



摩尔根与他的果蝇实验
866年9月25日,摩尔根生于美国
肯塔基州的列克辛顿。从小他就对生物颇感兴趣。1880年,他进入肯塔基州立学院学习生物学,1886年获得动物学学士学位。摩尔根的研究兴趣极为广泛,他一生的科研工作总是在生物的进化、遗传和发生等广阔的领域里不断地交换着研究课题。在1910—1935年期间,他集中研究果蝇的遗传问题。在1935—1945年期间,他研究胚胎学及其与遗传和进化的关系。
摩尔根不仅对于果蝇的遗传学研究最负盛名,同时他对于胚胎学、细胞学以及进化论的研究也都作出了显著贡献。为了表彰他在创立染色体遗传理论方面的功绩,摩尔根被授予1933年度的医学或生理学诺贝尔奖金。摩尔根的功绩不仅是在染色体遗传理论方面和胚胎学方面所取得的重大成果,而且在实验方法上,他首次把定量分析方法应用于解决生物学问题,这就促使遗传学很快地有了飞跃的发展,并为现代的新兴科学——基因工程奠定了基础。
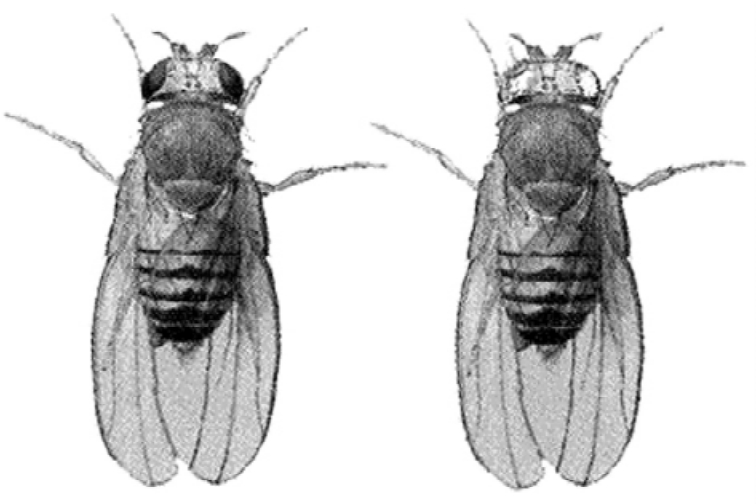
白眼雄蝇(右)与红眼果蝇
摩尔根是于1908年前后开始养殖果蝇的。1910年,他在一个培养瓶里,偶然在一只雄果蝇身上发现了一个细小而明显的变异:一般的果蝇都是红眼,而那只变异的雄果蝇却是白眼。摩尔根让这只白眼雄果蝇与红眼雌果蝇进行交配,结果所得到的后代都是红眼。在子一代中进行兄妹交配而产生子二代,其中发现了一些白眼果蝇。摩尔根惊奇地注意到,所有的白眼果蝇几乎绝大部分出现在雄性身上,偶尔也会出现一只白眼雌果蝇。
摩尔根把白眼果蝇的出现,称为性别连接,意思是说,白眼基因是被连接在性染色体上的。后来,摩尔根的性别连接观点被证明可以适用于一切有性生殖的生物体,并能说明许多其他模糊不清的遗传类型,包括人的红绿色盲和血友病。摩尔根的果蝇研究成果,第一次揭示出一种或多种遗传特性与某一特定的染色体的明确联系。
总之,摩尔根发展了孟德尔的遗传学理论。他在以果蝇为材料进行的实验遗传学的研究中,发现了伴性遗传的规律。他和他的同事们在发现连锁、交换和不分离现象等的基础上,发展了染色体遗传学说,认为染色体是孟德尔式遗传性状传递机理的物质基础。
摩尔根进一步创立了基因学说,认为基因是组成染色体的遗传单位,并且证明基因在染色体上占有一定的位置,而且呈直线排列。他还认为,在个体发育中,一定的基因在一定的条件下,控制着一定的代谢过程,从而体现在一定的遗传特性和特征的表现上,基因还可以通过突变而发生变化。他在遗传学方面的著作有《基因论》《孟德尔式遗传机制》等。
