



第三章 肺部疾病的免疫学基础
肺脏虽是气体的交换器官,但也是许多具有抗原性的异种蛋白质进入体内的重要途径,其中也包括许多病原体。虽然每天都可吸入有感染性的微生物,但健康的肺脏基本上还是一种无菌的环境,表明它必须具备能中和及消除有害微生物的有效机制。不过这些机制并不完全,而且很多呼吸道疾病的发生都是由于肺脏消灭病原体能力的缺陷所致,而在无免疫缺陷时,多数的肺部感染均为急性和自限性的。
肺脏除了可以接触到一些感染性的颗粒以外,尚可摄入为数甚多具有抗原性的惰性异种蛋白质,例如植物花粉、真菌孢子、哺乳动物和昆虫的蛋白质,如蟑螂和尘螨。这些物质往往与哮喘的发生有关。肺部就成为首次致敏和以后多种抗原物质攻击的场所。
为了对付肺脏所遭遇到的感染性攻击,如呼吸道病毒、细胞外和细胞内细菌、真菌和寄生虫,呼吸道中必须具备有效的免疫系统。此种免疫系统包括各个不同的组分,如物理屏障、天然免疫和获得性特异免疫等。
一、肺部的天然免疫
(一)物理性免疫
肺脏及上呼吸道具备许多在滤除颗粒物质中起重要作用的物理屏障。鼻道可以阻止大于10μm的颗粒通过喉部。咳嗽和作呕反射在防止分泌物、大的颗粒和胃肠内含物进入呼吸道时是绝对需要的。这些极为重要的基本防御机制的缺失,如在运动神经疾病或延髓麻痹中所见的神经疾患,即可引起因异物坠入而发生的肺部感染或吸入性肺炎,这些都是有生命危险的。一旦颗粒通过声带,则主要的物理屏障为黏膜纤毛屏障。从气管至最小的软骨支气管中的上皮细胞均由纤毛柱状上皮细胞所组成。上皮细胞和黏膜下组织中含有丰富的黏液腺。这些腺体能分泌黏液至呼吸道管腔中。在此,黏液附着于纤毛的末端,形成连续的覆盖物。这种黏液层通过纤毛的协同搏动,以5~10 cm/min的速度向外移动,然后被咳至喉部,并被吞入。每天约有2 ml痰液由正常呼吸道排出。黏液由混有细菌的分泌黏蛋白、营养物和废物所组成。黏蛋白是一种长的纤维肽,并富含复杂的O-连锁寡糖。已发现有8种黏蛋白基因,每一基因中均表现有相当多的多态性以及所分泌黏蛋白型别的变异。黏蛋白的理化性质及其特殊的黏度和弹性,在其对宿主的免疫作用上是很重要的,因为它可使盐类、离子和气体通过,而且可防止不需要的颗粒和微生物进入。如果黏度降低,则不能抵抗重力的作用而涌入肺泡;相反如果过于黏滞,如在囊性纤维化中所见的情况,则其就不能通过纤毛的作用而移动。黏度在很大的程度上取决于水分的含量,但类脂、蛋白质和离子含量也起作用。在很多的情况下这种黏膜纤毛装置的功能有缺陷。如吸烟有损于纤毛功能,增加黏液分泌并降低其黏度,使清除黏液困难,并导致反复性的呼吸道感染。有些不大常见的遗传性疾患,如Kartagener综合征,就是因纤毛功能障碍引起反复性感染所导致的支气管扩张。囊性纤维化时则有黏液黏度增加,这可能是疾病发生的部分原因。不过,高浓度盐类对防御素功能的影响也是其中的一种因素。由于支气管腺瘤或过敏性支气管肺曲霉病(ABPA)引起的支气管阻塞所伴有的支气管扩张,也是由于这种清除机制的丧失而发生的。
(二)抗微生物肽系统
天然免疫中的血浆蛋白组分,特别是补体系统,在呼吸道分泌物中并非以高浓度存在。但却有许多与天然免疫有关的肽类存在,它们多由白细胞产生,并分泌至黏膜表面以表现其抗微生物作用。
1.防御素 防御素是发现于所有品系生物,乃至果蝇中的最原始抗菌防御机制的一大类抗菌肽。根据其一级结构可区分为两种亚族,即α-防御素和β-防御素。目前已知在人类中有6种α-防御素和3种β-防御素。4种α-防御素存在于中性粒细胞的颗粒内,即人类中性粒细胞肽(HNP)1~4;另有2种α-防御肽则存在于小肠Paneth细胞(肠腺的嗜酸性粒细胞)。人类中的2种β-防御素(HBD)则广泛表达于上皮细胞,其中也包括支气管上皮细胞。HBD-1是固有表达的,HBD-2则是在炎症时诱生。最近尚发现有第3种对盐类不敏感、来源于支气管上皮细胞的β-防御素。防御素是分子量为(3~5)×103、对蛋白酶有抵抗力的分子,有3~4个二硫键,在人类中仅由β-折叠组成。其作用主要是通过导入类似通道的结构至细菌的细胞膜中。防御素在防御呼吸道感染中起作用,可能是因为它在肺部表现为盐类敏感抗生素的作用,但在囊性纤维化时则被灭活。防御素具有广谱的抗菌特性,包括对细菌、真菌、分枝杆菌、螺旋体的活性以及体外抗肿瘤细胞的作用。
2.乳铁蛋白(Lf) 乳铁蛋白是存在于乳汁、血液和白细胞中具有抗病原菌性质的肽类。它是蛋白质中转铁蛋白家族成员,并根据其所结合的铁质含量而有3种同工型存在。乳铁蛋白具有抗病毒、细菌和真菌活性,并且对某些肿瘤细胞有细胞毒作用。就与肺部的关系而言,它能抑制呼吸道合胞病毒(RSV)的生长,对流感嗜血菌和铜绿假单胞菌也有作用。此外还可抑制胰蛋白酶,但其在变应原攻击前应用时,尚可抑制绵羊中的晚期应答。
3.溶菌酶 这是在粒细胞和许多分泌液中所发现的一种高度阳离子化、分子量为14.3×103的肽类。它可降解革兰阳性细菌的糖类细胞壁;除非是革兰阴性细菌的外膜受到损伤,如被乳铁蛋白作用以后,一般对溶菌酶有抵抗力。
4.收集素 收集素(collectin)是C型外源凝集素的成员,有3股螺旋的类似胶原蛋白功能区以及外源凝集素功能区;且有类似胶原蛋白杆状的三维结构,在其C-末端则有其糖类识别功能区。此种三聚体结构有些可以形成复杂的分子。收集素包括表面活性剂蛋白A和D(SP-A/D),这些表面活性剂系由Ⅱ型肺泡细胞合成,并且在表面活性剂中发现。它们可通过其糖类功能区与细菌和病毒结合,从而有利于肺泡巨噬细胞的吞噬。
(三)天然免疫的细胞基础
1.NK细胞 自然杀伤(NK)细胞具有淋巴细胞的形态,但却有不同的受体表型;它们是CD3阴性、CD16(FcγRⅢ)阳性、CD56阳性、CD8阳性和CD4阴性。之所以称之为自然杀伤细胞,是因为他们能对某些型别的靶细胞表现细胞毒作用。其细胞毒作用有一部分是通过抗体依赖细胞毒作用(ADCC)来完成的,此时包被靶细胞的抗体与N K细胞上的Fc受体结合,从而促使两种细胞的黏附并激发细胞毒过程的发生。此种过程涉及穿孔素和粒酶的形成,并且与CD8+T细胞和通过Fas依赖性机制诱生凋亡所引起的细胞毒作用是相似的。一般认为N K细胞在对肿瘤细胞的细胞毒作用上很重要,但对肺部免疫方面的作用尚不清楚。N K细胞在肺组织中存在,但其数目较之CD4或CD8 T细胞为少。
2.粒细胞 中性粒细胞存在于正常人肺中,是痰液和支气管抽出液(BA L)中的主要细胞类型,提示它们能通过上皮细胞进入支气管管腔。BA L中的中性粒细胞生存时间不长可能是因为它们一旦离开了黏膜下组织后,很快发生凋亡。正常呼吸道中的中性粒细胞在每一基膜长度单位上,较之淋巴细胞要少10倍。在细菌性感染时,大量的中性粒细胞移入支气管固有膜肺泡间质,再通过吞噬作用和被抗体调理细菌的裂解而在宿主的防御方面起重要作用。支气管扩张时的呼吸道内和细菌性肺炎时的肺泡内都有大量的中性粒细胞存在。致命性的细菌性肺炎是粒细胞减少症的重要并发症。不适当的炎症反应而导致的中性粒细胞大量流入,可能引起有害的效果,如在急性的急性呼吸窘迫综合征(A RDS)和慢性的支气管扩张中所见者,此时过于激烈的宿主防御反应引起了大量的组织损伤。
嗜酸性粒细胞和嗜碱性粒细胞在正常肺中并不多见,但嗜酸性粒细胞可以是许多肺部疾病的明显特征,常与Th2型免疫应答有关,如哮喘时所见者。在蠕虫寄生虫疾病时亦可见到暂时性的肺部嗜酸性粒细胞浸润,此时常可见有通过肺部的移动期,如丝虫病(热带性肺嗜酸性粒细胞增加)。近年来,研究者深入研究了过敏性疾病中嗜酸性粒细胞进入肺部的分子机制,结果发现这主要是一种由细胞因子IL-4,IL-5和IL-13所控制的Th2淋巴细胞介导过程。与中性粒细胞相比较,嗜酸性粒细胞的吞噬能力较弱,一般是通过其所分泌的细胞毒介质,主要是碱性蛋白(MBP)的作用。
3.肥大细胞 肥大细胞大量地存在于肺部,密切靠近许多控制肺内部自稳过程的结构细胞,包括脉管系统、内皮细胞、神经组织和黏液腺。它们并不转移至呼吸道管腔,故在BAL或痰液中的含量极低。90%的肺部肥大细胞表现为黏膜表型,而不是结缔组织表型,故其仅表达类胰蛋白酶(tryptase),而不表达糜蛋白酶。与血液中的嗜碱性粒细胞相比较,肺部肥大细胞只表达较低水平的FcεRI受体,但结合于细胞的IgE与抗原的交联是它们被激活并释放出介质的主要途径。结果是对其功能的认识主要集中于其对哮喘的作用上。而在纤维化疾病,特别是外源过敏性肺泡炎时,它们可在BAL中被大量发现。肥大细胞是肺部组胺的主要来源,也可分泌大量的其他炎症介质,包括类脂介质PGD2。这一介质对肥大细胞和白细胞三烯C4有相对的特异性。白细胞三烯能刺激黏液分泌,并可引起支气管平滑肌收缩。近期研究证明,肥大细胞也可生成许多细胞因子,特别是与Th2型应答有关的细胞因子,如IL-4,IL-5和IL-13,但是也可产生TNFα。
4.肺泡巨噬细胞(A M) 肺泡巨噬细胞来源于骨髓,沿单核细胞系发育成为成熟的单核细胞而进入血流,在此循环24 h以上,然后移入肺部。在肺部,A M存在于肺泡间隙,黏附于肺泡上皮细胞以及小的和大的气道,并在黏膜层以及固有膜中发现。A M与外周血单核细胞在细胞形态、受体表型、吞噬能力和介质产生上都有所不同。至于A M究竟是来源于单核细胞的某一亚群,还是来源于整个单核细胞群体,目前还不清楚。有关A M移动的机制也是所知有限。不过,它们是一种长寿命的细胞,当其活化后,可以移动至淋巴结,只是详情未知。
