



爬行类动物的形成和进化
脊椎动物从水生到两栖类的转变,是动物进化的重要转折。但是两栖类并没有完全摆脱对水的依赖。两栖类还要在水里产卵,还要在水里度过幼年时期,如青蛙。青蛙的卵要在水里发育,小蝌蚪也只能在水里生活。
脊椎动物由水生到两栖类后,经过漫长的进化才完全摆脱了对水的依赖,进化为爬行类。由两栖类进化为爬行类,首先是从生殖方式的进化开始,羊膜卵的出现才标志着两栖类进化为爬行类。鱼类和两栖类的卵结构简单,只有在水里才能防止卵里水分的蒸发。而羊膜卵结构复杂,是在体内受精的。受精的羊膜卵产在地面或其他适当的场所,都能利用一定的环境自行孵化,有的在母体内孵化。把卵产下后再孵化的是卵生,在体内孵化出幼体再生的称为胎生。由卵生到胎生,这是动物进化的一个重要标志,也是由两栖类向爬行类进化的一个重要标志。
羊膜卵和两栖卵有本质区别,鱼类的卵和两栖类卵属于非羊膜卵,由于结构简单,只能在水里卵化,而羊膜卵结构复杂,可以在陆地卵化。长期生活在陆地的两栖类,由于陆地生活的影响,使生理结构发生了一系列变化,特别是卵里羊水为胚胎发育提供了一个生存环境,卵就慢慢地离开了水而能发育。体内受精卵又比两栖类和鱼类的体外受精卵更能很好地生存,两栖类的生殖功能的进化为生物进化到爬行类提供了必要条件。
一、爬行类的特征
爬行动物对陆地生活的适应及其原始特点,主要有以下几方面:
1.具有陆上繁殖的能力,其特点就是体内受精和产大型的羊膜卵。爬行类都产大型的羊膜卵,卵内贮备丰富的卵黄,保证胚胎在发育中有足够的营养;卵外被有坚韧的(革质或石灰质的)卵膜,起防止内容物流散、水分蒸发、机械损伤的作用。羊膜腔内充满羊水,胚胎得以在液体的环境中发育。
2.皮肤角质化,结束了皮肤呼吸。爬行类的皮肤一般被覆一层干燥的角质鳞,以阻止水分大量散失,从而开始了完全用肺呼吸,并出现了增强肺呼吸的胸廓。肺呼吸的完善,也促进了血液循环,心脏和动脉弓的进一步分化,混血程度大为降低,使它们可以摆脱潮湿的环境而生活在陆地上。
3.结构和机能进一步完善。爬行类骨化程度很高,五指型附肢和带骨进一步发展,并与支持身体的中轴骨联系得更紧密。颈部的出现和最前两枚颈椎的特化,使头部活动自如,而不牵连身体的其他部分。神经系统的发展,新脑皮的出现,动物的主动活动性大大增强,代谢水平也显著提高。
二、爬行类的出现
生存环境的变化,是古爬行类出现的一个明显的外界因素。石炭纪末和二叠纪初,地球上又发生了强烈的造山运动,陆地各部分向上隆起,地形比过去变得更为复杂。随之而来的是气候的剧烈变化。石炭纪时期那种单一的气候,变成各式各样的气候条件,干燥和寒冷成为当时很多地区气候变化的特点。三叠纪初干燥更为严重,广大的石炭纪森林、沼泽、湖泊都大大地减少,代之而起的是耐干耐寒的裸子植物森林和成片的沙漠。根据植物化石上的年轮,推知当时已出现了寒冷的冬季。气候上的这些改变,显然不利于两栖类的生存,致使很多古代两栖类绝灭或再次入水。只有那些长期经受陆地生活锻炼,在干旱的气候条件下产生了新的适应,并能产生羊膜卵的古两栖动物,逐步进化为原始的爬行类。
