



钝叶黄檀异地保存林种群结构特征
孙永玉12,陈艳彬3,罗长维1,李昆1,2
(1.中国林业科学研究院资源昆虫研究所,昆明650224;2.国家林业局元谋荒漠生态定位站,元谋650224)
摘要:为了解钝叶黄檀种质资源异地保存林的种群动态过程及其适应机理;采用标准地调查及种群大小结构分析法,对钝叶黄檀异地保存林种群结构特征、生长情况进行调查分析。结果表明该种群处于稳定增长的状态,根据生长表现来进行初步筛选,镇沅种源、双江种源的种质材料在生长表现上优于云县种源和龙陵种源。在株高、地径结构分布上,4个种源的分布趋势相一致;钝叶黄檀种质资源异地保存林能够切实有效的保存所收集的种质资源,可为以后的育种工作提供丰富的遗传材料。
关键词:钝叶黄檀;种质资源保存林;种群生态特征
钝叶黄檀(Dalbergia obtusifolia Prain)又名牛肋巴、牛筋木,为蝶形花科(Papilionaceae)黄檀属(Dalbergia)的一种旱季半落叶乔木树种,是紫胶虫(Kerria cheninsesMaldh.)最佳生长地干热河谷二半山区阳性耐旱代表性树种。钝叶黄檀是我国生产应用最广泛的紫胶虫寄主植物,冬代保种效果较好,其树体化学成分和有机成分适于紫胶吸食和生长[1-2],我国50%的紫胶原胶、80%的种胶均利用钝叶黄檀生产,在我国紫胶生产中起着重要的作用,国内外目前关于该植物及相关属种的相关研究限于化学成分、有机成分、生理、种苗、生态学特征与病虫害等方面[1-6]。近年来,随着钝叶黄檀分布区滥砍滥伐和过度放牧等人为影响,导致该树种在我国的自然分布不断缩小,许多种质材料大量丧失,资源保护面临着前所未有的严重局面[4,7]。基于此,中国林业科学研究院资源昆虫所在云南景东建设了钝叶黄檀种质资源异地保存林。同一植物不同种群间由于长期生存在不同环境中,选择压力的不同,必然会造成种群内基因和基因频率或大或小的差异,但每个种群的基因在一定时期内是相对稳定的[8-9]。目前,植物异地保存林内不同种群特征的研究尚无报道,在同一异地保存林中,所保存的不同种源材料因为其遗传因素的差异,其种群特征也应有所不同。研究钝叶黄檀种质资源异地保存林内不同种源保存材料间的种群特征差异,可为进一步从分子生物学机制上研究其基因差异奠定基础,并能深入了解钝叶黄檀种质资源异地保存林的动态过程及其适应机理,为该种群的保护及其他植物的保存提供理论基础与科学依据。
1 材料与方法
1.1 研究地点
钝叶黄檀异地保存林在中国林科院资源昆虫所景东试验站枇杷山,面积为1.4万m2。该地位于横断山脉南端,经纬度E100.82°、N24.42°,海拔1260m,属亚热带季风型气候,年平均气温18.3℃,日平均气温≥10℃的活动积温为6440.9℃,年均日照2131.6小时,无霜期355天,年平对湿度77%,年均降雨量1086.7mm,降水丰沛但干湿季分明,5~10月为雨季,降雨量占全年的87.14%,11月至次年4月为雨季,降雨极其稀少,降雨量仅占全年的12.86%,植被盖度为56%(旱季)。枇杷山地势平缓,坡度不大,土壤类型为赤红壤。
1.2 研究时间
不同种源钝叶黄檀种子于2005年5月上中旬采自钝叶黄檀主要分布区,分别为澜沧江干热河谷流域的龙陵、金沙江干热河谷流域的云县和双江、红河干热河谷流域的镇沅4个县市区。2005年6月年进行钝叶黄檀的营养袋育苗工作,并于2006年6月雨季初进行钝叶黄檀种质资源异地保存林建设工作,造林株行距为2m×3m,造林和管理水平一致。调查时间为2008年11月。
1.3 研究方法
1.3.1 样地选择和样地调查