AM可作为吞噬细胞而发挥其主要作用,能吞食吸入的颗粒、被调理的细菌以及衰老的细胞。与所有的白细胞一样,AM也可以各种不同的活化状态存在,由静止的细胞乃至完全被刺激的介质产生细胞。它们能产生许多介质,包括类脂(特别是LTB4)和趋化因子、趋化吸引剂以及许多细胞因子。特别是与Th1型免疫有关的细胞因子,如IL-1,TNFα,IL-12,IL-10,TGFβ。IL-6也可以因为AM的活化而大量产生。AM可因许多介质的作用而活化,特别是由革兰阴性菌产生的LPS和Th1淋巴细胞产生的IFNγ。虽然AM也可通过抗原呈递细胞而发挥作用,但多数的研究结果提示,它们能抑制T细胞应答,因而在防止对吸入抗原的不适当炎症反应上起重要作用。与此相一致的是,肺部炎症动物模型中AM的去除能促使炎症反应明显加强。这种抑制效应在动物中证明是由于NO的产生所介导的。AM的发育依赖于GM-CSF,并且在GM-CSF剔除小鼠中由于有AM功能失调而表现肺部免疫缺陷。这些小鼠由于表面活化剂代谢缺陷,可以发生肺泡蛋白沉积症(alveolar protinosis)。GM-CSF对AM功能的影响是由转录因子PU.1所介导的。
5.细胞因子和趋化因子 如同其他部位一样,肺部免疫系统受到免疫细胞和组织细胞所分泌肽类介质的控制。这些肽类介质总称为细胞因子,它们是一大类不同的分子,并可区分为许多基因家族,均与内稳功能和炎症反应有关。其主要的功能与生长因子、细胞活化剂和趋化吸引剂相同。它们是通过与细胞膜上的受体结合而表现其活性,并转导活化信号至细胞质。细胞是否对一特定的细胞因子发生反应,取决于它能否表达针对这一介质的受体。不过功能后果在不同的细胞类型之间有所不同,这取决于信号转导途径的不同,而这些转导途径则是由于受体与其细胞因子配体结合的结果,由受体转导信号至细胞质中的第二信使常是由于对其他蛋白质磷酸化方式的改变所引起的,这种过程可通过诱导酪氨酸或丝氨酸/苏氨酸残基(激酶)的磷酸化或脱磷酸化(磷酸酶)来完成。信号转导途径常对转录因子产生效应,引起mRNA产生和蛋白质合成的增加。
与免疫系统有关的两个主要细胞因子家族为白细胞介素和趋化因子。白细胞介素是原被称为淋巴细胞生长因子的异质性多功能介质,分子量约为20×103,其受体有两条链:一条链与白细胞介素结合;另一条链则与信号转导有关。信号转导链常与其他受体共有,例如IL-3,IL-5和GM-CSF,α-链各不相同,但是β-链则是三者共有。同样,被认为是IL-2受体一部分的γ-链可以和IL-4和IL-9结合,细胞因子的命名多按照定性的次序和原始的功能,而不是根据其结构上的特征。例如,IL-8是趋化因子家族中的一成员,而其他的生长因子和相关的细胞因子,如TGFα,TGFβ,TNFα,PDGF和VEGF则不包括于白细胞介素家族之内。与肺部免疫有关的一些白细胞介素和相关细胞因子的主要功能总结如表3-1所示。
表3-1 与肺部免疫相关的主要细胞因子

(续 表)

趋化因子是(8~10)×103家族的趋化吸引肽类,根据其一级结构上的共有特征,特别是两个保守性半胱氨酸残基的结合而区分为4个亚族。CXC家族中的半胱氨酸由一可变性的核苷酸隔开,并对中性粒细胞和T细胞有趋化作用。CC趋化因子家族中的半胱氨酸则是相邻的,对T细胞、单核细胞、嗜酸性粒细胞、嗜碱性粒细胞和树突状细胞都有趋化作用。另外尚有一CX3 C家族成员(Fractalkine)和XC家族成员(单一保守的半胱氨酸)(Lymphotactin),CXC趋化因子再可根据在半胱氨酸以前有无谷氨酸-亮氨酸-精氨酸残基而进一步区分。有这些残基者对中性粒细胞有趋化作用,而没有者则对淋巴细胞表现趋化作用。
趋化因子受体是7个穿膜结合G蛋白受体的大家族成员。它们均通过Gαi亚单位来转导信号,这一亚单位对百日咳毒素敏感,并可介导趋化作用和黏附过程中白细胞停滞的活化步骤。趋化因子中的一项重要特性是其对个别细胞类型由于趋化因子受体的差别表达不同所表现的特异性。例如,多数个体的嗜酸性粒细胞仅表达CCR3,因而只能对能与此种受体结合的趋化因子,如eotaxin发生反应。趋化因子受体在淋巴细胞上的高度异质性表达使得趋化因子在个体发育和致敏作用上都起主要作用,因为这两种过程都需要有淋巴细胞亚群的精密组织区室化(compartmentalization),即在发育时一系列由大的区室到小的区室越分越细的过程,以及炎症反应中的白细胞浸润程式。已有日渐增多的事实说明CCR3结合趋化因子在哮喘时引起肺部嗜酸性粒细胞浸润中的重要性,并且证明IL-8是介导中性粒细胞移入肺部的关键性趋化因子。很多趋化因子都是由肺部上皮细胞产生的。其中包括T细胞反应性趋化因子IP-10,Mig和能与CXCR3结合的I-TAC(表3-2)。
表3-2 趋化因子及其受体

6.白细胞向肺部的移动 白细胞向肺部组织中的移动能力在天然免疫和特异性免疫中都很重要。这种细胞移动在健康者的正常的肺部自稳过程以及炎症反应中都可发生。在炎症反应中,白细胞移动的力度和程式均有所不同。移入正常肺部的主要细胞为T细胞和单核细胞,这些细胞可以分化成肺泡巨噬细胞。NK细胞和中性粒细胞亦可存在,但嗜酸性粒细胞和B细胞则较少见。嗜碱性粒细胞在正常和炎症肺部均不多见。肥大细胞可能是以前体细胞形式进入肺部,然后就地分化。虽然在炎症时树突状细胞迅速移入肺部,但对其基线移动情况仍是所知不多。白细胞向炎症肺部的移动在以Th1介导的免疫应答为特征的细菌性感染时,多以中性粒细胞为主;而在以Th2介导免疫应答为特征的过敏性和寄生虫疾病时,则以嗜酸性粒细胞为主,但两者之间也有一定程度的重叠。在很多炎症疾病时也有T细胞和单核细胞的增加。
白细胞由全身循环向组织中的移动是通过毛细血管后静脉内皮细胞完成的,并且受到一些逐步发生相互作用的调节。白细胞首先连接于内皮细胞上,这一过程主要是选择素(selectin)家族所介导的,即内皮细胞上的P-选择素和E-选择素和白细胞上的L-选择素。这些受体结合至高度糖基化类似黏蛋白的配体[如P-选择素糖蛋白-1(PSGL-1)]后,即可在完全的应激条件下表现功能,当白细胞通过血管流动时,以较快的速度将其捕捉。白细胞一经连接于内皮细胞,就可沿血管表面滚动,直至其被表达于内皮细胞上的趋化吸引物所活化,并且引起黏附受体中整合素(integrin)超家族成员的功能性上调,然后与表达于内皮细胞上的免疫球蛋白家族黏附受体成员结合。因此,整合素α4β1(V LA-4)结合至VCA M-1;β2或CD18整合素LFA-1和Mac-1结合至ICA M-1和ICA M-2以及α4β7结合至MAdCA M。这样就可牢固地扣留白细胞,使其以后在局部生成趋化吸引物的影响下,直接移位至组织的各个部位。包括上述趋化因子家族在内的趋化吸引物系由多种细胞产生,特别是寄居的细胞群体,如支气管上皮细胞、成纤维细胞和平滑肌细胞。由此可见,白细胞的移动受到许多黏附受体和趋化吸引物的调节,从而发生多种多样的信号转导,使不同的白细胞亚群根据时间和刺激性质的不同而发生差异性移动。
T细胞和单核细胞基线移动至肺部的机制尚了解不够(见以下所述有关淋巴细胞向肺部归巢的调节)。对于肺部炎症疾病时细胞移动的机制则有较多的研究报告,特别是应用各种炎症刺激的动物模型以及应用单克隆抗体和基因剔除动物以确定白细胞移动的分子调节的研究。如上所述,肺炎和急性肺损伤时,中性粒细胞向肺泡实质的移动主要是通过毛细血管。在正常情况下,当中性粒细胞通过肺部毛细血管时必须变形,而在中性粒细胞活化时可引起细胞结构上的改变,从而防止细胞变形,并且引起中性粒细胞在肺部循环中与黏附受体的独特性螯合。当动物注射了补体片段,并引起白细胞减少时亦可观察到这种情况。动物经抗CD18或抗L-选择素抗体处理后,此种螯合作用就成为暂时性的,表明黏附受体与保持脉管系统的完整性有关。与全身循环时中性粒细胞移动大都取决于CD18整合素的情况不同,中性粒细胞通过肺部毛细血管到达肺泡至少只是部分不依赖于CD18。例如在CD18缺陷小鼠中,在注入了肺炎链球菌以后,中性粒细胞移入肺部的过程并不受损,而当注入大肠埃希菌、LPS或铜绿假单胞菌时,中性粒细胞的移动可降低约75%。E-选择素或P-选择素在中性粒细胞通过肺部毛细血管时都不起主要作用。对嗜酸性粒细胞移动的分子控制机制的研究则比较详尽,主要研究这种细胞在哮喘中的作用。总之,嗜酸性粒细胞的移动受到Th2细胞因子IL-5,IL-4和IL-13的协同调节。IL-5能使嗜酸性粒细胞生成增加和易于由骨髓中释出,IL-4和IL-13则可诱导内皮细胞上VCA M-1和P-选择素的上调,在此可形成嗜酸性粒细胞的螯合。IL-4和IL-13也可诱发上皮细胞和成纤维细胞生成的CC趋化因子的表达。这种趋化因子可以与表达于嗜酸性粒细胞上的主要趋化因子受体CCR3相结合。这两种细胞因子还可控制嗜酸性粒细胞移入呼吸道中的固有层。对单核细胞移动情况的研究不多。在大鼠模型研究中有一报告提出,气管内LPS注射时,约有30%的抗CD18抗体能抑制单核细胞移入肺组织,而抗VLA-4抗体则无作用。抗CD18抗体对中性粒细胞的移动亦无作用。两种抗体的联合作用则可抑制80%的细胞进入BAL。
二、肺部的特异性免疫
(一)一般概况