爬行类最早出现于石炭纪的晚期,到了二叠纪已成为常见的动物,到中生代几乎遍布当时的海洋、陆地和空中,所以中生代也被称作爬行类时代。早期的两栖动物和早期的爬行动物间的过渡是很明显的,化石蜥螈就是一个比较典型的中间类型,是长约1米的蜥蜴形动物。它的头骨在外形和结构上都和其他坚头类很相似。如肩带紧靠头骨之后,颈特别短,脊椎的分化不明显,有耳裂和连齿等,这些都和古两栖类相似。然而蜥螈已出现了爬行类的特征,如头骨有单个的枕髁,肩带具发达的间锁骨,有两枚荐椎,指的排列和指节数也和爬行类一样,腰带和肢骨都较粗壮,适于爬行。正由于蜥螈兼有两栖类和爬行类的双重特征,因此古生物学家中,有的认为它是两栖类,有的则认为它是爬行类。实际上是两栖类和爬行类的过渡。
蜥螈这样的位置是进化上最好的证据,因为它阐明了类群间的连续。关于爬行类的起源问题一向争论不休,人们都根据自己已知的化石材料来追溯爬行动物的种族发生。由于蜥螈的分类地位不定,因此那些认为它是原始爬行类的人们,长期地把它作为爬行类的祖先,并表示后来各类群爬行动物都是由它辐射进化而来的。然而化石材料表明:两栖类进化为爬行类发生在石炭纪晚期,而蜥螈却发现于二叠纪初的地层。由此看来,蜥螈不可能是爬行类的祖先。由于古爬行类门类繁多,形态、生态各异,除了主张它起源于一个祖型者外,也有人认为它有两个祖型或多个祖型。不过,尽管看法很多,然而爬行类作为一大类群来说,它们无论在解剖上、生理上以及生态上都与鱼类、两栖类有相同的特点,而这些基本的共同性也正是它们彼此间的亲缘所在,因此单源说还是容易为人们接受。目前一般认为爬行类起源于石炭纪前的两栖连齿类中的蜥螈形类,它们是结构轻巧、特化很少的原始类型。

蜥螈
三、爬行类的进化
杯龙类是爬行纲进化的主干。因为这类动物的椎体内凹,像一只只杯子,所以叫杯龙类。与其他爬行纲类群的主要区别是头骨不具颞孔,故称无颞孔类。

林蜥
最早的杯龙类化石是发现在北美洲加拿大新斯科舍上石炭纪下部地层里的林蜥,个体小,只有三四十厘米长。稍后的有在美国新墨西哥州下二叠纪地层里的湖龙,个体比较大,大约有1米多长;还有在美国得克萨斯州下二叠纪地层里发现的大鼻龙,个体小,长也只有30厘米左右。它们都属于杯龙类中的大鼻龙形类。它们的头骨构造上有许多两栖类的特征,头盖坚固,头骨后部是截平的,上下颌很长,上面有许多锋利尖锐的牙齿,看出它们是肉食性的,依靠捕食小型的两栖类过活。它们在许多解剖性质上和现代蜥蜴有点相似,在行动方式上也差不多。
从大鼻龙形类,到二叠纪晚期分化出另外的一支杯龙类,叫前棱蜥,也是小型的,也和现代蜥蜴相似,从晚二叠纪生活到三叠纪。和前棱蜥亲缘关系比较近的有锯齿龙,是一类比较粗笨的大型爬行类,有二三米长,是植食性的,主要生活在晚二叠纪。
另外有一类叫阔齿龙类,也是在美国得克萨斯州下二叠纪地层里发现的,骨架表现了原始杯龙类的特征,所以有人认为也应该归在杯龙类里。
杯龙类特别是大鼻龙形类被认为是后期发展起来的爬行动物的基干,从它分化出许多其他类型的爬行动物,是继往开来的代表。它本身除其中一支前棱蜥生活到三叠纪外,却都在二叠纪末就绝灭了。
中龙类出现在晚石炭纪,只在南美洲巴西南部和南非洲两个地方找到过它们的化石。中龙是一种小型细长的爬行动物,上下颌伸长,颌上有长而锋利的牙齿,肩腰带比较小,四肢纤长,脚变长成为宽阔的桡足,有一条长而灵活的尾。这些形态特征说明它是水生的,一般认为是淡水爬行动物,靠吃鱼类和其他水生小动物生活。