摩尔根于1945年12月4日逝世于美国加利福尼亚州的帕萨迪纳。
下面我们就来细说这个白眼果蝇的故事。
果蝇这种实验材料是1908年在纽约冷泉港卡内基实验室工作的卢茨向摩尔根推荐的。这是一种常见的果蝇,学名称为“黑腹果蝇”。
实验材料的选取往往是决定研究工作成功与否的关键,它在遗传学发展史中表现得尤为突出,不仅是在摩尔根选用果蝇,还有孟德尔选用豌豆,以及后来分子遗传学家们选用真菌、细菌和噬菌体都生动的证明了这一点。可以说,遗传学发展史中,每一次实验材料的选取都导致了一次学科发展的飞跃。以哺乳动物为实验材料,饲养管理一般都较复杂,生长期又长,而且由单基因控制的性状少而难寻,所以,一般不适合遗传学理论研究。这也许就是遗传学基本定律首先从植物中发现的主要原因吧。而果蝇体型小,体长不到半厘米,饲养管理容易,既可喂以腐烂的水果,又可配培养基饲料,一个牛奶瓶里可以养上成百只。果蝇繁殖系数高,孵化快,只要1天时间其卵即可孵化成幼虫,2-3天后变成蛹,再过5天就羽化为成虫。从卵到成虫只要10天左右,一年就可以繁殖30代。果蝇的染色体数目少,仅3对常染色体和1对性染色体,便于分析。作遗传分析时,研究者只需用放大镜或显微镜一个个地观察、计数就行了,从而使得劳动量大为减轻。
在野外采集到的果蝇,眼睛都是红色的,称为“野生型”。1910年5月,摩尔根在实验室里饲养的一群红眼野生型果蝇中,发现了一只白眼果蝇。摩尔根独具慧眼,立刻认识到这只白眼果蝇的巨大价值。晚上他把这只珍贵的白眼果蝇带回家,放进床边的广口瓶里之后,才安心睡觉,白天再把它送回实验室去。在实验室里,摩尔根使这只白眼果蝇与尽可能多的野生型红眼雌果蝇交配,十天后产生了1240个子裔,形成了一个庞大的果蝇株系。
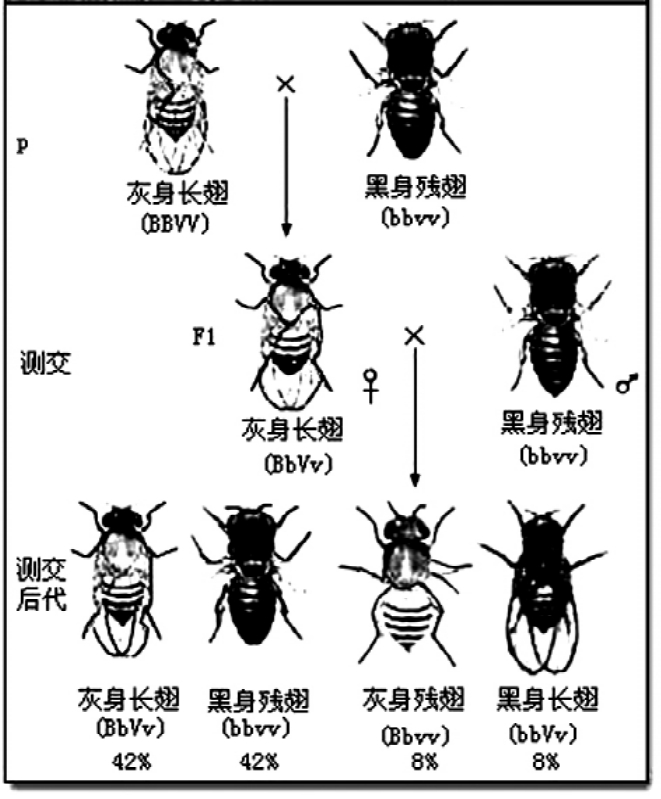
白眼雄蝇与红眼果蝇杂交,子一代全是红眼果蝇。子一代自交,子二代的结果完全是孟德尔式的,其中红眼果蝇2688只,白眼果蝇728只,两者比率约为3.4∶1,而约占1/4的白眼果蝇则全是雄性个性。摩尔根的这一结果,以“果蝇的有限性遗传”为题发表在1910年7月22日出版的《科学》第32卷第120页上。如果没有后面的结果,则摩尔根的发现只不过是孟德尔学说的又一例证,说明孟德尔学说也适用于昆虫。而后面的结果表明,白眼基因与性别有关,这里面就有新的东西了。摩尔根在论文中没有急于宣布眼色基因一定与性染色体相关联,只不过说,眼色基因的分离与两条性染色体的分离一致。他在该论文中的解释略显复杂,也存在一些细节上的错误,但结论是正确的。同年及翌年,摩尔根又连续发表了两篇论文,终于把基因与染色体的关系确定无疑地联系在一起了。
