个con-10基因是在分生孢子受精作用中表达的,但是它的功能目前还未知。
B.sod-1基因的密码子偏爱性
sod-1基因显示了一种密码子的偏爱性,这似乎是粗糙链孢霉的一个特性,像am,bm1和细胞质tRNA合成酶基因。这种偏爱性在密码子第三个位置的嘧啶表现强烈,sod-1基因对以嘧啶结尾的密码子的偏爱性是96%,相对trp-1和his-3偏爱性是74%,这两个基因决定了核苷酸序列的偏爱性。sod-1只利用61个密码子中的32个,而trp-1/his-3普通片段只比sod-1基因长一点点却利用了50个密码子。最值得注意的是sod-1基因没有一个密码子是以A结尾的。

图7-3 粗糙链孢霉细胞溶质Cu,Zn-SOD的核苷酸序列及旁侧序列
(摘自DO Natvig,et al.J.Biol.Chem.,1990)
ATG起始密码子的核苷酸被标记出来,氨基酸从缬氨酸开始以斜字体表示出来,缬氨酸是成熟蛋白质的第一个氨基酸。这个编码序列被三个内含子阻断,它们的5′和3′内部一致序列下面用横线表示出来。箭头代表寡聚核苷酸序列的引物。探针SD1和SD2序列在真正的基因序列上面。错误匹配的碱基下面也用线标示出来。星号标记的启动ATG序列的5′在粗糙链孢霉是保守的,而第一个内含子的一段序列和2号内含子的一段序列很相似,也和酵母的CYC1基因的UAS1位点很相似。这个氨基酸序列和原来报道的氨基酸序列仅在69位不同,它的这个位置是苏氨酸,而原来的是丙氨酸。
G和C总共占四个sod-1外显子的67%,而内含子和侧翼序列则分别是51%和40%。对于粗糙链孢霉来说这个高的G和C含量是很普遍的,这可能主要是因为密码子的偏爱性。
7.1.1.3 原核细胞发光菌leiognathi中Cu,Zn-SOD基因特征
A.原核细胞发光菌leiognathi中Cu,Zn-SOD基因的氨基端前导肽
发光菌leiognathi的SOD基因是第一个被分离和测序的含有前导肽的Cu,Zn-SOD,相对于真核细胞的Cu,Zn-SOD定位在胞质,这一前导肽的存在可推测该SOD定位于细胞膜或者周质,这也证明了Cu,Zn-SOD的适应性,同时可以解释为什么典型的真核形式的Cu,Zn-SOD能够在原核细胞中发现。
图7-4表示出该含有前导肽的Cu,Zn-SOD核苷酸序列及氨基酸序列。从与其他基因的相同序列的比较可推测出它的调控序列,富含嘌呤的17~23位的SD序列:AAGGATA,-10到-35位序列:TTTAAT和TCAGCTAT分别为-7到-12位及-33到-40位。3′终止区包含简单的转录终止位点:残基561~585的前10个和后10个核苷酸形成的自身互补的茎以及中间断开的5个核苷酸形成的环,随后的是4T残基(587~589)的AT富含区的起始位点。

图7-4 发光菌leiognathi中Cu,Zn-SOD基因和读码框的核苷酸序列
(摘自Howard M S.J.Biol.Chem.,1987)
转录起始位点的核苷酸的编号为1;星号置于每第10个核苷酸残基的上方。核苷酸指定为假定的-35和-10序列,下画线表示出转录的终止径环结构,前导肽的氨基端的Met和成熟肽氨基端的Gln也以下画线表示出。
B.原核生物发光菌leiognathi的密码子偏爱性
表7-1列出原核细胞发光菌leiognathi中Cu,Zn-SOD中编码基因的使用情况:64个密码子中有17个从未被使用过。从而也说明了原核生物发光菌leiognathi的密码子偏爱性是不同于真核细胞的。
表7-1 原核细胞发光菌leiognathi中Cu,Zn-SOD基因中密码子使用情况

(摘自Howard M S.J.Biol.Chem.,1987)
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