中龙类有许多特征是属于杯龙类的,它的椎弓是膨大的,和杯龙类的很相似,所以很可能它和杯龙类有共同的祖先。由于它特化程度较高,只代表爬行动物中一个很古老而独立的进化分支。
中龙类只生活在晚石炭纪到早二叠纪,很快就绝灭了。
盘龙类的化石发现于北美洲,特别是美国的得克萨斯州、俄克拉何马洲和新墨西哥州的上石炭纪和下二叠纪地层里。
盘龙类的头骨在许多方面和大鼻龙类的十分相似;脊椎骨有间椎体;四肢和杯龙类相似,但是比较细长。
盘龙类似乎也是从大鼻龙形类分化出来的。它本身只生活在晚石炭世到早二叠纪。但是从它发展进化而成的、生活在二叠纪中、晚期和三叠纪的兽孔类,却和哺乳类相似,也是哺乳动物的祖先。所以兽孔类也叫似哺乳动物,仍然属于爬行类。
始鳄类的化石最早是发现于美国堪萨斯洲上石炭纪地层里的岩龙,其次是发现在南非二叠纪地层里的杨氏鳄。
岩龙是一种没有特化的爬行动物,形态和习性有点像蜥蜴,身体和四肢都细长,适宜在地面上快跑。杨氏鳄也是结构轻巧的小型爬行动物,能在地面上快跑。
始鳄类是中生代称霸的爬行动物——恐龙类的最早祖先。但是它们一直生活到新生代开始,虽然始终不十分繁盛,却活到比它的后代恐龙类还晚。
“龙”原是我国古代虚构的一种神异动物,鹿角、牛鼻、蛇身、鸟爪、有须、有鳞,认为它能兴云作雨,并且象征着封建帝王。

龙
现在我们把一些古代的爬行动物叫龙,和这种虚构的动物是不同的。只是因为那些古代爬行动物和虚构的“龙”有点相似,我国的古生物学家就借用这个字来命名那些古代的爬行动物了。在拉丁文的学名里,许多古爬行动物名字后面都带有sauria,意思是蜥蜴。因为这些爬行动物的形状和现代的蜥蜴相似,蜥蜴本身就是一种爬行动物。
“龙”在我国古代也的确和一些爬行动物联系在一起,比如把蛇叫“龙衣子”,把一种蜥蜴类的爬行动物叫做“石龙子”。这就是说,在我国古代虚构龙这种动物时,也吸取了爬行动物的某些特点。
前面讲的都是早期的爬行类。随后无颞孔类在进化中通过辐射分化,产生出无孔亚纲、下孔亚纲、调孔亚纲和双孔亚纲等类型。下面简单介绍一下这四个亚纲。
第一亚纲无孔亚纲,头骨的眼眶后面没有颞孔。它包括杯龙目、龟鳌目、中龙目。杯龙类和中龙类是两类早期的爬行动物,龟鳌类是现存的爬行动物,它们在爬行类中属于比较低级的种类。
第二亚纲下孔亚纲,头骨每侧有一个侧颞孔,位置靠下,在耳窝外缘的鳞骨和后眼眶的下方。它包括盘龙目、兽孔目。盘龙类是早期爬行动物,已经绝灭。由它发展到兽孔类。兽孔类就是似哺乳动物,虽然本身也已绝灭,却由它发展出哺乳动物。
第三亚纲调孔亚纲,头骨每侧有一个上颞孔,位置靠上,在鳞骨和后眼眶骨的上方。它包括原龙目、楯齿龙目、蜥鳍目、鱼龙目。原龙类也是从杯龙类早期发展出来的一支。蜥鳍类包括幻龙类和蛇颈龙类。
第四亚纲是双孔亚纲,头骨每侧有两个颞孔,由鳞骨和后眼眶骨把它们分隔开来。它包括始鳄目、啄头目、有鳞目、槽齿目、鳄目、蜥臀目、鸟臀目、翼龙目。这一亚纲包括中生代的各种龙和现存的爬行动物蜥蜴、蛇和鳄。
下面我们就说一说这四类爬行动物的进化情况。
(一)无孔类和它们的残存后裔——龟鳌类
无孔类中的杯龙类和中龙类,它们都在二叠纪到三叠纪绝灭了。