玉米的有性杂交实验
摩尔根指出:如果假定控制眼色的基因位于X染色体上,而Y染色体上则不带控制眼色的等位基因,那么实验结果就能得到完满的解释。红眼基因(+)是显性,带有红眼基因的X染色体用X+表示;白眼基因(w)是隐性,带有白眼基因的X染色体用Xw表示。基因型为XwY的雄果蝇,由于Y染色体上没有控制眼色的基因,隐性基因得以表现,所以是白眼果蝇。当白眼雄果蝇与野生型雌果蝇X+X+杂交,子一代的基因型是X+Xw和X+Y,即雌雄果蝇都为红色复眼,且雌果蝇是杂合体。子一代个体相互交配,结果是在子二代中有3/4是红眼果蝇,1/4是白眼果蝇。雌果蝇全为红色复眼,但其中有一半是纯合体,另一半为杂合体。雄果蝇则红眼、白眼各占一半。
前面已经谈到,将遗传物质与染色体联系在一起,在耐格里、魏斯曼时就早已有之,鲍维里、德弗里斯和萨顿甚至把这种联系描述得十分详细。但这些人所谈的联系都仅仅是推测,并没有将某一个具体的基因定位在某一条染色体上。摩尔根却做到了这一点,他把红眼等位基因和白眼等位基因定位在X染色体上,并用实验证实这些基因是由X染色体携带着遗传的,这就使基因在染色体上的假说有了坚实的基础,而且还是把一个特定的基因归属到一条特定的染色体上,更有甚者提出这条特定的染色体还与性别有关。
白眼果蝇在基因学说的发展史上起了不可估量的作用,以致引起关于最初那只白眼果蝇来历的争论。向摩尔根推荐果蝇作为实验材料的卢茨,曾出版过一本富有魅力的书《多样性的昆虫》。他在该书中声称:“摩尔根教授访问我们研究所时,我曾对他提及,在我繁衍的一个血统清楚的果蝇株系中,出现了一只白眼果蝇。但因忙于异常翅脉果蝇的研究而无暇顾及白眼果蝇。摩尔根要走了这只活的白眼果蝇并使之繁殖,终于使白眼果蝇在后代重新出现。我讲这些话,绝非自夸。如果我当时意识到那只白眼果蝇会成为价值无比的珍品,也许就不会把它赠送给别人了。然而,我是把它送给了一位杰出的研究者。实际上,黑腹果蝇应该称为摩尔根的遗传果蝇。”

玉米的套袋隔离