根据4个地理种源钝叶黄檀的种植地点,选择生长发育良好、结构整齐、层次分明、坡度相对比较平缓的地段设置了4个标准地样方(见表1)。由于钝叶黄檀种质资源异地保存林人为干扰比较频繁,每年都有除草等人工管理行为,钝叶黄檀异地保存林中除有历年来种植的印楝(Azadirachta indica)和板栗(Castanea millissima)的孤立木以外,没有自然萌发的其他种类林木,林下主要草类品种为禾本科(Poaceae)的短颖马唐(Digitaria microbachne)、大画眉草(Eragrostis cilianensis),莎草科(Cyperaceae)的香附子(Cyperus rotundus),菊科(Asteraceae)的清明草(Anaphalis nepalensis)、三叶鬼针草(Bidens pilosa)、紫茎泽兰(Eupatorium adenophorum),蝶形花科(Fabaceae)的假地豆(Desmedium heterocarpum)等。
表1 调查样方基本情况

1.3.2 各样方生长情况调查
对各样方的钝叶黄檀进行每木调查,分别记录各自的株高、地径、冠幅长、冠幅宽和存活率等指标。
1.3.3 保存林立木大小结构划分
由于年龄结构相同,种群的大小及结构的划分用地径和树高分,各为四个立木级别。地径的划分标准:Ⅰ.0<X<2cm,Ⅱ.2cm≤X<3cm,Ⅲ.3cm≤X<4cm,Ⅳ:>4cm。株高的划分标准为Ⅰ.0cm<X<100cm,Ⅱ.100≤X<150cm,Ⅲ.150≤X<200cm,Ⅳ.>200cm。在此基础上对钝叶黄檀种质资源异地保存林各调查样方调查指标进行分析。采用SPSS与EXCEL统计软件进行数据分析与作图。
2 结果与分析
2.1 不同种源钝叶黄檀人工种群存活情况调查
钝叶黄檀种质资源异地保存林作为一个栽培面积较大的人工林群落,由于管理得当,不同种质材料保存率都比较高(见表2),龙陵种源保存率高达96.42%,其次为双江种源的92.81%和云县种源89.25%,镇沅种源的保存率在4个种源中最低,为77.78%。经过2年的野外生长观察,不同种源的钝叶黄檀均可适应旱季恶劣的水分条件并能正常生长。
表2 不同种源种质材料保存率(T)

2.2 钝叶黄檀不同种源间生长指标比较
表3 不同种源种质材料生长情况
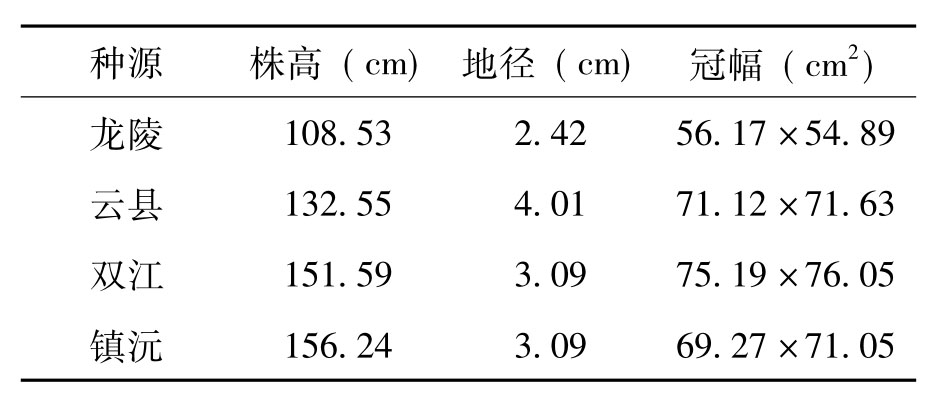
表4 不同种源间地径和株高间的LSD分析

注:不同种源同一指标间,具有相同字母表示差异不显著,不同字母表示差异显著,**上标表示差异极其显著。
镇沅种源和双江种源平均高生长和平均地径生长在4个种源中相对较好(见表3),平均株高分别为156.24cm和151.59cm,地径3.09cm。云县种源钝叶黄檀在4个种源中地径生长量最高,平均达到4.00cm,株高生长也达到132.55cm。龙陵种源在4个家系中生长情况较差,平均株高为108.53 cm,地径2.42 cm。双江种源在4个种源中冠幅最大,平均冠幅为75.19×76.05 cm2,其后依次为云县种源、镇沅种源和龙陵种源。从野外生长调查情况上分析,龙陵种源、双江种源生长较好,其次为云县种源,龙陵种源的生长在4个种源中最差。方差分析表明(见表4)不同种源间株高、地径和冠幅之间的差异也比较显著。
2.3 不同种源钝叶黄檀基于地径的立木级结构分布
图1 不同种源钝叶黄檀人工保存林径级结构分布图
用径级结构代替年龄结构研究钝叶黄檀种群,对4个样地的统计数据进行综合,其结果如图1所示,不同种源间地径分布差别稍有差异。在调查样方中,龙陵种源279株个体中Ⅰ级、Ⅱ级别的植株占调查总数的72.69%,云县种源的Ⅰ级、Ⅱ级别植株也占该种源的60.90%,说明龙陵种源和云县主干细弱的植株比较多,Ⅰ级、Ⅱ级别的个体多。双江种源和镇沅种源Ⅲ级、Ⅳ级别个体较多,分别占各自调查数的47.90%和52.4%,说明这2个种源总体生长良好。径级结构分布调查表明,在钝叶黄檀保存林中,不同种源间的地径生长差别明显。
2.3 不同种源钝叶黄檀基于株高的立木级结构分布

图2 不同种源钝叶黄檀人工保存林株高立木级结构分布图
