



第一节 玉米育种技术快速发展
一、20世纪80年代以前我国玉米育种状况
大约在16世纪初期玉米被引入我国,至今已有500年左右的种植历史,并已成为我国主要栽培作物,形成了从东北到华北平原至云贵高原的斜向玉米带。但我国近代玉米育种研究起步较晚,在新中国成立以前,各地区种植的基本上为地方品种。纵观我国玉米育种的历史,可将其分为两个时期,即1926~1949年的创建时期和1949年以后的杂种优势利用时期。
1925年赵连芳在中央大学农学院率先开始玉米自交系和杂交种选育工作。1926年王绶、郝钦铭、翁德齐、孙仲逸在南京金陵大学农学院亦开展玉米自交系的分离和杂交种的组配工作,虽以教学为目的,但开创了我国近代玉米育种的先河。随后的24年里我国玉米育种开始了艰难的创建过程。1930年南京中央大学农学院赵连芳、金善宝与丁振麟开始玉米育种工作,至1934年已选育玉米自交系500余份,并用40余份自交系配制杂交组合。同年金善宝发表了我国第一部玉米育种专著《近代玉米育种法》,首次系统地介绍了美国近代(1914~1934年)玉米杂交育种理论和技术成果,成为我国近代玉米育种的重要参考书。1931年山西省铭贤学校在太谷进行引种美国玉米试验,从中选出优良品种“金皇后”,先后被推广到华北、东北、西南和东南各省,成为20世纪50~60年代中期栽培面积最大的玉米品种之一。1936~1943年范福仁在广西农事试验场进行了较系统的玉米自交系选育和杂交组配,先后育成一批有利用价值的自交系和双交种,并适应地区试种。1943~1945年张连桂、李先闻在四川省农业改进所先后育成404、411、452、458等四个双交种。1938~1940年中央农业实验所将吴绍暌、蒋德麒从美国带回的64份玉米杂交种分别在成都、贵阳、昆明、柳州等地进行比较试验,从中选出有利用价值的Wisc.696、Cornell 29-3等杂交种。1941年戴松恩在《农报》发表文章《抗建期中玉米杂交种之推广问题》,指出美国双交种在我国种植,适应性较差,增产效果不显著;而应利用地方种质选育自交系,再与外来自交系组配双交种,这一观点至今对我国玉米杂交育种仍具有重要指导意义。同年王绶等与西北区推广繁殖站合作,通过改良玉米地方品种选育自交系,育成了武功白玉米和武功综交白玉米。1947~1948年吴绍暌、郑廷标在南京金陵大学进行硬粒型与马齿型品种间杂交组合比较试验,获得较好的增产效果,比双亲增产22%~24%。1947~1949年陈启文等在山东莒县引进并推广“金皇后”,并与小粒红组配成品种间杂交种“坊杂二号”。1949年以前,因处于抗日战争和解放战争时期,我国玉米育种工作缺乏系统性和连贯性。虽然在地方品种改良和自交系、杂交种选育上取得了一些成果,但在生产上发挥的作用十分有限。
1949~1959年,随着新中国成立,对粮食的需求增大。1949年12月吴绍暌在全国农业生产会议上提出杂种优势利用策略和措施,即一方面选育和利用品种间杂交种,另一方面着手开展自交系间杂交种选育工作。从此玉米杂种优势开始在玉米育种和生产上应用。1950年国家提出“去雄选种、培育品种间杂交种、综合种、双交种”的全国玉米改良和杂交种推广方案。通过群众性的玉米良种收集、鉴定和评选,短期内完成了一次品种更换,促进了玉米生产发展。据统计,1950年全国共收集地方品种14 000份,评选出良种2 000个,在生产上推广200多个,种植面积较大的有43个,其中主要有金皇后、金顶子、白马牙、英粒子、辽东白等。随后,全国一些主要农业科学研究所和农业院校相继开展了品种间杂交种和自交系选育工作。由于没有品种审定工作,品种一旦表现优良,即在生产上应用,因此一大批品种间杂交种迅速得以推广。如山东的坊杂2号、齐育25,东北的大金顶×黄马牙,河北的白头霜×大红袍,以及春杂1号、春杂2号、白杂6号、长杂4号、小粒红×黄马牙、大籽黄×金黄后等。
1960~1965年,在品种间杂交种推广应用的同时,玉米育种工作者着手进行自交系和双交种选育工作。1960年前后,国内育成了第一批双交种。由于双交种的产量明显高于品种间杂交种,因此迅速在生产上推广。如山东省农业科学研究所育成的双跃3号、双跃4号,北京农业大学育成的农大3号、农大4号、农大7号,四川农学院育成的川农7号、川农15号,河南新乡地区农科所育成的新双1号、新双2号、吉林农科院育成的吉双83、黑龙江农科院育成的黑玉号等。但双交种在生产上种植面积不足100万公顷,仅占全国玉米面积的4%。