无孔类爬行动物中,现存的只有龟鳌类。
龟鳌类构造特殊,和其他爬行动物都不相同,有人甚至提出应该把它另辟一类叫副爬行动物,和真爬行动物相区别。
真正的龟鳌类出现在中三叠纪或晚三叠纪,样子已经和现代龟鳌类相差无几。最早的祖先类型叫原颚龟。以后又出现两栖龟,也是一种原始的类型。它的躯体有坚固的甲壳保护,颈部很短,头部不能缩进壳里,或者只能稍稍收缩一点;头骨数目已经减少,牙齿已经从颚骨边上消失。到侏罗纪,从两栖龟分化出两个分支:一类叫侧颈龟,它的颈能向两侧方向弯曲,纳入壳里;另一类叫曲颈龟,也叫隐颈龟,它的颈能曲成S形直缩入壳里,是龟鳖类中比较成功的一类。

原颚龟
龟鳌类看来也是杯龙类的后裔。在它们的整个进化历史中,头骨的数目虽然已经减少,但是仍然趋向于保持原始爬行类那种坚固的头骨,在许多进步的龟鳌类中,头盖骨又有开孔和退化现象。另一方面,龟鳖类在长期生存斗争中又出现各种有用的适应。它是现存爬行动物中没有牙齿的一类,颚骨上的牙齿消失了,形成了角质的喙嘴,这种喙嘴对切割肉类和植物同样有效。肢体变得强壮,陆生种类足短趾少,海龟的足变成桡足,适宜于游泳。而真正的特化是甲壳的发展,肋骨分化发展包裹了肢带和肢骨的上节,来支持保护性的骨质背甲。在腹面发生了骨质腹甲。脊甲腹甲都覆盖有角质甲套,在两侧互相连接,使它们成为完全装甲的爬行类。龟鳌类就是这样取得了笨重的保护适应,尽管牺牲了灵活性,却能经得起时间的考验,一直延续到现在。
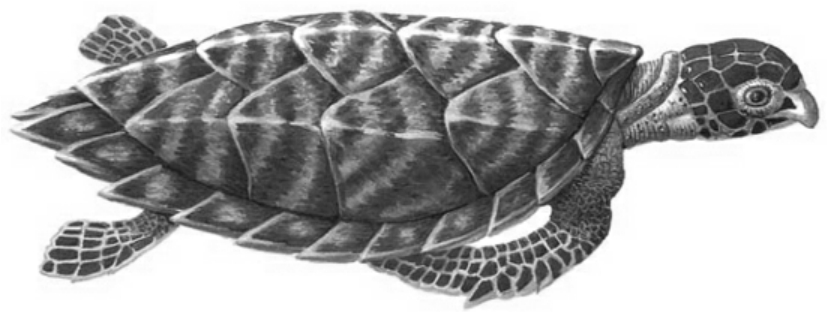
玳瑁
现存龟鳖类大约400多种,多分布于热带和温带,适应各种生活环境,如河流、沼泽、森林、沙漠等。在白恶纪,有一些龟鳌类回到海里,成为海龟,如玳瑁、绿龟等。有的长1米以上。
(二)下孔类爬行动物和似哺乳动物
最初的盘龙类叫蛇齿龙类,其中有:早二叠纪的巨蜥虫,是一种中等大小的爬行动物,体长1.5米,具有蜥蜴般的体形;二叠纪的蛇齿龙,是一种比较大的爬行动物,体长1.5米到2.4米。它们都是些吃鱼的动物,主要栖居河流、池塘边。
蛇齿龙类进一步向两个方向发展:一支发展成为巨大、凶猛、陆生、肉食性的楔齿龙类;另一支发展成为巨大的、植食性的基龙类。
到中二叠纪和晚二叠纪,盘龙类发展成为兽孔类,一直延续到三叠纪。兽孔类化石在世界各地都有发现,而南非的平原发现的更多。我国云南、四川、新疆、山西、内蒙古等地也是著名的兽孔类化石发现地。兽孔类就是似哺乳动物。
兽孔类主要的一支是兽齿类,它的特征:不像一般爬行动物那样牙齿不分化,全是一个类型的,而是分化成门齿、犬齿、颊齿,颊齿上还长有齿类。这和哺乳类——兽类相同,所以叫兽齿类。原始兽齿类叫丽齿兽类,早期进步的兽齿类叫兽头类。有一类叫包氏兽类,是特化了的类型。兽齿类中更重要的是犬齿兽类,晚期的进步的兽齿类可以犬齿兽作为代表。它生活在早三叠纪,身体最大有一头大狗那么大。它的头骨较大,长而窄;眼后有扩大了的颞孔,可容纳非常强大的闭合下颌骨的肌群;上颞骨也扩大了,下颞骨中的齿骨发展成为很大的一块骨头;牙齿已经高度分化。从犬齿兽类发展成为三列齿兽类。我国云南禄丰上三叠纪地层里发现的卞氏兽是世界闻名的,构造特征已很接近哺乳动物,只是下颌仍然具有爬行动物的特征,由一块以上的骨头组成。它属于兽齿类中的三列齿兽类。兽齿类中还有一类叫鼬龙类,构造特征上更接近哺乳动物。鼬龙类中有一种叫双关颌兽,发现在南非上三叠纪地层里,它在上下颌之间同时有两组关节。原来爬行类的上下颌之间由关节骨和方骨相连接,而哺乳类是由下颌骨和鳞骨相连接的。双关颌兽却既有爬行类的关节骨和方骨关节的残余,又有哺乳类的下颌骨和鳞骨的直接关节。