株高立木级结构分布图也表明不同种源间立木级株高分布差别较大(见图2)。龙陵种源Ⅰ、Ⅱ低矮株高级别的树木个体较多,占调查总数的82.15%,说明该种源株高总体偏矮。云县种源Ⅰ级、Ⅱ级别株高占总调查数的61.84%,Ⅲ级和Ⅳ级别的占总调查数的38.16%,种源内较高的树木相对也较少。双江种源Ⅰ级别株高占总调查数的12.68%,Ⅲ和Ⅳ级别的占51.05%,该种源高大的树木总体上较多。镇沅种源高大的个体较多,Ⅲ和Ⅳ级别的个体占调查总数的52.38,株高在0~100cm的矮小植株数量也比较少。各种源地径和株高结构分布图也验证了4份种质材料的野外具体生长情况,在野外生长中,镇沅种源、双江种源的树木较其他两个种源普遍高大,种源内生长表现优良的个体比较多;云县种源次之;龙陵种源内低矮植株多,生长普遍较矮。
3 讨论
研究乔木种群结构时,一般从遗传结构、空间结构、年龄结构和大小结构4个方面来分析。钝叶黄檀种群的遗传结构已经进行了RAPD分析,相关研究尚待发表。在钝叶黄檀异地保存林中,不同种源种质材料都种植在景东枇杷山,虽然各种源具体种植地点的坡向和坡度稍有区别,但土壤、水分、气候、降雨等空间结构特征都相同,因此空间结构不能用来反应不同种源钝叶黄檀的种群生态特征。种群的大小和年龄结构是种群内不同大小、年龄个体数量的分布状况,不仅反映种群不同大小和年龄个体的组配情况,也反映了种群数量动态及其发展趋势,并在很大程度上反映种群与环境间的相互关系及在其在群落中的作用和地位[10]。对于钝叶黄檀异地保存林,种群的年龄较比较一致,育苗及栽培时间都为3年,栽培密度和管理水平也一致,用不同种源钝叶黄檀的大小结构来分析各自的种群动态是比较适宜的方法,许多学者通常采用大小结构代替年龄结构的方法[10-13]分析植物种群结构特征,本文采用株高和地径来代替传统的年龄结构分析钝叶黄檀不同种源的种群生态特征,结果与具体野外的生长情况比较吻合。
某一物种对特定生境发生了基因型反应而产生相应的基因,是种内适应不同环境条件或地理区域的结果,也是环境因子对一个种群内许多基因型选择和控制的结果[12]。不同种源钝叶黄檀保存率、生长情况和立木结构的不同,这是景东当地自然环境因子选择和不同种源钝叶黄檀自身遗传特性相结合的结果。钝叶黄檀种质资源异地保存林由于管理得当,4个种源的保存率都比较高,龙陵种源、双江种源、云县种源和镇沅种源的保存率分别为96.42%、92.81%、89.25%和77.78%。根据生长表现来进行初步筛选,镇沅种源、双江种源的种质材料在生长表现上优于龙陵种源和双江种源,具体的家系筛选和个体筛选仍需从生长表现、紫胶虫吸食成分分析及放胶效果等多方面进行综合观察和验证。
影响钝叶黄檀种质资源异地保存林基因库容量大小的因素主要来源于种植时不同种群、家系、个体的数量以及选择方法。许多发展紫胶产业的胶民一般会从树姿较好、枝条平展、产胶产量高且易于收放胶的钝叶黄檀母树上采集种子进行繁殖,这样繁殖的个体遗传基础大部分相同,在后代的繁殖上会造成近交,后代的遗传素质会严重下降,特别是只从一棵树上采集的种子育苗建立的种群,植物的一半遗传物质来自于同一亲本,另外一半的来源也非常有限,造成了遗传分离负荷较大。由于近交有时候不影响结实率或影响很小,再用来育苗和建立新的种群,随着近交的循环,基因库将逐渐减少,植物的遗传素质也会降低,种质质量就会下降。因此,在钝叶黄檀异地保存林种群的建立过程中,采集了不同分布流域的多个种源,并集中种植在同一地段,力图扩大所保存的钝叶黄檀基因库容量,经过2年多的野外生长观察,各种源种质材料保存率较高,生长良好,个体间变异较大,该保存林能切实有效的保存所收集的钝叶黄檀种质资源,并能为以后的遗传育种工作提供丰富的选择材料。
[1]Bogdanova V T.Some differences in chemical composition of lac host plants[J].Rastitelnve Resoursvl.1965,1(3):380-383.