1.抗原识别 特异性免疫包括两大部分,即细胞免疫和体液免疫。前者主要由T细胞介导,故亦可称为细胞介导免疫(CMI),这是因为胸腺(thymus)在T细胞的个体发育上起重要作用。后者则是基于体液或抗体的免疫,抗体是由B细胞产生的。如同所有的白细胞一样,T细胞和B细胞都是来源于骨髓中的干细胞。特异性免疫的基础是抗原识别受体。在T细胞上有两种多链结构的抗原识别受体,即多数的α/β和少数的γ/δ异二聚体以及多链的信号转导元件。T细胞抗原识别受体或T细胞识别受体(TCR)是约含10~18个氨基酸的线性多肽。这些多肽曾经在抗原呈递细胞(APC)中被加工处理过,并被呈递至表达于APC上的主要组织相容性复合体(MHC)的抗原结合槽中的TCR上。B细胞抗原受体在结构上与IgM抗体相似,并且可识别三维结构完整的抗原。这就说明T和B细胞所识别的表位是不同的。由于淋巴细胞发育过程中受体成分的复杂基因重排(rearrangement)结果,每个B细胞和T细胞抗原受体所识别的抗原结构对这一细胞也是有其独特性的。在抗原刺激后,B细胞中有进一步的基因重排以增强抗体的亲和性,即所谓体细胞成熟(somatic maturation)。这样就形成了特异性免疫应答具备能与自然界无数抗原结构发生反应多样性的独特性。T细胞在其发育早期进入胸腺,在此发生正向和负向选择。负向选择包括消除能与自身抗原发生反应的潜在性T细胞,以降低自身免疫性发生的危险,而正向选择则可以选择30%已经生产性地重排了其TCR的T细胞,而70%的T细胞重排的结果只是形成无功能性的TCR。
2.主要组织相容性复合体 另有一些能使免疫应答特异性增强的机制,如抗原必须与主要组织相容性复合体(MHC)受体一起呈递至T细胞,以及需要有T细胞辅助B细胞形成抗体。有两种类型的MHC受体,即MHC-Ⅰ类受体和MHC-Ⅱ类受体(在人类称为HLA抗原)。MHC-Ⅰ类受体由3个功能区的单一链与共同表达的β2-微球蛋白所组成,并且广泛表达于所有的有核细胞上。MHC-Ⅱ类受体由一条α-链和一条β-链所组成,并且只是固有地表达于专职APC上,特别是树突状细胞和B细胞;炎症时则可在许多类型的细胞上表达。MHC-Ⅰ类和MHC-Ⅱ类受体在结构上相似,其呈递抗原的部分有一类似贝壳状的结构,在此抗原多肽位于TCR所在处两条脊之间。MHC受体均是高度多态性的,但是T细胞和B细胞多态性的程度并不相同,人类中有3种MHC-Ⅰ类基因,即HLA-A,HLA-B和HLA-C基因,以及至少6种MHC-Ⅱ类基因,其中的3种,即DR,DQ和DP在抗原呈递过程中尤为重要。每一个MHC基因都有许多不同的等位基因,这些等位基因原本可通过血清分型法鉴定,最近可用测序法检出,即某一个体可在核苷酸水平上由其MHC结构来表现其独特性。外周的T细胞与在胸腺中者不同,不能直接与游离的抗原发生反应,而必须是通过M HC受体适当提呈的抗原,这样就限制了T细胞与自身抗原发生反应的可能性。T细胞受体与M HC的相互作用极为特异,在抗原多肽、M HC受体或T细胞受体上只要有单一氨基酸的改变,即可有明显的反应的发生。某些多肽只能与某些M HC等位基因产物结合,这样就可以解释某些自身免疫病与M H C表型的相关性,如90%的关节强直性脊柱炎患者均有H L A B27等位基因,而只有10%以下的正常人群具有这种表型。一般认为这是因为M HC等位基因以高度的亲和性与某些交叉反应性肽类结合,从而导致不适当T细胞刺激的危险性增加。CD4结合M HC-Ⅱ类受体稳定彼此之间的相互作用;而CD8则与M H C-Ⅰ类受体结合。因此表达CD4的T细胞只能识别由M H C-Ⅱ类受体所呈递的抗原;而CD8+T细胞则只能识别由M H C-Ⅰ类受体呈递的抗原。一般而言,M HC-Ⅰ类受体呈递内源性细胞抗原,即细胞内寄生的细菌和病毒;M HC-Ⅱ类受体则呈递为APC所吞噬的外源性抗原肽,包括细胞外细菌、寄生虫和无感染性的抗原。由此可见,CD8+T细胞(常称细胞毒性T细胞)一般认为在抗病毒的宿主防御上很重要,因其能裂解表面表达M HC-Ⅰ类受体的病毒感染细胞。CD4+T细胞则与细菌和寄生虫感染的细胞介导免疫、非感染性抗原,如变应原的免疫应答以及协助B细胞产生抗体(其原始的名称即为T辅助细胞)有关。不过事实上这种区分并不是绝对的,因为CD4细胞显然也可对病毒抗原发生反应,而且对细胞内寄生的病原体,如结核分枝杆菌和肺孢子虫的宿主防御也很重要,并已证明CD8细胞能生成许多细胞因子,参与协助B细胞作用,并可对非病毒抗原发生反应。
3.抗原呈递 抗原必须与表达于APC上的自身M HC一起提呈至T细胞。APC具备摄取和处理抗原的生化机制,并能将处理后的抗原负载于M HC受体之上。此外,APC还必须表达能与T细胞表面的反受体(counter receptor)相结合的辅佐分子,并能提供T细胞活化与增殖时所必需的T细胞受体所能提供者的信号。一般认为,幼稚T细胞对于活化的要求极高,特别是在体外,需要有高浓度的抗原和APC上辅佐分子的高度表达。已证实有许多不同类型的细胞能在体外呈递抗原至记忆性T细胞,并可促进抗原特异性T细胞增殖。这些细胞包括巨噬细胞、B细胞、上皮细胞,甚至嗜酸性粒细胞。但是,树突状细胞呈递抗原的功效较之其他类型的细胞高100倍,其本身就可决定淋巴结中幼稚T细胞的活化,并且也是记忆性细胞的主要APC,B细胞在与T细胞相互作用以产生抗体时也可呈递抗原至T细胞。
树突状细胞可在体内除了脑部以外的所有的组织内发现,在肺部形成支气管上皮细胞内或其下的稀疏而广泛的网络。树突状细胞来源于骨髓中的前体细胞,有两种类型:一种类型来源于淋巴样前体细胞;另一类型来源于髓样前体细胞。淋巴样前体细胞在胸腺内发现,在此它们与自身反应性T细胞的负向缺失有很重要的关系。淋巴样树突状细胞也可在淋巴结中发现,但在肺部的主要类型为髓样型。其形态特别,有较长的细长突起由细胞体伸出,形成其特殊的类似蜘蛛的形状。
树突状细胞的发育过程中经过一系列的发育步骤,特别是在组织中有其不成熟时期,此时细胞能有效地通过吞饮作用、吞噬作用,或者通过抗体结合至Fc受体等方式摄取抗原,但不能由于低水平的M H C受体的表达而部分地呈递抗原。当细胞摄取了抗原以后,就可通过输入淋巴管转移至局部淋巴结。在肺部的情况下,就是肺门和气管旁淋巴结,并在此成熟成为极为有效的抗原呈递细胞。这种细胞上有很高水平的MHC-Ⅰ类和MHC-Ⅱ类受体的表达,还有能与T细胞上CD28相结合的辅佐分子CD80和CD86的表达。这时的细胞就不再能有效地摄取抗原了。此种与细胞功能要求相关的成熟和转移过程受到生长因子和趋化因子的控制,如GM-CSF能在体外使树突状细胞成熟;树突状细胞也可表达许多趋化因子受体。它们可协助细胞通过输入淋巴管的转移。
虽然幼稚细胞在淋巴结中致敏比较明确,但对记忆性细胞能在组织中被刺激的程度还是不大清楚的。炎症疾病时树突状细胞很快地转移至组织,而且细胞因子,如GM-CSF的释放构成炎症过程的一部分,并可促使组织树突状细胞在其到达淋巴结以前成熟,以便于以后提呈抗原至组织中居留的T细胞。对此过程的一种抗衡作用表现在肺泡和支气管巨噬细胞的下调作用上,这些细胞虽传统上被认为均是APC,但却可能对淋巴细胞活化具有重要的免疫抑制效应。也有许多研究证实AM在体外能抑制外周血中T细胞的抗原特异性增殖。这种效应部分上是由AM所释出的NO作用于树突状细胞所介导的,但确切的机制尚不清楚。在哮喘时,AM也被活化,且丧失其免疫抑制效应,使慢性T细胞刺激得以发生。
4.淋巴细胞活化 T淋巴细胞可区分为幼稚细胞、母细胞、效应细胞和记忆细胞。人类淋巴细胞的这些亚群可根据同工型CD45的受体表型来区分:CD45RO可作为记忆细胞的标记;CD45RA则是幼稚细胞的标记。表达于新近活化细胞上的活化标记为CD69,HLADR,CD25和VLA-1(very late activation antigen)。幼稚细胞不能识别可为其受体所识别的抗原。母细胞可以被抗原刺激,并可表现效应功能,如细胞毒作用、辅助B细胞和细胞因子产生。记忆细胞则可在其效应时期中识别抗原,并可再次成为静止的细胞。幼稚细胞对于抗原刺激较之记忆细胞有更严格的要求,需要有专职抗原呈递细胞(主要是树突状细胞)的呈递抗原,而且只有在次级淋巴样器官,如淋巴结、Peyers结和支气管相关淋巴样组织(BALT)内方可受到刺激。宿主与抗原首次接触所出现的原发应答需要有大量抗原的刺激,发生较慢,且较之由于刺激记忆细胞群体所形成的续发应答的数量、力度上为低。T细胞不论是在原发应答或是续发应答时,一旦接触了抗原就可立即增殖,分泌细胞因子和表现效应功能。如无持续的抗原刺激,这些细胞中的多数死亡,仅存留少量的长寿命的记忆细胞。血液中的多数淋巴细胞都是静止细胞,T细胞效应功能多在组织中表现。Sallusto等提出,外周血记忆细胞可根据淋巴结归巢受体CCR7的表达而进一步区分:CCR7阴性记忆细胞多进入组织,且在抗原刺激后立即产生细胞因子;而CCR7阳性细胞则进入淋巴结,增殖以补充记忆细胞群体。但只是有限地生成效应性细胞因子,如IFNγ。同样,B细胞也可以幼稚细胞和记忆细胞存在,在其效应时期则为产生抗体的母细胞或浆细胞,B细胞需要有来自T细胞的刺激以产生抗体,即所谓T细胞协助。B细胞主要存在于淋巴结内,邻近生发滤泡处。幼稚B细胞亦可摄取结合于其受体上的抗原,加以处理,然后呈递至记忆T细胞(它们事先曾经受到树突状细胞的刺激)。然后T细胞通过可溶性细胞因子及膜结合信号(特别是CD40:CD40配体)刺激B细胞。IL-4的产生与B细胞的转换至生成IgE有关;IL-6和TGFβ则可引起生成IgA的转换。IgG主要在淋巴结内产生;IgA以及IgE则主要由肠道固有层中的B细胞产生。
仅由抗原受体的T细胞刺激尚不足以促使T细胞增殖和表现其效应功能,尚必须另有次级信号存在。这种信号是由辅佐受体CD28与其表达于APC上的反受体CD80和CD86结合后产生的。此外,T细胞尚需要有APC产生的IL-1的刺激,从而以自分泌的方式引起细胞增殖。另外有一些辅佐分子,如LF A-1/ICA M-1和ICA M-3以及CD2/LF A-3都可提供附加的扩增信号。如果不能提供这些信号的全部程式,则T细胞就会变为无反应性而不能发生反应,或者由于凋亡而死亡。
(二)Th1/Th2型黏膜免疫