摩尔根不承认最初这只活的白眼果蝇来自卢茨,一个相信卢茨的读者在美国《遗传杂志》上发表对卢茨一书的书评,摩尔根立即作出了回答,但却是略带暧昧的回答。他写道,自己确实向卢茨要过果蝇的饲养品系,但是,其中并没有卢茨所说的白眼果蝇,卢茨虽然发现过一只白眼果蝇,但当时就是死的,也不包括白眼果蝇的先代。如果先代中带有白眼果蝇基因,虽然先代本身是红眼,但后裔中总有可能分离出白眼果蝇来。实际上,卢茨所赠的果蝇株系,其后代中未出过白眼果蝇。后来也了解,白眼是果蝇中最常见的一种突变型。“与应用这一突变型材料作出科学发现相比较,发现白眼突变型本身则并不是那么重要。”
上述摩尔根所说的最后这一句话无疑是对的,且不论最初那只白眼果蝇的来历如何,摩尔根以它作为实验材料,在遗传学史上第一次证明了基因位于染色体上,并且发现了伴性遗传规律。因此,这个白眼果蝇的故事不仅仅是一个戏剧性的插曲,而是值得大书特书的一项重大的研究进展,它开创了摩尔根基因学说的先河。摩尔根于1909年开始培育果蝇。他对德弗里的月见草突变的印象很深,并试图通过不同的化学药品、不同温度的处理以及钙和X线辐射在他所培养的果蝇中引起突变,但是都未成功。然而在他的果蝇系谱培养中有一个白眼雄蝇出现在红眼果蝇的正常种群中。
这一个简单的事实,在实验室培养中出现了单个异常的个体,却引起了一场名副其实的雪崩式的研究热潮。首先提出的问题是,这个“白眼”性状是怎样产生的。将这一罕见的白眼雄果蝇与同一代的雌果蝇交配,摩尔根发现虽然F1代都是红眼,但在F2代又出现白眼雄蝇,这表示白眼的遗传因子是隐性,它必然是由红眼基因骤然变化所产生的。摩尔根在几年前曾访问过德弗里在荷兰的实验室,也认为这新等位基因的起源采用了德弗里的“突变”这一术语。由于德弗里的进化突变学说以及月见草突变的染色体性质,这一术语的转移引起了不利的后果,在随后的二三十年中造成了一定的思想混乱。然而遗传学家和进化论者最后都习惯了摩尔根赋予“突变”这词的新含义。
在生物学的历史上还很少有像摩尔根与他的同事一道工作那样亲密无间。因此很难确定摩尔根实验室的大量研究成果或发现究竟应归功于谁。有些历史学家倾向于将几乎所有的功绩归之于他的学生和同事。这未免太过分。应当记住自从摩尔根于1910年7月发表他的关于果蝇研究的第一篇文章以后的两年中他连续发表了13篇有关20多个果蝇性连锁突变型的发现与行为的文章。在发现白眼后不久又发现了两个性连锁隐性突变型:“翅发育不全”,“黄色体色”。毫无疑问摩尔根在很早就阐述了孟德尔遗传的大部分机制,这些都是他本人所作的贡献。