不过这些发现在晚三叠纪的似哺乳动物并不就是哺乳动物的直接祖先,因为在发现它们的同一地层里,已经多次发现了真正的哺乳动物,可知哺乳动物的出现在它们之前,是从它们的祖先类型中分化出来的,它们只是原始哺乳动物的堂兄弟罢了。
(三)调孔类爬行动物
调孔类爬行动物中最原始的是原龙类,是从杯龙类中早期发展出来的一支。
原龙类出现在二叠纪,延续到三叠纪。
二叠纪的原龙类是小型的、形状像蜥蜴的爬行动物,是陆生的,可能生活在灌木丛里,伺食昆虫或其他小爬行动物。到三叠纪,原龙类向不同的方向特化。在三叠纪结束时,原龙类趋于绝灭。
在三叠纪,从原龙类沿三个独立的方向发展,它们的代表是楯齿龙类、幻龙类、蛇颈龙类。它们都是海生的爬行动物。楯齿龙类生活在早三叠纪,它们特化为浅海里生活的爬行类,靠吃海底介壳类动物过活。楯齿龙类的结构笨重,身体粗壮,头骨、颈部和尾部都短,四肢骨中等长度,四足是比较小的挠足。背部脊椎上方有一排骨质的瘤状突起。腹面有坚固骨棒组成的腹肋筐。牙齿特化,前排成水平的,像是很有效的钳子,后排成宽大的磨石状,可压碎坚实的海生介壳类。楯齿类随着三叠纪的结束而绝灭。
幻龙类与楯齿龙类是同时代的海生爬行动物,也只生活在三叠纪。
这是一些从小型到中型的长形爬行类,有很长的可弯曲的颈,有发达的腹肋筐,四肢变长,相当强壮,四足是短的桡足,能爬上陆地。从颌骨和牙齿观察,它们显然是靠吃鱼类生活。
蛇颈龙和幻龙同属调孔类中的蜥鳍目,蜥鳍目爬行动物也叫鳍龙类。蛇颈龙是大型进步的鳍龙类。
蛇颈龙基本继承了幻龙的型式,只是躯体增大。它的样子活像一条蛇套在一只乌龟壳里,头很小,颈很长,躯体宽短而扁平,四足是很大的肉质挠足,能快速划动,且能迅速转身。它的上下颌骨有所改进,是凶残的肉食类,不仅吃鱼,也吃自己的幼仔和其他海生爬行类。

蛇颈龙
蛇颈龙类从晚三叠纪开始出现,身体逐渐扩大;到侏罗纪已遍布世界,身长达3米到6米;到白垩纪末,身长达到最长,可达18米。
蛇颈龙在侏罗纪和白垩纪的水域里称霸一时,到白垩纪末绝灭了。在这个时候海生爬行动物都趋于绝灭,只有属于无孔类的海龟仍然留在大海里。
(四)双孔类爬行动物
双孔类爬行动物可分为两大类:一个次亚纲叫鳞龙次亚纲。包括在石炭纪就出现的始鳄目,以及到现在还有后裔的两个目,一个叫喙头目,一个叫有鳞目。有鳞目包括蜥蜴类和蛇类。另一个次亚纲叫初龙次亚纲。包括一个最早的初龙类槽齿目,一个到现在还有后裔的鳄目,两个已经绝灭的恐龙目——蜥臀目和鸟臀目,一个已经绝灭的空中爬行动物翼龙目。
槽齿类在二叠纪末出现,到三叠纪末绝灭。它的历史虽短,种类也不多,但它却是统治中生代的主要爬行动物恐龙类的祖先。
喙头类出现在三叠纪初期,在三叠纪曾经繁盛过一时,分布到全世界。三叠纪以后,分布就受局限。