1963年河南省新乡农科所育成了我国第一个玉米单交种——新单1号;1965年开始大面积种植,标志着我国玉米生产已由种植双交种时代转向单交种时代。到1980年初期,我国玉米生产经历了3次单交种的更换,即20世纪60年代中期的新单1号、白单4号等,70年代后期至80年代初期的丹玉6号等。单交种的种植面积逐渐扩大,由70年代中期的55%上升到70年代末的67.7%(1978年统计)。80年代以后,单交种在我国玉米主产区逐步普及,至1997年,已占全国玉米总面积的86.3%,品种类型亦发生了很大变化。
二、玉米育种种质基础渐趋拓宽
育种材料是玉米育种的物质基础,种质基础广泛是选育优良品种的保证。玉米原产于中南美洲,因此我国的玉米种质资源几乎都来自国外。由于我国地域辽阔,自然条件复杂,经过长期的自然选择和人工选择利用,以及天然授粉渐渗杂交造成的基因重组等原因,逐渐演化形成了各种不同生态适应型的地方品种,例如北方春玉米区的火苞米、白苞米、小短红、金顶子、白头霜,黄淮海夏玉米区的野鸡红、小粒红,东南玉米区的满堂金、小金黄,西南玉米区的大子黄等。这些地方品种基本上为硬粒型,适合在复杂生态条件下种植。在我国亦有极少数通过基因突变产生的新类型玉米(如糯玉米)。曾孟潜(1987)认为中国云南西双版纳等热带、亚热带地区是糯玉米的起源中心。在杂交种推广以前,我国玉米生产上应用的主要是引入的“金皇后”、“英粒子”和“白马牙”等马齿型自由授粉品种,并演变形成我国玉米区广泛种植的各种马齿类型地方品种。在当时生产力较低的年代,这些地方品种发挥了重要作用。由于我国长期处于闭关自守状态,玉米外来种质交流较少,利用效率也较低。
随着我国玉米育种研究的逐步开展,自由授粉品种在玉米生产中的地位相应降低。育种家们首先利用地方品种进行品种间杂交种或综合种的选育;与此同时也从地方品种中选育出一批优良自交系,如矮金525、混517、塘四平头、瑞北一、埃及205、吉63等。利用这批自育自交系与从美国引入的C103、M14、Oh43等高配合力抗病自交系,组配出我国第一代玉米单交种,如新单1号,白单1号、2号、4号,丰收101、103、105和吉单101等。从1971年全国两杂育种座谈会上,我国开始确立玉米杂交种利用原则,即“以单交种为主,双交种、三交种、顶交种及综合种,因地制宜、合理搭配种植”。这一原则加速了我国玉米自交系和单交种的选育进程。到20世纪70年代中期选育出自330、获白、二南24、甸11、黄早四、旅28等自交系,并进一步从美国引入高配合力自交系Mo17、B73,培育出我国第二代单交种,如豫农704、中单2号、丹玉6号、郑单2号、京早7号等。实践证明,自330、黄早四和Mo17在我国玉米育种和生产上发挥了巨大作用。
从20世纪80年代我国玉米杂交育种的历史可以看出,玉米育种的原始材料主要来源于以下几个方面:①从农家品种中选育一环系,如旅28、获白、甸11、塘四平头等。②从单交种中选育二环系,如自330、二南24、吉63等。从二环系的来源和系谱可见,大部分与国外种质资源关系密切。有些二环系直接选自引入的杂交种,如武105来源于美国杂交种Wis341;有些为国内系与国外系杂交组合的选系,如吉63选自吉双1号;有些是从含有国外系的综合种中选出的,如混517等。③从美国玉米带引入的自交系,如Mo17、B73、C103、M14、Oh43等。这些自交系大多来源于瑞德、兰卡斯特和克鲁格种质类群。
根据吴景锋(1983)的统计,1978年全国种植6.67万公顷以上的32个单交种仅由50个自交系组成,其中超过33.33万公顷的杂交种亲本主要有自330、塘四平头、获白、旅28、矮金525、M14、C104。到1980年种植6.67万公顷以上的29个杂交种由36个自交系组成,其中超过33.33万公顷的杂交种亲本主要为自330、塘四平头、获白、旅28、矮金525、M14、Mo17和大黄46。这种现象既说明了优良种质的重要性,同时也显示我国自交系的使用较为集中,育种种质基础比较狭窄。
20世纪80年代以来我国玉米育种又经历了两次大的品种更换。主要单交种包括丹玉13、吉单101、京早7号、烟单14(黄417、户单1号)、郑单2号、豫农704、恩单2号、勋单1号(七三单交)、东农248等,其中中单2号仍是大面积种植的品种。90年代推广的单交种主要包括掖单13、掖单2、四单19、豫玉18、西玉13、本育9、吉单159、成单14、沈单7、农大60、龙单13等,其中中单2号、丹玉13仍是大面积种植的品种;90年代末期至21世纪初期推广的品种主要有农大108、豫玉22、鲁单50、农大3138等。