早在20世纪80年代就已证实,小鼠的CD4T细胞可以根据细胞因子产生的类型而区分为若干亚群,一种亚群细胞称为Th1细胞,可产生IFNγ和IL-2;另一亚群则称为Th2细胞,则可产生IL-4,IL-5,IL-6和IL-10。这两亚群细胞都可产生IL-3和G M-CSF。第三种亚群称为Tho细胞,能产生这些细胞因子的混合物。以后在人类的T细胞克隆中也发现有相似的情况,但是细胞因子程式并不相同。例如,IL-10的产生在人类中就不十分明显。以后又鉴定出一些具有Th1(IL-12)和Th2(IL-9和IL-13)型T细胞应答程式的细胞因子,也发现了CD8 Tc1和Tc2克隆。IFNγ的产生使Th1细胞与抗细菌性感染(如结核分枝杆菌感染)的典型细胞免疫以及自身免疫病有关;而IL-4,IL-5和IL-6的产生则使Th2细胞与抗体产生以及宿主对蠕虫病和过敏性疾病有关。广义而言,这种区分在动物模型和人类疾病中都已证实。因此,自身免疫性应当视作为Th1免疫的失调。而过敏性为Th2型免疫的异常。何者决定T细胞以Th1型应答或Th2型应答对抗原发性反应,是一个复杂的问题,而且目前对此的了解也不够。简言之,IL-4和IFN-γ(以及IL-12)分别抑制Th1和Th2细胞的产生,而且T细胞受到诱导的细胞因子环境至关重要。另外,抗原剂量(低剂量有利于Th1)、抗原性质(变应原为具有酶活性的无感染性的蛋白质)以及免疫途径(黏膜免疫较之全身性免疫更有利于Th2)也都与之有关。近年来在西方国家内特应症和过敏性疾病发生率的上升,促进了研究者对于何者决定Th1/Th2应答的问题进行研究。结果表明,虽然遗传因素起相当重要的作用,但在某一相同基因群体中短期内发病率的增加则可证明环境因素可能起着更为重要的作用,与变应原接触的增加也是部分的原因。目前有人认为,为了防止宿主的排斥,胎儿的环境有利于Th2型免疫应答对由母体循环中获得抗原的反应,这些抗原包括诸如HDM之类的变应原。胎儿宿主体内以后可以产生替代基础Th2免疫的Th1应答。在特应症个体中可能由基因程序化的结果,Th1应答发育成熟缓慢,结果是使Th2应答能得以牢固地形成。例如已证实在特应症婴儿中IFN-γ的产生有缺陷。这种理论与有关的理论认为在缺少与细菌和病毒性抗原接触的现代生活中的早期儿童,其Th1应答不明显的事实是一致的。但是这一理论却不能解释为什么Th2应答发生于接触了变应原或寄生虫抗原的免疫幼稚的成年人,一般认为,记忆性T细胞(特别是Th2细胞)对于以后的再次刺激多保留其Th1/Th2的等同性,提示这是次级淋巴样组织中的微环境,在此时产生最初的对Th1或Th2应答都很重要的诱导作用。树突状细胞对影响这一过程极为重要,而且近期事实表明,其所产生信号的性质对于T细胞发育的方向有深远的影响。虽然个别T细胞克隆在细胞培养中可保留其Th1/Th2等同性,但是也有可能在事先致敏个体的外周血单核细胞(PBMC)的抗原刺激时,经过用适当的细胞因子或抗细胞因子抗体(如用IL-12抑制Th2应答)处理后,可使之转向Th2型的炎症应答。至于这种现象究竟是由于Th2细胞等同性的改变,或者是由于生成新的记忆性细胞群体,还是少部分Th1记忆细胞的新生,都不十分明确。由Th1向Th2细胞转变的措施已被用作为治疗哮喘的基础,如用IFN-γ治疗哮喘,但目前的疗效不大,因为有刺激产生有组织损伤作用的Th1应答的危险性。
(三)肺部作为共同免疫系统的一部分
通过肠道、呼吸道和泌尿道黏膜表面接受抗原刺激的黏膜免疫系统有别于全身免疫应答。在全身免疫应答时,抗原刺激是通过血流而进入各个内脏,如肝脏、胰腺和骨髓。皮肤一般认为是全身免疫系统的一部分,不过它更接近于黏膜表面,有一种直接与外界环境接触的特殊化的上皮细胞。肺脏有丰富的血液供应,它与肾脏一样可视为既有全身免疫又有黏膜免疫组分的器官。在正常条件下,多数抗原通过黏膜表面进入体内。这些抗原既可是活的生物,如细菌和病毒,也可是无生命的抗原,如食物抗原和吸入的蛋白质,肠道和呼吸道黏膜有广泛表面面积存在,说明宿主必须处理大量的抗原物质。其结果是黏膜免疫系统必须发育成特殊化的淋巴样组织网络,用以调节免疫应答。在此,必须区分免疫应答的诱导或致敏时期和效应时期。前者包括抗原被提呈至次级淋巴样组织中的幼稚T细胞;后者则表现经抗原刺激T细胞的各项功能。肠道次级淋巴样组织为Peyers结(PP)和肠系膜淋巴结。PP中有一种特殊的上皮细胞,称为M细胞,它们能直接在肠腔中摄取抗原,但PP并无输入淋巴管。呼吸道与之相应的组织是鼻相关淋巴样组织(NALT)和支气管相关淋巴样组织(BALT)。BALT由支气管上皮细胞以下的聚集的淋巴细胞所组成,它和PP一样也无输入淋巴管和高内皮细胞小静脉(HEV)。根据物种和年龄的不同,BALT的存在也有所变化,如在家兔和豚鼠小鼠的某些品系中较为明显,在正常成人中不存在,但由20岁以下的人体中取出的肺脏中约有40%具有这一结构。人类中的NALT以Waldemeyers环淋巴样组织为代表,这种淋巴样组织包括腺样增殖体、腭淋巴样组织和扁桃体,而扁桃体的结构更接近于传统的淋巴结。
公共黏膜免疫系统概念的关键性特征是,T细胞和B细胞在此系统的某一方面被致敏或受到诱导,如在肠道致敏或诱导,则可策动系统中的其他方面,如肺部表现其效应功能。这种情况表明,T细胞和B细胞有一共同的归巢机制,用以移动至黏膜组织。这一理论的事实基础来自动物模型的研究,某一器官的致敏可导致其他器官的黏膜免疫系统中具有抗原特异性细胞。例如,用HIV gp160蛋白鼻内免疫小鼠后,能产生血清、肺部和阴道中的HIV-1中和抗体。这项研究所存在的问题是,经鼻免疫黏膜的抗原也有可能先进入血流,引起全身性免疫应答,然后再进入其他部位。对此问题的解决需要有更多对有关器官特异性淋巴细胞移动过程分子基础的了解。而现有有关事实提示,引导T细胞和B细胞至肠道和呼吸道的信号之间是有差异的。例如有大量的B细胞寄住于肠道的固有膜(体内80%的Ig产生细胞位于结肠黏膜),但与T细胞相比较,呼吸道中的B细胞相对较少。MAdCAM-1是肠道归巢T细胞的一种着在分子或着在素(addressin)就不表达于呼吸道。在β7基因缺失的小鼠中,肠道淋巴细胞有明显的减少,但对肺部淋巴细胞的数目上的影响极少。
有关肺部和肠道免疫性之间具有共同性的研究为数不多。共同黏膜免疫系统的概念可用以解释为什么某一器官中的基于免疫的炎症疾病,如溃疡性结肠炎可以与另一器官中的疾病,如支气管扩张有关。但是这种肺部与肠道疾病的联系并不多见,而是与其他非黏膜器官疾病的联系较为多见,如哮喘与特应性皮炎,肺纤维化与关节炎之间的联系。
(四)黏膜耐受性
免疫系统必须能区别对宿主有害的抗原刺激(如致病性微生物)和对宿主无害的惰性物质。一般而言,黏膜免疫系统对惰性物质有耐受性,而对致病性微生物则有活性。许多免疫介导疾病,如哮喘、外源性过敏性肺泡炎和结肠炎的发生是由于免疫系统的致耐功能缺陷;而传染病则是由于活性应答的缺陷所致。已有愈来愈多的事实表明,为感染性微生物所刺激的天然免疫组分可对特异性免疫系统发出信号,使其向活性应答方向发展。在这方面,耐受性就是一种有缺陷的状态。此外除了有能刺激活化性免疫的抗原存在以外,尚需有其他信号的事实,就可用以解释当进行实验使动物对多种抗原致敏时需要使用佐剂的原因。耐受性是一个极为复杂的现象,对其的了解仅是开始。较多的研究集中于经口耐受性,这一现象系指用适当的方式将抗原经口注入抗原,经过暂时性的免疫应答后,即对抗原刺激不发生反应而表现为耐受性。可用低剂量或极高的剂量的抗原喂动物使之致耐。低剂量致耐是由于抑制性细胞的诱导,而高剂量致耐则是由于T细胞无反应性(抗原特异性T细胞对抗原不起反应)和T细胞缺失(抗原特异性细胞因凋亡而消失)两者的联合作用。如将抗原静脉注入,即可通过T细胞的无反应性和缺失,而不是抑制细胞应答的诱导机制而诱发耐受性。而且高剂量耐受性尚可使抗原在血流中出现,因而成为全身性刺激物。已知有两种抑制性T细胞与耐受性有关,一种亚群为CD4阳性,并且产生与耐药性诱导有关的主要细胞因子TGFβ。这种细胞称为Th3细胞,它们除能诱发其所针对抗原的耐受性以外,还可诱发针对旁立无关抗原的耐受性,包括用免疫优势肽致敏后针对整个抗原的耐受性。Th3应答的作用与Th1应答的作用相反,而且可以相互抑制。Th2应答一般较之Th1应答对组织的损害作用较低,但却能扩大Th3应答的作用。不过其作用力度不够,而且也并不是十分需要的。控制Th3应答发生的途径尚不完全了解。结肠炎动物模型中的研究显示Th3细胞的生成和TGFβ的产生有缺陷,另外有一机制认为位于上皮细胞间的γ-δCD8 T细胞与之有关。但是在CD8缺失的动物中耐受性正常,而在CD4缺陷的小鼠中不能形成耐受性。故CD4机制可能是更重要。此外,人类肠道中的γδ-CD8细胞并不像小鼠中者那样明显。另外尚鉴定出一CD4+/CD25+T细胞亚群也与肠道中的耐受性有关。这一亚群的缺失可导致小鼠中的结肠炎。虽然这些细胞并不分泌IL-10和TGFβ,但其免疫抑制效应却需要有些细胞因子的存在。免疫抑制性CD4+/CD25+细胞也可在人类的外周血中发现,但其在肺部免疫中的作用尚不清楚。
近年来对于通过呼吸道致敏(常用鼻黏膜致敏)而诱导耐受性已有较多的研究,通过呼吸道致敏可涉及由Th1向Th2应答转变。早期研究中发现,用卵白蛋白鼻内免疫小鼠后可引起起始的Ig E应答,然后转变至Th1型的抗体产生程式,并可受到γ-δT细胞、小鼠CD4细胞和大鼠CD8细胞的调节。最近的研究则用与自身免疫病有关的抗原或多肽高剂量鼻内致敏。结果也发现有明显的由Th1向Th2应答的转变,也是受到CD4或γ-δCD8 T细胞的控制。另有一高剂量对尘螨抗原所致的耐受性模型中发现,抗原的释放引起暂时性的CD4应答,后继之以无应答性或无反应性。此种情况可能与调节性CD4细胞有关,而有别于前述经口耐受性中产生TGFβ的CD4 Th3细胞。