现存的喙头类爬行动物只有一种,叫槟齿蜥或咏头蜥,残存在新西兰附近的少数岛屿上,故叫新西兰蜥蜴。它的头部还有松果孔,表明它还保留有原始的形态,被看作是一种活化石。
有鳞类包括蜥蜴和蛇。蜥蜴和蛇在颚骨和翼状骨上有发育完好的牙齿。蜥蜴类从三叠纪晚期开始,到侏罗纪就已经沿着各种不同的适应路线辐射发展,且以后一直保持着这一特色。白垩纪曾经发展出某些巨蜥类,体长达9米以上,适应于回到海里去生活,如海王龙,属于沧龙类,短期繁盛后到白垩纪末绝灭。
现代蜥蜴类大约3800种,是现存爬行动物中种类最多的一个类群。
蛇类是所有爬行动物中最后进化形成的,实际是高度特化了的蜥蜴。现今蛇类大约有3000种,仅次于蜥蜴的一个类群。它们生活在地球上的许多地方。大部分生活在森林、草原、荒漠和山地里,少量生活在树上、地下和水里。
鳄类最早出现在三叠纪末期,从原鳄发展到早侏罗纪的中鳄,从早侏罗纪到白垩纪十分繁盛,继续到新生代初。在白垩纪,从中鳄又发展出两支比较进步的类型:西贝鳄类和真鳄类。前者已经绝灭了。真鳄类就是现代的鳄类。白垩纪和第三纪,真鳄类的分布范围比现在广泛。现代鳄类只分布在热带和亚热带地区,种类不多,只有大约25种,但它还是现存爬行动物中身体最大和凶猛的一个类群。因为鳄皮可以制革,现代一些鳄类被大量猎杀,有绝灭趋势,已经引起科学家关注。
四、恐龙类的活动和灭绝
双孔晋纲的蜥龙目和鸟龙目俗称为“恐龙类”,它们按骨盆位置的不同分别起源于三叠纪的槽齿目。三叠纪晚期是这两大类恐龙崛起的时代。
在现代的动物分类学里,蜥龙目一共分为三个亚目,这就是兽脚亚目、古脚亚目、蜥脚亚目。在兽脚亚目里又分虚骨龙次亚目和食肉龙次亚目;在蜥脚亚目里又分板龙次亚目和古蜥龙次亚目。
乌龙目一共分四个亚目,是鸟脚亚目、剑龙亚目、甲龙亚目、角龙亚目。
蜥龙类虽然腰带像蜥蜴,但和蜥蜴的亲缘关系并不太近,它是槽齿类的后裔。
最早的蜥龙类以北美洲的腔骨龙作代表,发现在新墨西哥州北部上三叠纪地层里。这是一些小型的两足行走的恐龙,骨骼中空,结构轻巧,身长2米左右,体重不过20多公斤。它的后肢强壮,形似鸟腿,善于奔跑,前肢短小,有如灵活的“手”适于攀援和掠取食物。它的身体以臂部作为支持点,后面有长大的尾和躯体前部保持平衡;颈部比较长,能弯曲;前端是尖狭的初龙式的头骨,颌骨长,装有锐利的锯齿状的槽齿,说明它是肉食性的。腔骨龙属于虚骨龙类。
虚骨龙类是蜥龙类中的兽脚类的祖先类型。兽脚类的特点之一是肉食性,二是始终保持两足行走的方式,属于蜥龙目的兽脚亚目。兽脚类从侏罗纪向各个方向进化,其中有的发展成为大型、凶猛的肉食龙。
肉食龙出现在晚三叠纪,到侏罗纪开始繁盛。永川龙是到现在为止我国发现的最完整的肉食恐龙化石。永川龙是在我国四川永川县上侏罗纪地层里发现的,全长约8米,高大约4米。它的头大而高,呈三角形,头上有六对大孔,其中眼孔最大,说明视力很好;嘴里长一排利剑般的牙齿;颈部较短,身躯不长,但尾部比较长,是奔跑时的平衡器官;前肢短而灵活,爪又弯又尖,像鹰爪;后肢长而粗壮,用三趾着地,类似现代的涉禽。永川龙能迅速追捕同时代的行动迟缓的植食性恐龙,追上以后猛扑过去,用前肢将对手推倒在地,用后肢牢牢地踩住,然后用利剑般的牙齿撕开猎获物的皮肉。