从1980年种植6.67万公顷以上的玉米杂交种的遗传组成来看,亲本自交系的总数由1978年的50个下降到20世纪80年代末期的40个左右,其中国内一环系和引入系由1978年的22个和17个分别减至1987年的9个和5个,国内二环系由11个增加到27个。同时少数几个骨干自交系占有突出的地位,其中包括自330、获白、Mo17、黄早四。根据不完全统计,在已定名的杂交种中,由四个系组配的杂交种分别为46、22、42、和46个。可见,四大系在当时我国玉米生产中具有举足轻重的作用。与70年代的玉米育种种质相比,亲本自交系的使用更加趋于集中。但从杂交种的亲本组成来看,亲本自交系的种质来源发生了显著变化。第一,直接从外引或国内杂交种中选育二环系。由于这种方法易对亲本的某些性状进行改良,且会较快地获得稳定的自交系,用于组配新组合,因而得到广泛应用。根据曾三省(1990)统计,在3轮全国玉米区域试验的246个自交系中二环系占42.3%,如选自美国杂交种的中黄64、中黄32、沈5003、U8112、掖107等,南斯拉夫杂交种的南21、南23-32等,以及罗马尼亚杂交种的罗早4、罗31等。从国内杂交种中选出的自交系包括武109、武117、齐205、冬黄、大风7-1、铁75-55、原齐123、原齐150、京7、苏80-1等。第二,来源于群体或综合种的自交系比例明显上升,占利用总数的13.4%。鉴于20世纪70年代我国玉米种质资源贫乏状况,1978年李竞雄先生倡导开展群体改良工作。全国主要育种单位十分重视这项工作,根据生态区特点和自身力量调整了科研方向,采用不同方法合成了几十个广基或窄基群体,并进行群体选系工作,如综31、D729、辽轮814、东46等。同时开始着手从引入的墨西哥群体或热带和温带复合种质群体中选系,如木15、皖系11等。第三,来源于地方品种的选系应用面积缩小,但仍占总数的12.6%。地方品种的生态适应性很强,选育的自交系及组配的杂交组合抗逆性亦强,如耐贫瘠、耐阴湿、耐旱、耐低温等,因此地方品种选系仍是不可忽视的途径。如雄2-4、墩白、金09、交51等在玉米生产和区试中都占有一定的比重。第四,利用回交转育法获得的改良自交系已占有一定的比例。总之,80年代选系基础材料的来源已初步得到拓宽。
进入20世纪90年代,玉米种质扩增、改良与创新工作得到重视,已经从外来群体或半外来群体中陆续培育出一批新的优良自交系和杂交种,进一步拓宽了育种种质的遗传基础。利用热带、亚热带种质导入我国温带种质育成的新系开始运用于生产。截止1993年,我国玉米生产上基本形成了以丹玉13、中单2号、烟单14、掖单2号、掖单4号、掖单12和掖单13七个品种为主,以及Mo17、黄早四、自330、E28、掖478和丹340六个骨干系为主体的格局。单交种的更换主要由于掖单号杂交种的崛起及掖478和丹340的应用。对比1992年与1983年选系的基础材料来源可见,利用杂交种作选系基础材料的比率进一步加强,由1983年的37.9%上升到1992年的62.6%;地方品种选系进一步减少,由1983年的20.8%下降到3.2%,这说明单交种或三交种已成为玉米自交系的主要来源。但从选系的取材角度来看,育种素材已得到进一步拓宽。在这一阶段,育种素材不仅涉及到原有的种质资源,而且利用了美国玉米带的一批高配合力抗病杂交种(如78599等),同时还包含了热带、亚热带种质的利用(如Tuxpeno,ETO,Suwanl、也门品种等)。
从1996年,我国立项开展玉米种质扩增、改良和创新研究计划。核心内容是重点引进国外优质、抗逆、高配合力的核心种质和高效育种技术,在农艺性状和遗传评价的基础上,与国内现有种质利用相结合,确定我国实用的国内外核心种质的玉米杂种优势群,构建杂种优势利用模式,丰富我国玉米种质的遗传基础。截止2001年,形成了全国范围内的接力改良协作网,初步改良了19个热带、亚热带群体在我国玉米生态区的适应性,其中5个群体已适应了东北春玉米区的生态条件;合成了360份半外来群体。
野生种质资源的研究利用也取得了一定进展。四川农业大学(荣廷昭等,1997)通过栽培玉米与类玉米杂交获得了新的不育材料。中国科学院遗传所(郭乐群等,2000)将二倍体多年生大刍草基因转移到栽培玉米中,培育出异源自交系540等。先后利用540与沈5003配制出杂交种遗单6号、与选1302配制出高淀粉杂交种科玉10号。用二倍体多年生大刍草与一年生大刍草杂交选育出高产优质青贮饲料玉米品种8493。
三、玉米育种技术发展迅速
(一)群体改良与外来种质研究