上述的多数研究主要是在小鼠中进行,而且在人类中的应用并不肯定。为了防止慢性炎症状态,必然要对每天都可由呼吸道吸入的多数惰性抗原的某些种类产生耐受性。例如,尘螨抗原在非特应性个体中并不引起肺部疾病,但H D M特异性Th1 T细胞却可在外周血中检出,提示上述的耐受性并不起作用。A M的抑制作用也很重要,因为有一事实表明,A M在哮喘时无抑制作用。在αVβ6基因缺失小鼠中。可因中和了TGFβ效应而发生慢性肺部炎症,这一效应同时可以防止抗肿瘤药物博来霉素(Bleomycin)诱发的纤维化。可见肺部TGFβ的固有表达能表现负向免疫调控作用,而过量产生则可引起纤维化。但是这种效应与上述的Th3诱导的耐受性是否有关尚待证实。另有一种由IL-10所表现的免疫耐受性,即在用高剂量的黄蜂和蜜蜂毒素对过敏性个体进行脱过敏免疫治疗时,可以引起由一抗原特异性T细胞产生的IL-10释放。IL-10通过阻抑CD28而干扰抗原呈递细胞的共同刺激作用,从而抑制Th1和Th2细胞的活化。产生IL-10的T细胞亦可用其他变应原的高剂量免疫治疗方式而诱生,小鼠中针对呼吸道变应原的耐受性则是由于成熟树突状细胞产生IL-10后再又刺激产生IL-10调节性T细胞的结果。
总之,耐受性应答在肠道和呼吸道黏膜免疫应答中都很重要,并且受到许多复杂机制的调节,包括抗原特异性T细胞的无反应性或缺失、抑制性T细胞的活化以及天然免疫系统细胞对组织中效应T细胞应答的直接效应作用。耐受性应答可视作与组织损伤效应免疫应答有关的Th1应答的反映。而Th2应答则多被认为是非组织损伤性应答。有嗜酸性粒细胞和其IL-5和eatoxin表达的正常肠道则是一种Th2型的环境,与过敏反应性呼吸道的Th2环境相似。特应症的一项特点就是多数的特应症个体不会患有哮喘,即令此种个体曾在其呼吸道中有嗜酸性粒细胞数目增加的Th2介导的免疫应答表现亦属如此。
(五)肺部T细胞
正常肺脏除了是抗原进入的一种重要途径以外,也是富含淋巴样组织的器官。与其他种类的白细胞相比较。正常成年人呼吸道黏膜下组织和肺泡壁中有大量的T细胞。对许多哮喘病人的和对照者的免疫病理学研究结果表明,正常个体呼吸道中的T细胞数目较之中性粒细胞要多10倍,哮喘病人的T细胞较之呼吸道中无炎症者多2倍。但是支气管肺泡抽取液和痰液的分析结果就不十分可靠,这是因为淋巴细胞与粒细胞有所不同,它们往往不能通过上皮细胞屏障而停留于上皮细胞以内或位于其下。此外,原本少量存在于支气管管腔中的T细胞尚有返回组织中的可能性。在一些品系的小鼠中,BALT中的淋巴细胞聚集可见于呼吸道的分支处,但在成年人呼吸道中则无此结构。不过也可在气管杈隆凸下形成淋巴细胞聚团的趋势。虽然BALT的确切功能尚不清楚,但它也含有形态上与在淋巴结中所见者相同的HEV,提示BALT可能与将抗原呈递至幼稚T细胞的过程有关。在肺部95%的淋巴细胞为T细胞。只有少数的B细胞。这些T细胞几乎全部表达αβ-T细胞受体,这是与γδ受体是有所不同的,95%以上为CD45ROHICH记忆细胞。上皮细胞间T细胞在呼吸道中较为常见。而且其数目可在肺部炎症疾病时增加。不过这些研究一般难以进行,因为由于组织中捕捉和加工的结果,上皮细胞极易脱落,故无可靠的系统研究。同样上皮细胞间CD4和CD8细胞的比例也不清楚,但在BAL和分散的肺部细胞中总的比率如同在血液中者一样,都是2∶1。肺部T细胞对识别抗原的T细胞受体特异性程式表现多样性,与血液中所表现者并无显著的差异,提示肺部T细胞亦可识别大量的蛋白结构,尘螨过敏个体BALT细胞的研究发现有1%以下的HAM反应性T细胞,可以预期针对呼吸道病毒感染肺部T细胞的数目必然较大。病毒和细菌可使应用Ⅰ类四聚体的抗原特异性CD8长寿命效应细胞移向组织,其中也包括肺部。接触了细菌性抗原以后也可观察到类似的CD4+细胞移动。此外。小鼠中的Sendai病毒亦可引起CD4+特异细胞的生成,这些细胞能在肺部生存达6个月之久,再次刺激后可迅速产生细胞因子,可对Sendai病毒的再次感染表现保护性免疫力。
1.肺部T细胞的表型 肺部T细胞的受体表达程式与血液中T细胞有所不同。多数肺部的CD4和CD8 T细胞为CD69和V L A-1阳性细胞,CD69是体外刺激T细胞的早期活化标记;V L A-1则是晚期活化标记,也是胶原蛋白和层素(laminin)的受体。至于CD25(另一早期活化标记)的表达则较弱,或不表达。血液和肺部T细胞两者之间的另一显著不同为αEβ7的表达。αEβ7表达于50%的CD4细胞和80%的CD8细胞上,这显然是与血液中主要表达于CD8细胞上的情况是不相同的。αEβ6是E-钙黏着蛋白(E-cadaherin)受体。这种受体是否在上皮细胞间T细胞上表达尚未肯定,但很有可能无其表达,因为只有一小部分的肺部T细胞是上皮细胞间性质者。这种黏附受体表达程式的功能和意义尚不清楚。外周血中很少有细胞表达CD69,V L A-1或αEβ7,提示这些受体是因诱导而进入肺部者。在有TGFβ存在时的T细胞刺激可诱导αEβ7的表达。当T细胞通过肺部,或当大部肺部T细胞停留于肺部相当长的时间之际,CD69,V L A-1和αEβ7的表达很快地受到诱导,然后并受到下调。CD69,V L A-1和αEβ7的表达提示这是一种活化表型,但并不意味在正常肺部就有活化的效应T细胞存在。转基因小鼠中发现,所有的T细胞均可识别卵白蛋白,并在受到刺激以前的肺部T细胞即已增殖良好,且可分泌IL-2;但在吸入卵白蛋白以后,T细胞却停止增殖,并且开始产生细胞因子。如其为Th2细胞则产生IL-4,IL-5,而如其为Th1细胞则产生IF N-γ。人肺中的T细胞除非是受到刺激是不能产生细胞因子的。很有可能存在两种类型的记忆细胞。一种类型是在适当刺激下产生细胞因子,而且是位于组织中的效应细胞;另一类型则是通过增殖以再生记忆性细胞群体的记忆细胞。如此情况属实,则多数寄居肺部T细胞应为效应细胞。肺部T细胞的表型如表3-3所示。
表3-3 肺部T细胞的表型

2.T淋巴细胞的肺部移动 淋巴细胞由血流再循环至淋巴样组织,然后返回血流。幼稚T细胞只有通过特殊的高内皮细胞小静脉(HEV)方可进入次级淋巴样组织。记忆细胞则是通过毛细血管后内皮细胞穿过淋巴结,其中有的细胞再通过输入淋巴管返回血流。有一部分的组织记忆细胞,可能是已经活化者则仍停留于局部组织。幼稚细胞的循环并无任何特殊器官或某组淋巴结的偏向性,但是记忆细胞则多是回归至其首次接触抗原的器官和有关淋巴结,如此将增加抗原特异性细胞识别抗原的机会。这种移动程式的调节受控于T细胞上的黏附(归巢)受体及其内皮细胞上的配体(着点素)以及趋化因子受体和趋化因子。目前,研究者已对有关次级淋巴结、肠道和皮肤的器官特异性归巢机制进行了大量的工作。结果显示,L-选择素和趋化因子6-C-kine(SLC)与CCR7的结合可以引导T细胞归巢至外周淋巴结;α4β7与MAdCAM-1的结合则可引导致肠道的归巢;作为E-选择素以及趋化因子TARC配体的皮肤淋巴细胞抗原(CLA)与CCR4的结合可引导致皮肤的归巢。目前尚未发现特异性肺部淋巴细胞归巢受体,但在肺部的确有内皮细胞黏附受体的存在。肺部T细胞也的确有明显的趋化因子和黏附受体表型,提示在肺部也有选择性的移动程式存在(表3-4)。
表3-4 肺部内皮细胞黏附受体的表达
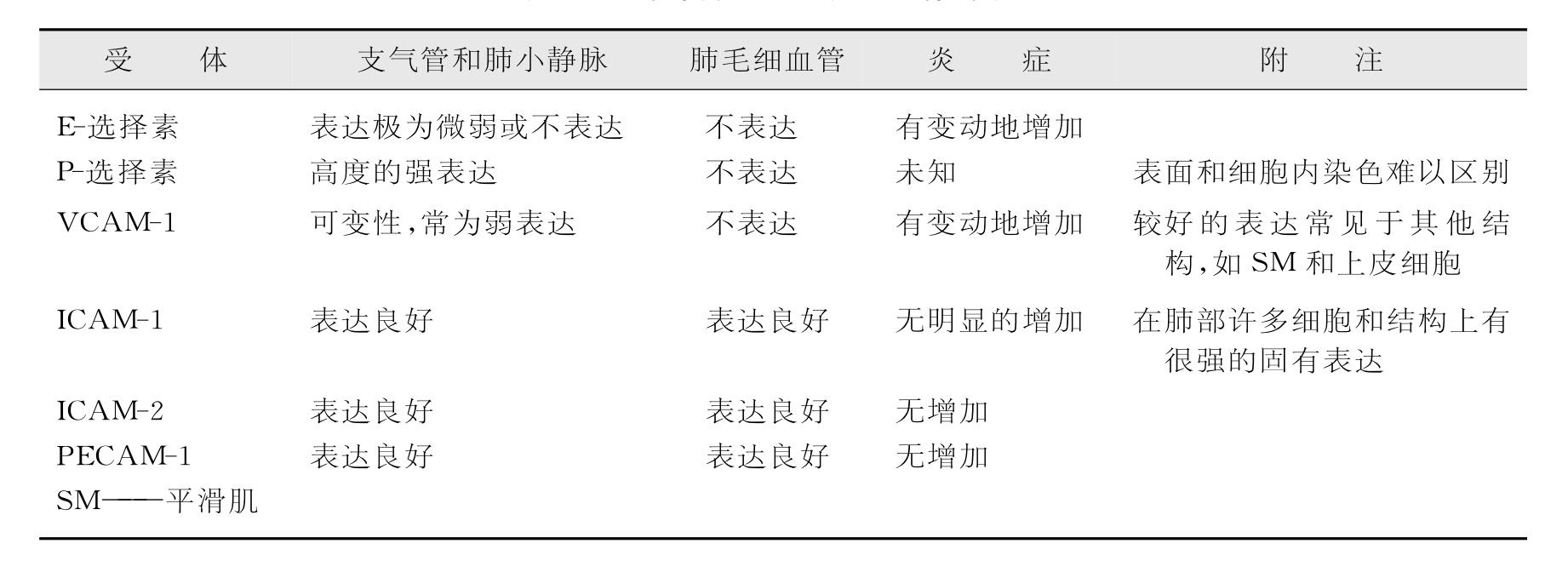
此外,特别是在绵羊中的研究已证实了肠道和呼吸道中再循环的方式。在大鼠的鼻相关淋巴样组织(NALT)中的T细胞结合为L-选择素,而不是MAdCAM-1依赖性的,提示其与外周淋巴结的联系较之肠道淋巴结更为密切。在绵羊中的情况则不同,肠道相关T细胞是α4β7高表达,L-选择素低表达;外周淋巴结T细胞是α4β7低表达,L-选择素高表达;肺部淋巴液中的T细胞则是这两种受体的表达均低,提示是绵羊中的不同移动方式。
一般应用小鼠作为动物模型进行有关淋巴细胞归巢至肠道和周围淋巴结的研究结果,对于肺部并不十分适用。小鼠的肺脏与人肺在解剖上极不相同,特别是在呼吸道的结构上,如其结构中的固有层、软骨和平滑肌就较少。另外,上皮细胞多为单层细胞,而不像人肺中所见者为层状上皮细胞。正常小鼠的肺间质或固有层中的淋巴细胞为数较少,而且在BAL中发现者也很少,肺部淋巴细胞归巢行为特征为区室化,即每种淋巴细胞都具有不同的移动行为方式以及其可能的功能。这些区室包括肺泡壁和肺泡间的间质性淋巴细胞、支气管固有层淋巴细胞、上皮细胞间淋巴细胞、BAL中所收集呼吸道管腔中的淋巴细胞和肺部脉管内的淋巴细胞。
过敏性炎症动物模型研究结果表明,T细胞的移动部分依赖VLA-4,VCAM-1和P-选择素。同样,用绵羊红细胞气管内攻击小鼠中向肺部移动的T细胞则富含有E-选择素和P-选择素配体,而且小鼠肺中E-选择素和P-选择素的表达如同VCA M-1的表达一样也有所增加。以后的研究发现抗V L A-4抗体能抑制移动过程,而且在Fuc T VⅡ缺陷小鼠中的移动也可能因不能表达选择素配体而被抑制。对正常肺黏附受体表达的研究比较困难,而且多数的黏附受体表达研究采用肺癌切除标本的浅层支气管内膜活检,或者采用肺部炎症疾病活检。正常肺中P-选择素在呼吸道和肺静脉内皮细胞上的表达良好,但不表达于毛细血管内皮细胞。炎症疾病时P-选择素表达上调的范围难以确定,因为在正常个体中也有很强的染色性,而且细胞内和管腔内染色性的差异不大,难以区分。毛细血管的缺少染色性是与毛细血管的剪切应力(shear stress)较低有关,因而不需要有选择素来捕捉白细胞。E-选择素在非炎症肺部的表达微弱,而在炎症肺部呼吸道内皮细胞上的表达往往受到上调。ICA M-1,ICA M-2和PECA M则广泛表达于无炎症状态的所有类型内皮细胞。ICA M-1尚可在肺内的其他结构上表达,包括支气管上皮细胞。VCA M-1多在呼吸道和肺静脉内皮细胞上微弱表达或表达不定,而在肺毛细血管内皮细胞上则不表达。但是VCA M-1的表达在平滑肌和固有膜中的其他细胞以及某些个体的呼吸道上皮细胞中则很明显。其表达在炎症中的上调范围不十分肯定。M AdCA M-1在正常情况下不表达于肺部内皮细胞。