[2]M.S.Haque.Free amino acids in the anal fluids of female lac insect feeding on different host plants[J].Indian J.1978,46(3):291-298.
[3]孙永玉,李昆,杨文云,等.钝叶黄檀种子与幼苗的地理种源变异[J].林业科学研究,2005,18(3): 296-299.
[4]孙永玉,李昆,陈晓鸣.中华紫胶虫优良寄主植物钝叶黄檀研究[J].林业科技开发,2005,19(5):7-9.
[5]陈玉德,侯开卫.紫胶虫优良寄主——钝叶黄檀的生态生物学特性[J].广西植物,1980,(1):12-15.
[6]中国科学院综合考察委员会紫胶考察队编.紫胶虫的寄主植物[M].北京:农业出版社,1972:4-7.
[7]孙永玉,李昆.紫胶虫寄主植物研究现状与展望[J].西南林学院学报.2002,22(3):75-80.
[8]刘智慧.四川省缙云山栲树种群结构和动态的初步研究[J].植物生态学与地植物学学报,1990,14(2):120-128.
[9]Pacala,S.W.,Silander,J A J.Tests of neighbourhood population dynamicmodels in field communities of two annualweed species[J].Ecologicalmonographs.1990,60(2):113~134.
[10]朱学雷,安树青,张立新,等.海南五指山热带山地雨林主要种群结构特征分析[J].应用生态学报,1999,10(6):641-644.
[11]吴承祯.洪伟.吴继林,等.珍稀濒危植物长苞铁杉的分布格局[J].植物资源与环境学报,2000,9(1):31-34.
[12]周纪伦.植物种群生态学[M].北京:高等教育出版社,1992.
[13]Jonathan Silvertown,Deborah Chalesworth.Introduction to Plant Population Biology[M].Oxford:Blanckwell publishing,1988.
Study on community structure inDalbergia obtusifolia Prain ex situ conservation forest
Sun Yong-yu1,2,Chen Yanbin3,Luo Changwei1,Likun1,2
(1.Research Institute ofResource Insects,CAF Kunming,650224;2.SFA's Yuanmou degraded ecosystem location station;3.Panzhihua state forest farms,Panzhihua,617007)
Abstract:[Objective]The research aimed to study the population structure characteristics and adaptation mechanism of Dalbergia obtusifolia Prain ex situ conservation forest.[Methods]Used sample tree-plot field investigation and population size structuremethods,investigated the forest population structure characteristics and growth conditions.[Results]The conservation rate of different geographical provenance were all high for the appropriatemanagement,and the populationswere increasing steadily,it can effectively conserve Dalbergia obtusifolia Prain germ.The primary evaluation result based on growth shows that Zhenyuan provenance was the best in four geographical provenances,and then Shuangjiang,Yunxian and Longling provenance.Stumpage distributing fig based on tree heigh and collar diameterwere similar among four provenances.[Conclusion]The ex situ conservation forest can effectively conserve germplasm resources and offer genetic stock for breeding.
Key words:Dalbergia obtusifolia Prain;germ conservation forest;community structure
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