自1919年美国Hays等把轮回选择方法用于品种改良后,玉米群体改良工作得到了较大发展。群体改良是指通过对玉米开放授粉品种或人工合成群体进行轮回选择(Recurrent selec-tion),使产量或其他性状获得周期性改进,从而创造改良的玉米育种材料。轮回选择方案可以打破连锁遗传,重组或积累有利基因,扩大遗传变异性,丰富种质基础,克服遗传脆弱性。改良群体既可以在生产上直接利用,亦可作为选系的基础材料。
美国和国际玉米小麦改良中心(CIMMYT)在玉米群体改良上做了大量卓有成效的工作。美国依阿华州立大学(Iowa State University)从1939年开始对坚秆综合种BSSS群体进行改良,至今已经历十几轮回选择。从C0、C5和C7轮次中分别选得B14、B37、B73、B84等一批优良自交系。目前,在美国玉米育种和生产中使用的自交系大多含有BSSS血缘。据Hallauer(1988)报道,1980年美国玉米杂交种亲本主要为11个骨干自交系,其中8个具有BSSS血统。除BSSS外,美国玉米育种家还合成并改良了一系列较有影响的群体,如密尼苏达13、西北马齿型、Lea米ing、抗螟虫群体BS1(H)C1、兰卡斯特复合种BSL(S)C6、早熟群体BS20、改良ETO群体BS16。同时分别于1984年和1986年引入哥伦比亚晚熟群体Tuxpeno和泰国群体Suwanl,经驯化改良后,目前已作为美国玉米带的育种材料。CIMMYT从世界各地收集和整理玉米种质资源,在农艺性状评价的基础上,合成了一系列综合种、群体和基因库;并采用改良半同胞法、S1或S2法、相互轮回选择法对合成种质进行了改良。目前,CIMMYT拥有各类玉米群体和基因库100多个,培育自交系450余份。
我国玉米群体改良工作起步较晚,20世纪70年代末才逐步得到重视和加强。经过20多年的努力,已培育出一大批优良群体并进行了初步改良,如中综2号、中综3号、中综4号、中群13、中群14、黄综、品综4号;豫综2号、豫综5号;陕综2号、陕综5号;忻综5号、忻综6号;辽旅群体、辽综群体;吉综A、吉综B、吉综D、吉综F、吉林库I等;WBM、WLS、LBM、LLS;东农群体1、东农群体2等。目前,从这批群体中已选育出一批优良自交系(如金黄96B、辽轮753、辽轮814、吉921、吉1037、吉1925、吉1928、豫25、武126、中自02、CA375、东8-1-1-2等,并组配出不少优良杂交种,如由金黄96B组配的协单969和由CA375组配的中单9409正在生产上推广。在我国,采用的玉米群体改良方法主要有混合选择(如中综3号、豫综5号、中群13、中群14等)、半同胞轮回选择法、全同胞轮回选择法、S1选择法、改良S1选择法、S2选择法、半姊妹轮回选择法等。
中国农科院用改良S1法、改良S1~HS联合鉴定法、改良S1和HS交替选择法对中综4号进行了3轮改良,三种方法每轮的产量增益分别为8.0%、7.2%和6.2%,配合力分别提高3.1%、5.1%和4.5%。吉林农科院利用改良穗行法对吉林库-I进行了5轮半同胞轮回选择,平均每轮的产量增益为6.4%,株高和穗位降低,抗倒性和抗病性显著提高,生育期提前34天。群体改良研究进展为我国玉米育种的持续发展奠定了长远基础。
长期以来,玉米育种工作者对新品种的选育,总是趋于选择适应性好、配合力高、综合性状优良的自交系作为杂交亲体,由此少数优良种质的利用频率过高,导致我国玉米育种的种质基础日益狭窄。这种现象增加了玉米杂交种遭受病虫侵害的危险性,品种间相近的血缘关系使其基因利用潜力受到限制,由此成为产量提高、改善品质、抗性增强的主要障碍。自20世纪80年代中期以来,我国玉米总产和单产徘徊不前,杂交育种处于爬坡状态,其根本原因仍是种质遗传基础狭窄,在品种产量和抗逆性改良与创新方面没有大的突破。虽然我国已进行了一些群体改良工作,但由于自身种质资源的局限性,品种改良效果不十分明显。从玉米育种的实践分析,我国玉米育种实质上是引种-驯化-选择利用-创造变异-再选择利用的过程。因此利用外来种质是拓宽我国现有玉米种质基础的重要途径。迄今为止,美国玉米带的各类材料以其适应性广、配合力高等优点成为我国利用较多的外来种质。然而来自玉米多样性中心的热带、亚热带种质具有我国乃至美国玉米种质所不具有的遗传变异,例如抗病性强(郭海鳌等,1995)、适应性好(黄长玲等,1993)、植株根系发达、茎杆韧性好(刘纪麟,1991)。研究表明温、热杂交种不但能适应温带地区,而且还拥有热带、亚热带种质的抗逆性好,生态适应性强,稳产性高等优点。为了有效地利用热带、亚热带群体扩增我国玉米种质基础,中国农科院作物所等科研单位相继从国外引进一批优良的热带和亚热带玉米群体,并对其进行适应性改良和中外种质的杂种优势研究。在温带地区种植热带、亚热带群体,由于对光周期的敏感性,往往表现晚熟、高秆、易倒伏、雌雄不协调、经济系数和产量较低,不能直接利用。因此对热带和亚热带种质的改良就成为其利用的前提。目前改良途径主要有以下几种:
1.混合选择。一种方法是将热带和亚热带种质在过渡地区进行适应性选择。在这种生态条件下,由于基因型和环境的互相作用,群体内会出现个体间性状的变异,可以从中选择适宜温带利用的个体,然后再逐步引种到温带地区种植,继续选择育成新的适应性群体或自交系。这一选择方法的利用已取得良好的效果。吴景锋(1978)将也门Tihama(北纬14度)的白粒综合种先引至Baitna(北纬16度3分)。在1978~1980年间,按照适合温带利用的要求,选择优良植株自交,从大群体中选留6个基本株的15份4代种子。1981年引至北京种植,其中仅3个系雌雄协调,继续自交选择,至1983年选出了适合温带利用、性状基本稳定的也铁19和也铁21两个自交系。四川农业大学(荣廷昭等,1998)采用混合选择法从Suwan1群体中选育出对西南山区具有特殊适应性的优良自交系S37,已被全国十几家育种单位引用,组配成多个优良杂交种。张世煌(1997)组织实施了一个从南到北的热带和亚热带种质接力改良计划。首先将热带、亚热带玉米群体分别在四川省和湖北省进行首轮改良,两年后将其传递给河南省和山东省育种单位继续作第二轮改良。目前已初步改良了19个群体,其中5个群体已适应了东北春玉米区的生态条件,并开始用于选系。另一种方法是对不适应的热带和亚热带种质直接在温带条件下进行改良,从中选择早熟、雌雄协调、抗病、抗倒伏的单株组建新群体。Hallauer(1972)从1963年开始对哥伦比亚复合种ETO进行六轮早抽花丝的混合选择,使抽丝期平均提前21天,株高穗位下降,获得自身产量和一般配合力效应都可以与玉米带群体相媲美的改良群体BS16。在我国,中国农科院作物所成功地改良了两个亚热带优质蛋白玉米基因库Pool 33和Pool 34在我国北方的适应性,自身产量大幅度提高。利用从改良群体中选出的自交系CA375与温带改良系齐205组配出中单9409(张世煌,1997)。吉林省农科院玉米所对泰国复合种Suwan1和Suwan2采用大群体混合选择法进行轮回选择,得到株高穗位适中、抗病性强,在吉林省基本正常成熟的RKⅠ和RKⅡ群体。以上说明,混合选择法是改良热带、亚热带玉米种质在温带条件下的适应性和光周期敏感性的有效途径。
2.温带与热带种质渗入杂交或回交选择。采用杂交或回交方法可以将热带、亚热带种质导入温带适应种质,创造半外来或部分外来种质。这种方法对改良基因型同质的外来种质的适应性和光周期敏感性十分有用。对少数基因控制的质量性状,如抗病性等,可通过杂交或回交的方法转移到温带种质中。中国农科院作物所和品种资源所合作(吴景锋,1991)通过构建和改良温热复合种质育成自交系中74-106,已组配几个优良杂交种通过省、市级审定用于生产。苏俊(1997)将Suwan1和墨黄9与甸11、系14、沈5003等自交系进行渗入杂交,共获得63份在温带可以利用的新材料,又经回交选择获得自交系材料400余份。山西省农科院通过热带种质导入选育出自交系太系131和太系19/02。河南省农科院利用含有50%热带种质的改良群体选育出自交系BT1,并育成“郑93-1”单交种。沈阳市农科所利用热带种质导入温带种质选育出自交系沈118、沈218、沈219,育成了沈试17、25和26单交种。在温热种质导入中,可能会出现某些优良基因的表达被适应性较强的温带种质基因掩盖的现象,出现综合双亲优良性状的个体较少,从中选择优系的概率较低等问题。为解决此问题,一般采取对半外来群体随机交配4~6次,以打破基因连锁的不平衡性,或利用温带种质适度回交,以缩小温热种质杂交后代的遗传变异,再进行选系。关于热带种质在半外来种质中的比率,不同学者的看法不一。一般的观点是适宜比率应因选择目标的不同而异,并取决于热带和温带种质自身的表现。
(二)玉米杂种优势群和杂种优势模式研究
利用杂种优势是提高玉米单产的主要措施之一。1918年Jones建议在生产上利用玉米双交种,为杂种优势利用创造了有利条件。到20世纪80年代初,美国单交种的种植面积占玉米播种面积的90%以上。我国现在杂交种普及率已达90%左右,产量获得显著增长。在杂种优势利用过程中,许多学者一直在探索揭示这一生物学现象的遗传机理,并提出了多种理论假说,这些假说亦从不同侧面部分解释了杂种优势机理。近年来,杂种优势群和杂种优势模式的研究备受重视,亦成为杂种优势研究的热点内容。