正如穆勒所说的:“无论将来会怎样重写和重新评价果蝇研究开始阶段的历史,摩尔根对交换现象的论证以及基因相距愈远交换也愈频繁的观点就像一声春雷,其重要意义绝不下于孟德尔学说的发现,这一点必然仍旧会被公认。”我在这里所要强调的是摩尔根本人单独地对连锁和交换问题作出的贡献,因为以后的讨论重点放在问题上而不是果蝇实验室中每个人的特殊贡献。摩尔根和他的同事在哥伦比亚大学的“果蝇室”中培养了成千上万的果蝇。他们在仔细审查这些果蝇中发现了新的突变不断出现。摩尔根很快从哥伦比亚大学的未毕业学生中挑选了斯特体范特和布里奇斯两人到他的实验室工作,后来穆勒也参加了这个研究小组。这个小组的亲密协作是生物学中的一段轶史佳话:“在科学实验室中过去很少有如此激动人心的气氛和如此持续的热情。这主要是由于摩尔根本人的态度,他将热情和批判精神融合在一起,大度、胸襟开阔并具有幽默感。”
在短短的几年之内传递遗传学的一切主要方面都被摩尔根和他的小组阐明清楚。凡是贝特森、德弗里、柯仑斯,凯塞尔以及其他早期的孟德尔主义者所没有找到的正确答案都由摩尔根小组出色地完成了。其所以如此,一个重要的原因是摩尔根虽然是胚胎学家出身,但是他将基因生理学和个体发生问题扔在一边,审慎地倾注全力于传递遗传学。他没有去臆测遗传定律而是探究事实和这些事实的最简单的可能解释。
一、等位基因
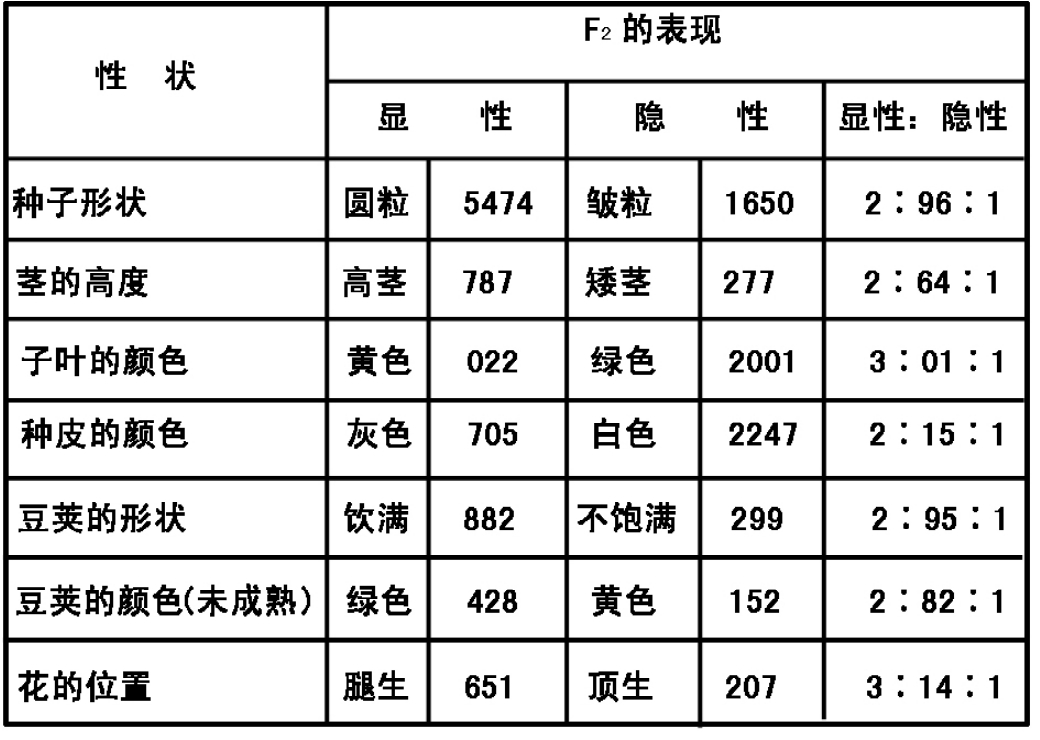
孟德尔充分了解表现型性状是成群出现,在他所选用的性状中则是成对出现。1900年以后的研究工作证实了对应于某个表现型状的物质基础可以有二者择一的表现形式或表达。将之直译成希腊文,这些二择一的决定因子就是相对基因或等位基因。孟德尔遗传中这样的一些表现型性状的二择一定子的发现便对变异的起因问题能作出完全新的解释。这使人联想起豌豆的光滑对皱皮、黄与绿,或者其他的类似成对性状可能有相同的物质基础。等位基因所表现的性状应当是同一遗传物质的两个译本。
1904年法国库恩奥在家鼠中发现一组性状可能有两个以上的等位基因。例如在家鼠这一特殊情况下其皮色可能是灰、黄、黑色。贝特森、凯塞尔、谢尔、摩尔根以及其他遗传学家后来也发现了这类复等位基因现象。人类的ABO血型就是人所共知的例子。斯特体范特首先解释了复等位基因现象,将之归因于同一基因的各种可供选择到状态。这完全否定了贝特森的基因作用的或有或无学说。在某些特殊情况下,一个基因有50个以上的等位基因,例如,牛的血型基因以及植物的某些亲合性基因、脊椎动物的组织亲合性基因。和孟德尔定律相符,在某一配子中永远只有单个等位基因,但在受精时它能够和存在于种群基因库中的许多不同的任何一个等位基因结合。在遗传学历史上后来还发现在某些杂交中基因的行为与等位基因相似,而在其他杂交中则并不如此。