肉食龙发展到白垩纪达到鼎盛时期。白垩纪的肉食龙中著名的有惧龙、霸王龙。白垩纪末,它们都绝灭了。
蜥脚类是一类巨大的植食性的、生活在沼泽湖泊里的恐龙,属蜥龙目中的蜥脚亚目。我国在四川宜宾马门溪的上侏罗纪地层里发现的马门溪龙,由我国古生物学家杨钟健命名叫建设马门溪龙,也属于蜥脚类。特别在四川合川县境内发现的另一具马门溪龙,叫合川马门溪龙,化石较完整,身长22米,高3.5米,体重四五十吨,在亚洲发现的数它最大。巨大的蜥脚类也在白垩纪末全部绝灭了。
乌龙类全部是素食性。植食性使乌龙类的牙齿趋于退化。它的颌骨常具喙的构造,且在下颌骨前方有一个分离开的没有牙齿的骨头,叫前齿骨,作为喙嘴的下面部分。这是乌龙类的最显著的特征之一。乌龙类为了防御肉食龙的袭击,长期适应的结果,发展出各种光怪陆离的防身设备:甲板、棘刺和角。这些装备给恐龙世界增添了奇异的色彩。
除乌脚类两足行走外,其余三类全为四脚行走,且前肢总比后肢细小。早期的乌脚类个体不很大,而后期的代表如我国白垩纪的青岛龙,则身长6米多,山东龙可达15米。剑龙类个体不很小,7米左右,但脑子却只有核桃大,最大特征是背上有两行三角形骨板。我国的剑龙,有四川晚侏罗纪的嘉陵龙、沱江龙等。甲龙类和角龙类个体相对较小,生活时代都仅限于白垩纪。在我国,甲龙亚目有宁夏的绘龙,角龙亚目有内蒙的原角龙等。
过去一直认为,恐龙类是冷血动物。现代有资料表明,至少一部分恐龙属温血动物。恐龙的姿态暗示着某种敏捷性;与现时的温血和冷血四足动物比,它们的骨骼隐含着热血性的导管。从捕食者和被食者的比例的化石记录亦可说明,恐龙获得了很多食物。这一现象与温血类的哺乳类相似,反而与冷血的爬行类不同。
恐龙出现于2亿年前的三叠纪中期,绝灭于6700万年的白垩纪末,在地球上曾独霸约1.4亿年之久。可以说中生代的水、陆、空都是它们的天下。但到了中生代末期,它们已都突然走上了绝灭的道路。恐龙绝灭的原因,尽管古生物学、地质学、生物学、物理学、天文学等工作者都进行了研究,但至今尚无定论。在这方面主要有两类学说。一类是渐进说,另一类是突变说。前者认为,恐龙发展的年代,气候温和,有许多沼泽地和浅海。后来,气候变冷,陆地扩大。这种变化到6500万年更为突出,从而引起许多动、植物的死亡,恐龙即在这种条件下逐渐趋于绝灭。从时间上说,大体持续300万年之久。在这时期,小型的爬行类,如蜥蜴和蛇还能存活下去,而躯体巨大的恐龙,就难以存活了。大多学者都持有这种看法。另一类是突变说,如美国学者阿尔瓦雷斯提出的小行星冲撞地球而导致恐龙突然死亡的学说。六千五百万年前,有一行星与地球相撞,使地球的大气充满灰尘。有几个月时间,昏天黑地,日月无光。植物因光合作用停止而普遍死去,食物链中断,恐龙也因此而绝灭。这个学说的依据是,在中生代白垩纪末期意大利、丹麦、新西兰等处的地层中,发现大量的金属铱,约比地球正常含铱量高出30倍,而高含量的铱只有在太阳系的其他行星上才有。支持突变说的人数不多,但其影响很大。此外,还有不少别的解释,甚至还有人认为恐龙根本就没有绝灭。总之,到目前为止,这个问题仍在争论之中。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