杂种优势群是指遗传基础广阔、遗传变异丰富,具有较多优良性状基因和较高产量一般配合力的种质基础群体;杂种优势模式亦指两个不同的杂种优势群之间具有较高的产量特殊配合力,相互配对成为强杂种优势模式(刘纪麟,2001)。依据杂种优势模式,从两个优势群中选出的自交系,相互组配产生强优势组合的几率亦相应较高。
美国是利用玉米杂种优势和研究杂种优势群最早的国家。迄今为止,在美国玉米带利用时间最长、范围最广的仍然是瑞得黄马牙和兰卡斯特两大杂种优势群和相应的杂种优势模式。两大系统又逐渐衍生演化形成几个较大的亚种质系统,如以Mo17为代表的C103系统(C103来源于兰卡斯特);以B73为代表的BSSS系统(在构建BSSS群体的18个自交系中至少有10个来自瑞得黄马牙);以Oh43为代表的西北马齿种系统;以W153R为代表的密尼苏达13系统和先锋公司杂交种选系的独特类群。典型的杂种优势模式包括瑞得×兰卡斯特;兰卡斯特×Oh43系统;瑞得×Oh43系统。近年来,兰卡斯特、瑞得系统与先锋种质选系的杂交组合也在上升。
在欧洲,硬粒型×美国马齿型是主要的杂种优势模式。在墨西哥以及中南美洲地区,以CIMMYT为中心的热带、亚热带地区的杂种优势群和杂种优势模式主要为墨西哥地方品种群体Tuxpeno和哥伦比亚合成群体ETO(Wellhausen 1978,Good-man 1985)。Vasal等(1992)利用自交系CML9(Tuxpeno血统)和CML21(ETO血统)按特殊配合力效应值将92份CIM-MYT热带自交系划分成两大群,然后分别将群内自交系经多次混粉重组成两个广基群体THG“A”和THG“B”,并建立A×B杂种优势模式。按同样方式,Vasal等(1992)又分别组建了亚热带杂种优势群STHG“A”和STHG“B”,以及配对杂种优势模式。在此基础上,有些学者又提出了一些其他杂种优势群和杂种优势模式,但多数基于上述模式。南斯拉夫的杂种优势模式为BS11(FR)C7×BS16(S2)C3和BSSS(R)C7×ZPEP。巴西的杂种优势模式为马齿型×Tuxpeno或Suwan1×Tuxpeno。阿根廷则以Flint×Recovered Flint为基本的杂种优势组合模式。
我国学者从20世纪80年代中期开始对国内玉米种质基础进行研究。吴景锋(1983)通过计算不同时期杂交种中主要亲本所占百分比,绘制了我国选育的主要二环系和部分外引系的系谱图。1986年王懿波等分析了河南省杂交种的种质基础。1990年曾三省分析了1978~1987年中国主要玉米杂交种生产和1984~1988年全国区域试验的资料,指出国内系×国外系类型杂交种的面积较大,并提出了杂种优势组的概念。1994年陈刚通过分析丹玉号杂交种的亲本组成,认为种质遗传基础主要来源于旅大红骨、兰卡斯特和瑞得三个系统。这一阶段大多数省份的玉米育种工作者均总结了各自种质利用状况,为进一步开展玉米育种工作奠定了基础。王懿波(1997、1999)对“八五”期间全国各省审(认)定的115个杂交种及234个亲本自交系进行遗传分析,结合育种实践,将我国玉米自交系分为5大杂种优势群、9个亚群;10种主体杂种优势模式和16种子模式。在1980~1994年我国玉米生产上利用的主要种质为改良瑞得、改良兰卡斯特、四平头和旅大红骨四大杂种优势群;利用的主要杂种优势模式为改良瑞得×四平头、改良瑞得×旅大红骨、Mo17亚群×四平头、Mo17亚群×自330亚群。
在利用系谱或地理来源分析方法划分杂种优势群的同时,国内学者也根据种质杂交后代的杂种优势表现和变异程度等数量遗传学方法和分子标记技术开展了杂种优势群研究。通过双列杂交方法分析自交系间特殊配合力,进而用于类群划分的方法有效且可靠,但工作量较大,使其操作性降低。彭泽斌等(1998)用此法将15个骨干自交系划为5个类群,证明了此法的可靠性。为减少工作量又使划群结果具有准确性,中国农科院作物所进一步采用NCII遗传设计对我国骨干自交系进行类群划分,证明目前我国主要使用3个杂种优势群和5个亚群,并确定5个亚群的标准测验种为四平头的黄早四、旅大红骨的丹340、瑞得的B73、PA种质的掖478和兰卡斯特的Mo17(图2-1)。进一步概括了我国玉米育种最常用的两个杂种优势模式。第一个是四平头×兰卡斯特,它有两种表现形式。在东北和华北春播玉米区表现为旅大红骨×兰卡斯特,其典型组合为E28×Mo17和Mo17×自330;另一个近似的表现形式为四平头×兰卡斯特,在黄淮海夏播区有一定的影响。第二个是四平头×PA种质,这是黄淮海夏播区的主要杂种优势模式,也表现为两种表现形式,其中一个是四平头×PA种质,典型组合为U8112×黄早四;另一个近似的杂种优势模式为旅大红骨×PA种质,典型组合为丹340×掖478。南方的种质复杂,杂种优势模式明显受北方的影响。但总的来说,我国的杂种优势模式具有当地×外来种质的特点,而且基本符合马齿型×硬粒型的规律(张世煌等,2000)。