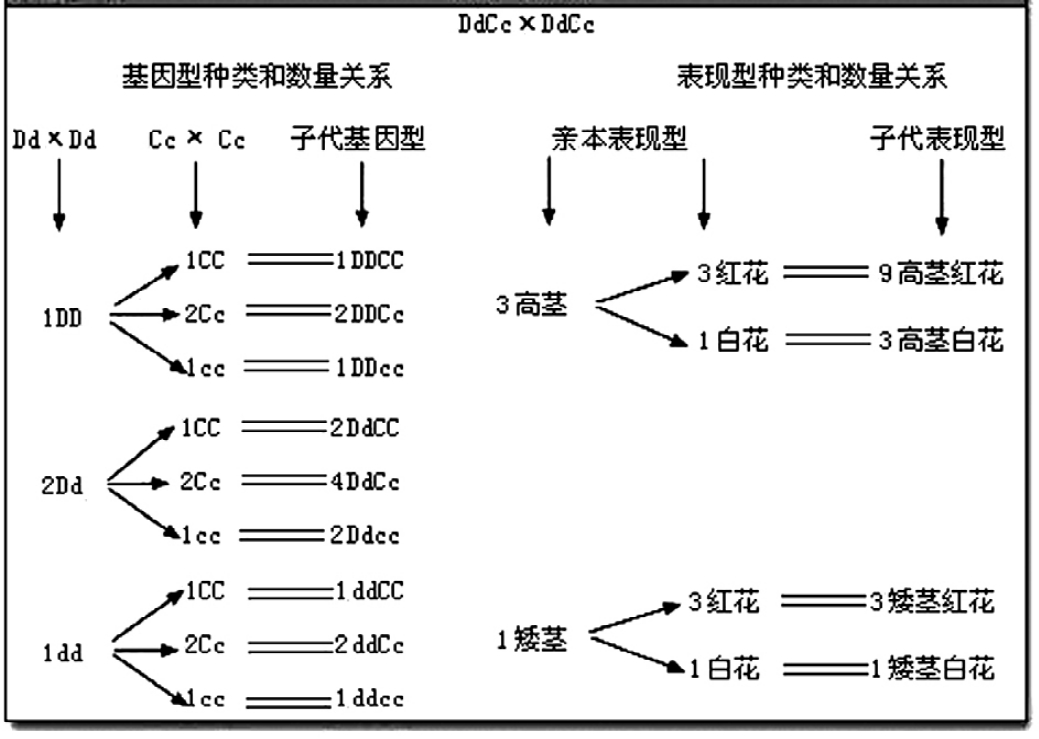
解答图表
摩尔根研究小组对果蝇的白眼基因以及果蝇的其他突变型的研究,明确地证明了一个基因可以突变成另一个等位基因,后者又能突变成第三、第四个等位基因。同样值得注意的是,这些突变步骤是可逆的,白眼果蝇偶尔也能产生红眼后代。也许最重要的发现是某个基因一旦产生了新等位基因,这新的等位基因就一直保持不变,除非其后代之一发生了新突变。因此基因的特点是,它的几乎完全稳定性。基因突变的发现并不是退回到软式遗传,恰恰相反,这证实了遗传物质是基本固定不变的。可以这样说,这是硬式遗传的决定性证明,即尽管遗传物质不仅具有内在的稳定性,然而它又具有进化演变所容许的突变能力。
很快就证实了一切其他生物都能突变,从人类和其他哺乳动物到最简单的动物、各种植物、甚至微生物。事实上从1920年直到1950年研究突变似乎是最有希望的、能阐明遗传物质本质的研究路线。突变过程的研究同时也提出了一些难于解答的问题。在突变时基因究竟发生什么变化?能不能在控制条件下产生变异?德弗里早在1904年就提到“X射验和镭射线都能穿透生物细胞,可以用来改变生殖细胞中的遗传颗粒”。自从1901年以后一直试图通过X射线,镭射线,温度骤变,或化学药品等办法诱发突变。由于各种技术上的缺陷,起初都没有得到明确结果。最后,通过穆勒百折不挠和独创的精神及倾注全部心血进行的研究一直到1927年才取得成功。
二、减数分裂
1902年以后,虽然有一些生物学家随意谈论染色体遗传学说,但是它的准确含义究竟是什么当时并不十分清楚。大多数人指的是茹提出的意见,即各个不同的遗传因子在染色体上的直线排列。然而这并不是问题的全部。细胞学家在1870年代到1890年代发现了许多肯定与遗传有关的染色体现象。1900年以后对这些现象进行了系统研究,尤其是摩尔根研究小组在1910年以后的研究,这大大促进了染色体学说的发展。
下面从染色体在配子形成时的行为开始介绍。卵子的细胞核和精子都是“单倍体”,也就是说它们具有的染色体数目是体细胞染色体数目的一半。在配子形成时染色体数目是怎样减成一半的而且这又怎样会影响遗传?