(六)肺部B细胞
共同黏膜免疫系统的概念原本来自肠道、呼吸道和生殖泌尿系统表面分泌物中Ig A浓度较之在血液中增加的特征,人类中的Ig A多由肠道固有膜中的B细胞和浆细胞产生。黏液分泌物中的Ig A主要以二聚体形式存在,即两个免疫球蛋白分子由一称为J链的多肽连接,并由组织中免疫细胞分泌。这一单位然后结合至表达于上皮细胞基底表面上,称为分泌片的受体上。此种复合体内在化后即被转运至其分泌细胞的近管腔表面。与肠道中的情况不同,呼吸道固有层中的B细胞很少。因此呼吸道分泌物(如痰液和BA L)中,较之肠道只有少量的Ig A。IgG亦可在支气管管腔内发现,但可能并不是在局部生成的。在特应性疾病中,局部组织B细胞尚可产生可在呼吸道分泌物中发现的Ig E。B细胞和生成Ig A细胞的缺乏是与Ig A缺陷个体中,一般很少有肺部症状的事实相一致的,除非是也有IgG亚类的缺陷。这与一般可引起支气管扩张的IgG缺陷有所不同。
三、结论
肺是致病性和非致病性微生物侵入体内的主要门户,因而必须具备包括屏障机制以及天然和获得性免疫系统在内的有效免疫系统与之抗衡。肺脏是特异性免疫应答的重要器官,与相关次级淋巴样结构一道,构成诱导性致敏作用的主要场所。不适当的免疫应答则可导致不良的后果;如在低免疫应答时导致感染;高免疫应答时发生肺部炎症疾病。肺部宿主防御机制的进一步了解至关重要,以期使这些疾病能得到有效的治疗。
(余传霖)
参考文献
1.Wardlaw AJ.Immunological basis of lung disease.In:Wardlaw AJ,Hamid Q,eds.Textbook of respirato-
ry cell and molecular biology.London:Martin Dunitz Ltd.,2002,47~71
2.Hoffmann JA,Kafatos FC,Janeway JCA,et al.Phylogenetic perspectives in innate immunity.Science,1999,284:1 313~1 317
3.Bals R,Wang X,Wu Z,et al.Human beta-definsin-2is a salt-sensitive peptide antibiotic expressed in human lung.J Clin Invest,1998,102:874~880
4.Liu L,Wang L,Jia HP,et al.Structure and mapping of the human beta-definsin HBD-2gene and its expression at sites of inflammation.Gene,1998,222 237~244
5.Harder J,Bartels J,Christophers E,et al.Isolation and characterization of human beta-definsin-3,a novel human inducible peptide antibiotic.J Biol Chem,2001,276:5 707~5 713
6.Goldman MJ,Andersen CM,Stolzenberg ED,et al.Human beta definsinⅠis a salt sensitive antibiotic in lung that is inactivated in cystic fibrosis.Cell,1997,88:553~560
7.Elrod KC,Moore WR,Abraham WM,et al.Lactoferrin,apotent tryptase inhibitor,abolishes late-phase airway responses in allergic sheep.Am J Respr Crit Care Med,1997,156:375~381
8.Hoppe HL,Reid KBM.Collections——soluble proteins containing collagenous regions and lectin domains——and their roles in innate immunity.Protein Sci,1994,3:1 143~1 158
9.Wright JR,Borron P,Brinker KG,et al.Surfactant protein A:regulation of innate and adaptive immune responses in lung inflammation.Am J Respir Cell Mol Biol,2001,224:512~517
10.Wardlaw AJ.Molecular basis for selective eosinophil trafficking in asthma:a multi-step paradigm.J Allergy Clin Immunol,1999,104:917~926
11.Bradding P,Holgate ST.Immunopathology and human mast cell cytokines.Curr Rev Oncol Haematol,1999,31:119~133
12.Holt PG,Oliver J,Bily KN,et al.Down-regulation of the antigen presenting cell function(s)of pulmonary dendritic cells in vivo by resident alveolar macrophages.J Exp Med,1993,177:397~407
13.Shibata Y,Berclaz PY,Chroneos ZC,et al.GM-CSF regulates alveolar macrophage differentiation and innate immunity in the lung through PU,1.Immunity,2001,15:557~567
14.Luster AD.Chemokines——chemotactic cytokines that mediate inflammation.New Engl J Med,1998,338:436~445
15.Choller-Martin S,Montravers P,Gibert C,et al.High levels of interleukin-8in the blood and alveolar spaces of patients with pneumonia and adult respiratory distress syndrome.Infect Immunol,1993,61:4 553~4 559
16.Sauty A,Dziejman M,Taha RA,et al.The T cell specific CXC chemokines IP-10,Mig and I-TAC are expressed by activated human bronchial epithelial cells.J Immunol,1999,162:3 549~3 558
17.Zlotnik A,Yoshie O.Chemokines:a new classification system and their role in immunity.Immunity,2001,12:121~127
18.Springer TA.Traffic signals for lymphocyte recirculation and leukocyte emigration,the multistep paradigm.Cell,1994,76:301~314
19.McEver RP,Moore KL,Cummings RD.Leukocyte trafficking mediated by selectin-carbohydrate interactions.J Biol Chem,1995,117:11 025~11 028
20.Kansas GS.Selections and their ligands:current concepts and controversies.Blood,1998,88:3 259~3 287
21.Yang J,Furie BC,Furie B.The biology of P-selectin glycoprotein ligand 1:its role as a selectin counter-receptor in leukocyte-endothelial and leukocyte——platelet interaction.Thromb Haemost,1999,81:1~7