图2-1 我国玉米种质的杂种优势群(张世煌,2000)
近年来,针对热带、亚热带与温带种质的杂种优势关系也进行了初步研究。李新海等(2000)采用NC-II设计研究10个热带、亚热带群体与我国玉米骨干自交系的杂种优势关系,认为Pob.43和Pob.21分别与Mo17、自330、丹340构成强优势组合,可能组成具有较大潜力的杂种优势模式。陈彦惠(2000)报道了我国温带玉米种质与热带、亚热带种质之间试验结果,提出自330×Pob.21、Suwan1×豫综5号、墨黄9号×黄早四、墨黄9号×旅9等杂种优势模式。韦国能(1998)报道,墨白1号(Tuxpeno 1),墨白94(Tuxpeno1p.B.C15),Suwan1和墨黄9(Amarillo Dentado 2)是20世纪80年代以来云南、广西生产种植的品种,并成为大面积种植的桂顶号、南顶号等品种的亲本,表明这几个热带、亚热带群体是适合我国南方育种利用的杂种优势群。刘纪麟等(1998)、张祖新等(1994)、卢洪等(1994)、刘勋甲等(1999)系统研究了我国三峡地区玉米地方品种的杂种优势群与美国种质BSSS-C9、Lancaster、热带种质墨黄9、Su-wan1的杂种优势模式;在此基础上,合成了地方种质、美国玉米带种质和热带种质各占适宜比例的4份复合种质群体:WBM、WLS、LBM、LLS。目前正在采用半同胞轮回选择法对4个广基群体进行改良。
我国西南地区拥有丰富的玉米地方种质资源,同时可直接引进与利用热带、亚热带和北美温带的优良种质,因此非常适合开展玉米种质扩增、改良与创新研究。在评价地方种质和外来种质农艺性状的基础上,发掘优质、抗逆、高配合力核心种质,研究两类种质的杂种优势关系,构建广基复合种质群体,并用于相互轮回选择改良,从而开发和创新我国玉米杂种优势群和杂种优势模式。
(三)玉米雄花不育的研究与利用
玉米是世界上最早利用雄性不育系生产杂交种的作物之一。20世纪70年代由于T型不育系对玉米小斑病T小种高度专化感染而造成小斑病爆发流行,使玉米雄性不育系的研究与利用曾一度陷入低潮。但由于常规制种需要大量繁重的去雄工作,若去雄不彻底,则会出现自交苗,大幅度降低产量。大致估算,F1代自交率若占10%,则杂交种增产潜力约降低5%;若自交率超过15%,则杂交种增产潜力将损失殆尽。利用强优势的雄性不育系配制杂交种是保证种子纯度提高玉米产量的重要途径。因此玉米遗传育种工作者一直非常重视雄性不育的研究与利用,并取得了一定进展。
1.玉米雄花不育的类型。在自然环境下,有许多因素可引起玉米雄花不育,如高温、缺水、辐射以及营养元素缺乏等,但这种类型不育无法稳定遗传,在生产上没有利用价值。受遗传控制的不育性按遗传特点可分为两大类,一类是细胞核雄性不育;另一类是质核互作不育。细胞核雄性不育受核基因控制,分显性和隐性不育两种。目前已发现4个显性核不育基因Ms和22个隐性核不育基因ms。质核互作雄性不育由细胞核不育基因与细胞质不育基因互作而共同控制。细胞核和细胞质基因均为不育基因S(rfrf),表现为不育,称为不育系;细胞核基因型不育而细胞质基因可育N(rfrf),表现为可育,父本可保持不育系的不育性,称为保持系;细胞核基因型可育N(RfRf)或S(RfRf),表现为可育,父本可恢复不育系的育性,称为恢复系。
2.细胞质雄性不育分类。细胞质雄性不育,可按细胞核基因对细胞质基因不育性的恢复专效性进行分类。Beckett(1971)用具有不同恢复基因的4个自交系将不同来源的细胞质雄性不育品系分为T、C、S三组(图2-2)。T组为孢子体雄性不育,败育时期较早,败育彻底。1970年,含T组不育细胞质的雄性不育杂交种占美国玉米总面积的75%~80%。由于遗传基础单一,造成当年专化侵染T组细胞质的玉米小斑病T小种大流行,全美国玉米严重减产。目前,T组细胞质雄性不育已基本停止使用。S组为配子体雄性不育,败育时期较晚,不育性的稳定性较差。S组细胞质没有发现小斑病及其他病害的专化侵染,目前发现的细胞质雄性不育品系大多属于S组。
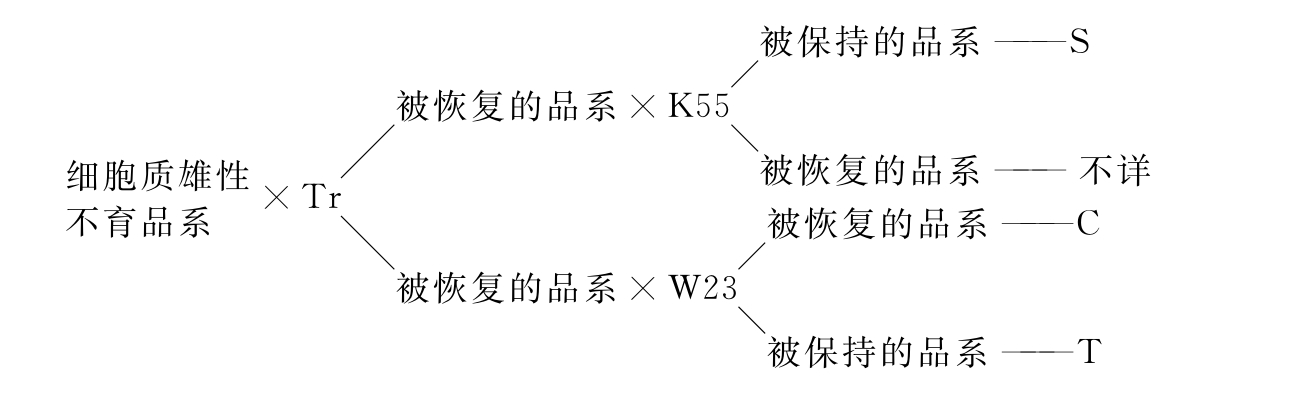
图2-2 细胞质雄性不育分类(Beckett 1971)