三、交换
由于任何生物的性状及决定这些性状的遗传因子远远超过它所含染色体的数目,所以从一开始就有人认为每个染色体必定携带有几个基因。这一点很快就被摩尔根研究小组证实。然而连锁群的发现又提出了新问题。如果一个染色体上的所有基因都紧密地连锁在一起,那么一个生物将会只有它所含有的染色体的数目那样多的独立遗传单位。这就会对重组施加极大的约束。德弗里在研究F2代杂种时所得出的结论是,F2杂种重组的可能机会远不符合全面连锁的观点。因此他提出在减数分裂的第一前期中,成对的亲本染色体发生“单位交换”。至于“有多少和哪一些则可能纯属偶然”,这表明交换总是相互交换。波弗利也曾预言过这类交换。遗传学分析很快就证明了同一个染色体上的基因连锁是不完全的。贝特森,桑德斯,庞尼特首先进行了这类观察研究。在香豌豆的两个变种杂交后的F2中,他们既没有得到预期的9∶3∶3∶1的比值,也没有发现简单的3∶1比值,却发现了69.5%的双显性个体,19.3%双隐性个体,以及两类5.6%杂合子。显然这两个性状的基因既不自由组合又非完全连锁。贝特森曾提出过一个特别假说来解释这一现象,但由于他不相信染色体学说,所以也就没有考虑交换。
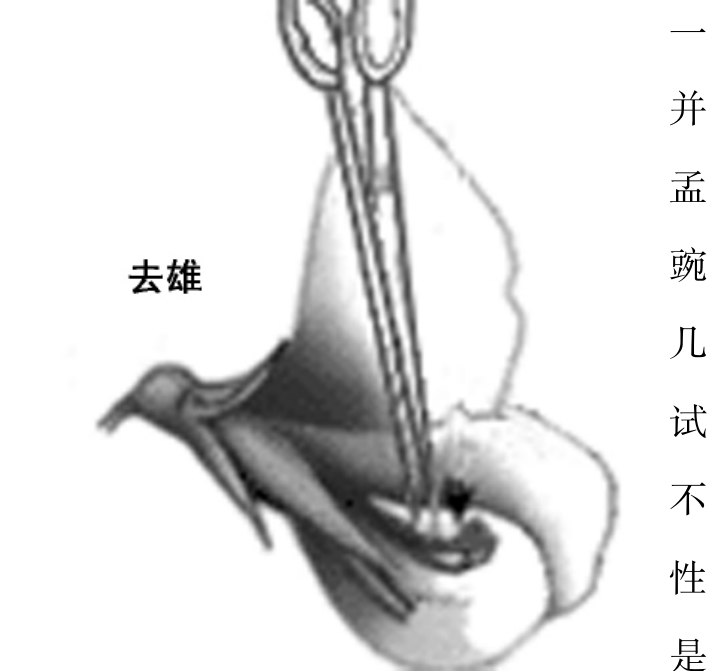
豌豆
过去常常有人提起孟德尔没有遇到连锁现象真是一件怪事。豌豆只有7对染色体,孟德尔也正好研究了7对性状。它们没有连锁起来是不是运气好从而为孟德尔减少了一项额外的麻烦?看来并不是这样。我们知道孟德尔在正式着手他的豌豆试验之前曾经花了几年时间进行预备杂交试验,他很可能将在几代不显示自由组合的一些性状弃置不顾。也可能是他由之取得试验材料的种子商偏爱自由组合的那些性状。最后,某些基因的基因图距足够大,即使是都处在同一个染色体上也能显示自由组合现象。
当摩尔根研究小组开始集中力量分析果蝇的遗传结构时,不符合完全连锁的例外情况成为一个严重问题。摩尔根及其同事发现,断裂连锁的幅度很大,有时可以低至1%,这样的变异性怎样解释?
举一个特殊情况为例。有一组三个隐性基因黄色,白眼,小翅位于果蝇的X染色体上。如果一个具有这三个基因的雄蝇和一个正常雌果蝇杂交,可以预料这三个隐性基因将以连锁群的形式在F2代中出现。实际上黄色和白眼的连锁在果蝇中有1.3%断裂,白眼小翅连锁有32.6%断裂,黄色小翅连锁有33.8%断裂。怎样解释这些数字?