22.Shimizu Y,Rose DM,Ginsberg M H.Integrins in the immune system.Adv Immunol,1999,72:325~380
23.Doerschuk CM.Adhesion molecules and cellular biochemical changes in acute lung injury.Chest,1999,116:S37~43
24.Hogg JC,Doerschuk CM.Leukocyte traffic in the lung.Ann Rev Physiol,1995,57:97~114
25.Doerschuk CM.Neutrophil rheology and transit through capillaries and sinosoids.Am J Respr Crit Care Med,1999,59:1 693~1 695
26.Mizgerd JP,Horwitz BH,Quillen HC,et al.Effects of CD18 deficiency on the emigration of murine neutrophils during pneumonia.J Immunol,1999,163:995~999
27.Li XC,Miyasaka M,Issekutz TB.Blood monocyte migration to acute lung inflammation involves both CD11.CD18 and very late activation antigen-4-dependent and independent pathways.J Immunol,1998,161:5 258~6 264
28.Sallusto F,Kremmer E,Palermo B,et al.Switch in chemokine receptor expression upon TCR stimulation reveals novel homing potential for recently activated T cells.Eur J Immunol,1999,29:2 037~2 045
29.Lambrecht BN,Prins JB,Hoogsteden HC.Lung dendritic cells and host immunity to infection.Eur Respri J,2001,18:692~7 044
30.Vermaelen K Y,Carro-Muino I,Lambrecht BN,et al.Specific migratory dendritic cells rapidly transport antigen from the airways to the thoracic lymph nodes.J Exp Med,2001,193:51~60
31.Robinson DS,Hamid Q,Ying S,et al.Predominant Th2-like bronchoalveolar T lymphocyte population in atopic asthma.N Engl J Med,1992,326:298~304
32.Kay AB,Ying S,Varney V,et al.Messenger RNA expression of the cytokine gene cluster,interleukin-3(IL-3),IL-4,IL-5 and granulocyte/macrophage colony-stimulating factor,in allergen-induced late phase cutaneous reactions in atopic subjects.J Exp Med,1991,173:775~778
33.Constant SL,Bottomly K.Induction of Th1 and Th2 CD4+T cell responses:the alternative approaches.Ann Rev Immunol,1997,15:297~322
34.Tang M L K,Kemp AS,Thorburn J,et al.Reduced interferon-γsecretion in neonates and subsequent atopy.Lancet,1994,344:983~986
35.Strachan DP.Allergy and family size:a riddle worth solving.Clin Exp Allergy,1997,27:235~236
36.Rissoan MC,Soumeelis V,Kadowaki N,et al.Reciprocal control of T helper cell and dendritic cell differentiation.Science,1999,283:1 183~1 186
37.Tschernig T,Kleemann WJ,Pabst R.Bronchus-associated lymphoid tissue(BA L T)in the jungs of children who had died from sudden infant death syndrome and other causes.Thorax,1995,50:658~660
38.Hiller AS,Tschernig T,Kleemann WJ,et al.Bronchus-associated lymphoid tissue(BA L T)and larynx associated lymphoid tissue(L A L T)are found at different frequencies in children,adolescents and adults.Scand J Immunol,1998,47:159~162
39.Csencsits K L,Jutila M A,Pascual DW.Nasal-associated lymphoid tissue phenotypic and functional evidence for the primary role of peripheral node addressin in naive lymphocyte adhesion to endothelial venules in a mucosal site.J Immunol,1999,63:1 382~1 389
40.van Cott TC,Kaminski RW,Mascola JR,et al.HIV-1 neutralizing antibodies in the genital and respiratory tracts of mice intranasally immunized with oligomeric gp160.J Immunol,1998,160:2 000~2 012
41.O’Garra.A cytokines induce the development of functionally heterogeneous T helper cell subsets.Immunity,1998,8:275~283
42.Maloy KJ,Powrie F.Regulatory T cells in the control of immune pathology.Nat Immunol,2001,2:816~822
43.Singh B,Read S,Asseman C,et al.Control of intestinal inflammation by regulatory T cells.Immunol Rev,2001,182:190~200
44.Lowrey JL,Savage ND,Palliser D,et al.Induction of tolerance via the respiratory mucosa.Int Arch Allergy Immunol,1998,116:93~102
45.McMenamin C,Pimm C,McKersey M,et al.Regulation of IgE responses to inhaled antigen in mice by antigen-specific gamma-delta T cells.Science,1994,265:1 859~1 871
46.Tian J,Atkinson MA,Clare-Salzler,et al.Nasal administration of glutamate decarboxylase(GAD65)peptides induces Th2responses and prevents murine insulin dependent diabetes.J Exp Med,1886,183:1 561~1 567