这些例外情况的数值通常多是按德弗里提出的单位交换的随意性这种偶然过程来解释。但是根据1900年代早期的细胞学研究可以作出不同的答案。自从波弗利和赫特维克那具有开拓性工作以来的20年中,对减数分裂细节的研究已取得长足进展。在第一前期中染色体的变化至少可以分为六个阶段。其中有一个阶段两个成对的染色体仍然很细,但是每个染色体已分开成染色质丝,即所谓四线期。这两个染色单体彼此反复交叉形成波状环。
比利时细胞学家简森斯指出,当四个染色单体彼此盘绕在一起时,一个父本的和一个母本的染色单体可以在它们彼此交叉处断裂,断头在重新连结时总是父本单体的断头连在母本单体的断头处,反过来也是如此。另两个染色单体仍然保持完整。这样一来就形成了一个“交叉”,在减数分裂第一前期的后期阶段是成对的两个染色体保持接触的地点。按简森斯的观点交叉表示一个父本的和一个母本的染色体的交换。最后结果将是一个由父本和母本染色体的片段组成的新染色体。摩尔根小组所研究的不完全连锁和简森斯的观点相符。
交换过程是如此复杂以致差不多花了30年时间最后才能决定哪一种解释是正确的。然而现在已充分证实交换发生在四线期,涉及四个染色单体中的两个单体。另外还证实在四线期一开始就发生交换。
摩尔根和他的助手斯特体范特认为,源于交换的不完全连锁的份额表示遗传因子在染色体上所处位置之间的直线距离。染色体在两个基因之间断裂的机会取决于这两个基因在染色体上的距离,距离愈近,断裂机会愈少。根据这一推理斯特体范特推算了基因在染色体上的位置和顺序并制出了普通果蝇的X染色体的第一份染色体图。他由之证实了当时所知道的这一染色体上的基因是沿着染色体作线性排列的。
在早期的实验中有一些结果相互矛盾。穆勒指出,在一个长染色体上可能发生双交换,而且交叉的存在将干扰在染色体上与交叉邻近处的进一步交换。考虑到这两种新发现的现象就排除了上述矛盾,摩尔根的一些反对者正是利用这些矛盾来怀疑交换学说的正确性。
遗传现象的染色体学说现在已经可以用基因学说来补充。1915年前后摩尔根及其同事研究了一百多种突变型基因。它们分成四个连锁群,和果蝇的四个染色体非常一致。连锁群的染色体实质的间接证明至此便告完整。然而一直到1931年斯坦才运用某些异常基因为交换提供了细胞学证明。同一年克莱顿和麦克林托克在植物方面也提出了类似证据。后来玉米成了细胞遗传学的优良研究材料。虽然它没有后来在果蝇研究中非常有利的巨型染色体,但是它所含有的全部十个染色体在形态上都不同,而且有时还有额外的染色体存在。麦克林托克利用玉米的这些特点进行了30多年艰苦而又出色的研究来解释基因的作用。这一解释虽然内容丰富又独到见解,却一直等了很多年,在分子遗传学家得到了相同结论之后才被举世公认。
这里所介绍的有关交换现象的历史是过于简化的,漏掉了很多复杂问题。例如交叉的实质一直长期争论不休。每一染色体臂上的交叉数目极不一致,事实上在某些情况下并没有交换现象,例如雄果蝇。关于第一次减数分裂中染色体的确切复制时间以及形成交叉的确切时间,甚至交叉的存在是否总是表示交换现象都有很多争议。最重要的是,染色体中不同的染色单体的行为更是众说纷纭莫衷一是。因而作为解释交换现象的简森斯和摩尔根的断裂并合学说并不被某些学者接受,例如,贝林又另行提出了“副本选择”学说,维克里提出“基因转变”学说。虽然这两种学说最后都没有得到公认,却促使人们进行了大量试验从而对交换现象以及基因的本质有了更深入的了解。现在还没有这三种学说的比较研究历史著作。对这些技术上的详细情况必须参阅细胞学和遗传学教科书。重要的是,一切看来是例外的情况最终都能按经典染色体学说加以解释。
孟德尔做的豌豆杂交试验的结果
对进化过程来说由交换而实现的染色体重建非常重要。它是父本和母本基因相混合的有效机制,并通过产生基因在染色体内的新组合来提供非常丰富的新遗传型以便自然选择发挥作用。
染色体另外一个作用是能促进重组,即在成熟分裂的减数分裂中父本和母本染色体的独立运动。1902年以前普遍认为父本和母本的染色体组作为各自的单独单位运动。例如某些学者以为在卵细胞的成熟分裂时所有的父本染色体都以极体的形式被排除,然后通过受精由来自父本的新染色体组代替。如果真是这样,单性生殖卵在成熟时就不会产生极体,然而波弗利证明单性生殖卵不仅产生极体,而且极体形成的方式和有性生殖卵的没有什么不同。另外,杂合的雌体产生具有父本基因的配子。最后,克莱顿发现在具有大小不同的染色体组的物种中,较大的染色体随意向两极运动。这是父本的和母本的染色体组并不作为单一的单位进行分离的决定性证据。然而有一种罕见的遗传现象、“减数分裂驱动”,阻止染色体任意移向两极。这能说明在某些情况下种群中保存了在其他情况下有害基因的现象。
所谓天才人物,指的就是具有毅力的人,勤奋的人,入迷的人和忘我的人。
——木村久一
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