47.Harrison LC,Dempsey-Collier M,Kramer DR,et al.Aerosol insulin induces regulatory CD8gamma-delta T cells that prevent murine insulin dependent diabetes.J Exp Med,1996,184:2 167~2 174
48.Hoyne G,O’Hehir RE,Wraith DC,et al.Inhibition on T cell and antibody responses to house dust mite allergen by inhalation of the dominant T cell epitope in naive and sensitized mice.J Exp Med,1993,178:1 783~1 788
49.Huang XZ,Wu JF,Cass D,et al.Inactivation of the integrin beta 6subunit gene reveals a role of epithelial integrins in regulating inflammation in the lung and skin.J Cell Biol,1996,133:921~928
50.Munger JS,Huang XZ,Kawahatsu H,et al.The integrin alpha v beta 5binds and activates latent TGF beta 1:a mechanism for regulating pulmonary inflammation and fibrosis.Cell,1999,96:319~328
51.Akdis CA,Blaser K.Mechanisms of interleukin-10mediated immune suppression.Immunology,2001,103:121~136
52.Akbari O,de Kruyff RH,Umetsu DT.Pulmonary dendritic cells producing IL-10mediate tolerance induced by respiratory exposure to antigen.Nat Immunol,2001,2:725~731
53.Kay AB.Pathology of mild,severe and fatal asthma.Am J Respr Crit Care Med,1996,154:S66~S69
54.Pabst R,Binns RM.Lymphocytes migrate from the bronchoalveolar space to regional bronchial lymph nodes.Am J Respr Crit Care Med,1995,151:495~499
55.Masopust D,Vezys V,Marzo AL,et al.Preferential localization of effector memory cells in nonlymphoid tissue.Science,2001,291:2 413~2 417
56.Reinhardt RL,Khoruts A,Merica A,et al.Visualizing the generation of memory CD4Tcells in the whole body.Nature,2001,410:101~105
57.Woodland DL,Hogan RJ,Zhong W.Cellular immunity and memory to respiratory virus infections.Immunol Res,2001,24:53~67
58.Hogan RJ,Zhong W,Usherwood EI,et al.Protection from respiratory virus infection can be mediated by antigen-specific CD4+T cells that persist in the lungs.J Exp Med,2001 193:981~986
59.Kilshaw Pj,Murant SI.Integrins on mouse lymphocytes relevance ti the mucosal immune system.Eur J Immunol,1991,21:2 591~2 597
60.Lee SC,Jaffar ZH,Wan KS,et al.Regulation of pulmonary T cell responses to inhaled antigen:role in Th1and Th2mediated inflammation.J Immunol,1999,162:6 867~6 879
61.Butcher EC,Williams M,Youngman K,et al.Lymphocyte trafficking and regional immunity.Adv Immunol,1999,72:209~253
62.Campbell JJ,Bowman EP,Murphy K,et al.6-C-kine(SLC),a lymphocyte adhesion triggering chemokine expressed by high endothelium,is an agonist for the MIP-3 beta receptor CCR7.J Cell Biol,1998,141:1 053~1 059
63.Camobell JJ,Haraldsen G,Pan J,et al.The Chemokine receptor CCR4 in vascular recognition by cutaneous but not intestinal memory T cells.Nature,1999,400:776~780
64.Campbell JJ,Brightling CE,Symon FA,et al.Expression of chemokine receptors by lung T cells from normal and asthmatic subjects.J Immunol,2001,166:2 842~2 848
65.D’Anbrosio D,Mariani M,Panina-Bordignon P,et al.Chemokines and their receptors guiding T lymphocyte recirculation in lung inflammation.Am J Respr Crit Care Med,2001,184:1 266~1 275
66.Abitorabi M A,Mackay CR,Jerome E H,et al.Differential expression of homing molecules on recirculating lymphocytes from sheep gut,peripheral and lung lymph.J Immunol,1996,156:3 111~3 117
67.Pabst R.Compartmentalization and kinetics of lymphoid cells in the lung.Reg Immunol,1990,3:62~71
68.Krug N,Tschering T,Holgate S,et al.How do lymphocytes get into the asthmatic airways?Lymphocyte traffic into and within the lung in asthma.Clin Exp Allergy,1998,29:8~10
69.Wolber FM,Curtis JL,Milak A M,et al.Lymphocyte recruitment and the kinetics of adhesion receptor expression during the pulmonary immune response to particulate antigen.Am J Pathol,1997,151:1 715~1 727
70.Wolber FM,Curtis JL,Maly P,et al.Endothelial selections and the alpha 4 integrins regulate independent pathways of T lymphocyte recruitment in the pulmonary immune response.J Immunol,1998,198:4 396~4 403
71.Southcott A M,Hemingway I,Lorimer S,et al.Adhesion molecule expression in the lung:a comparison between normal and diffuse interstitial lung disease.Eur Respr J,1998,11:91~98
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。