我国学者郑用琏(1982)根据Beckett恢复专效性原理,研究了国内若干细胞质雄花不育系的大田育性反应,提出了一组相应的胞质分类体系(图2-3)。凡能够被恢313恢复的不育系属于S组;被恢313保持而被自凤1恢复的属于C组;对恢313和自凤1均表现不育的属于T组。
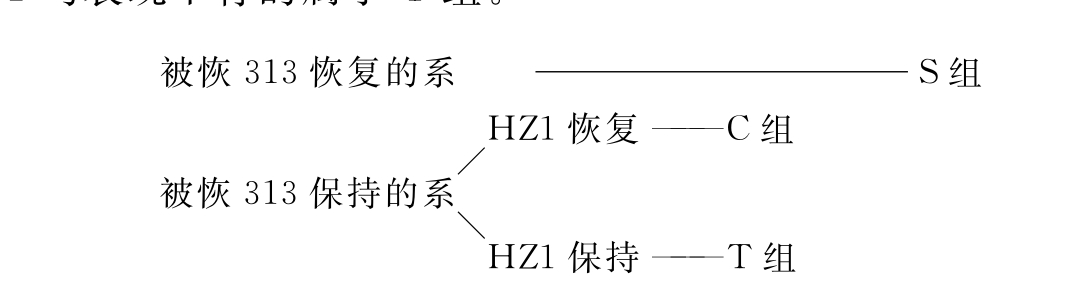
图2-3 我国雄性不育胞质分类体系(郑用琏1982)
温振民(1983)研究认为,依据不同不育胞质的特殊表型效应,可采用综合鉴定逐级分类。具体程序如下:首先在室内或田间鉴定不育系是否抗专化性侵染T型细胞质的玉米小斑病T小种,确定其是否属于T组;其次在抽雄后按不育系花药、花粉败育情况初步确定是否属于C组(孢子体雄性不育)或S组(配子体雄性不育);最后用特定自交系测交,按F1及其子代育性的恢保关系进行分析,最终按综合鉴定结果进行归类。
李小琴(2000)利用一套同核异质的不育系测定了55个新选系对不同不育胞质的大田育性反应。鉴定出1个能恢复T、S、C三种不育胞质的全效恢复系“HZ32”,以及C组的专效恢复系“吉6759、P111等”;S组的专效恢复系一恢“313、801”和对C组和S组均具有恢复能力的双效恢复系“S7913、牛2-1等”。确定了一组测验种集团,提出了三套玉米CMS育性专效分类系统,从而更加完善了我国不育胞质分类体系。
3.细胞质不育恢复基因的定位。1995年,李竞雄等利用B-A易位系将隐性细胞核雄性不育基因ms30初步定位于第4染色体长臂上,距甜质胚乳基因su 15~20厘米;1996年,又将ms2细胞核雄性不育材料与引进的黄绿苗基因材料进行杂交、分离,育成了带有黄绿苗标记的细胞核雄性不育系和保持系,为细胞核雄性不育系的利用探索出一条新途径。
随着高密度玉米分子标记图谱的发展,细胞质不育恢复基因的定位研究亦进入分子水平。Wise等(1994)应用RFLP标记将Rf1基因定位于染色体3的UMC153和UMC192之间,图距分别为1.2厘米和9.5厘米;同时将Rf2基因定位于染色体9的UMC153和UMC95之间,图距分别为3.8厘米和7.1厘米。Kamps等(1995)利用RFLP标记将Rf3基因定位在染色体2的BNL7.14和whp1之间,图距为6.4厘米。石永刚等(1997)利用RFLP和RAPD标记进一步将Rf3基因定位在UMC49和RAPD E08-1.2之间,图距分别为2.7厘米和4.8厘米;Sisco(1991)将Rf4基因定位在RFLP标记BNL114附近,图距为1.5厘米。恢复基因的定位研究为进一步开展恢复基因的分子标记辅助选择和克隆工作提供技术条件。
4.光、温敏感性雄性不育材料的发现。赫忠友于1992年首次发现温敏性雄性不育玉米材料,经过5年的观察研究进一步得到确定,并取名琼6-Qms。这一材料的特点为:在雄穗发育的特定时期,温度条件能导致雄花育性的转换,即在25℃以上的条件下,雄穗花药发育不良,药内花粉大部分败育,内外颖呈闭合状态;在25℃以下,基本表现正常开花散粉。这一特性由隐性基因控制,能稳定遗传。目前,通过育种程序已获得了不育性稳定的琼42-Qms及其可育近等基因系;并获得了一个与之连锁的RAPD标记。琼42-Qms的选育为采用温敏核雄性不育“两系法”配制杂交种提供了可能性。但由于在自然条件下温度变化的复杂性,如何有效地利用温敏感性雄性不育有待于进一步研究。
5.育成了一批高抗玉米小斑病T小种的新型不育系。针对T型不育系对玉米小斑病T小种高度专化感染的问题,1970年以来,国内部分育种单位先后育成了一批高抗玉米小斑病T小种的新型不育系,其中包括辽宁省昭乌达盟农业科学研究所的双-cms;华中农业大学的唐徐-cms;辽宁省农业科学院的L2-cms;河北省农业科学院的21A-cms等。
近年来,为了提高玉米杂交种子的质量,国内一些玉米育种单位加强了玉米雄性不育系的利用步伐,并取得了可喜的成就,已有部分品种实现了不育化制种,如C豫玉22、农大3138、华玉4号、C73单交种、S中单2号等。
(四)品质育种新进展
1.优质蛋白玉米。自1962年Mertz等发现软质胚乳的o-paque(o2)突变体赖氨酸含量达到0.4%以后,许多国家的玉米遗传育种工作者开始了高赖氨酸玉米品种的选育工作。但因o2玉米籽粒容重小、病害重、产量低等原因于20世纪90年代初期停止了此项研究,如美国、南斯拉夫等。我国的高赖氨酸玉米以及优质蛋白玉米(Quality Protein Maize,QPM)在李竞雄先生的支持下,经过20余年的联合攻关,已取得了不少成绩。
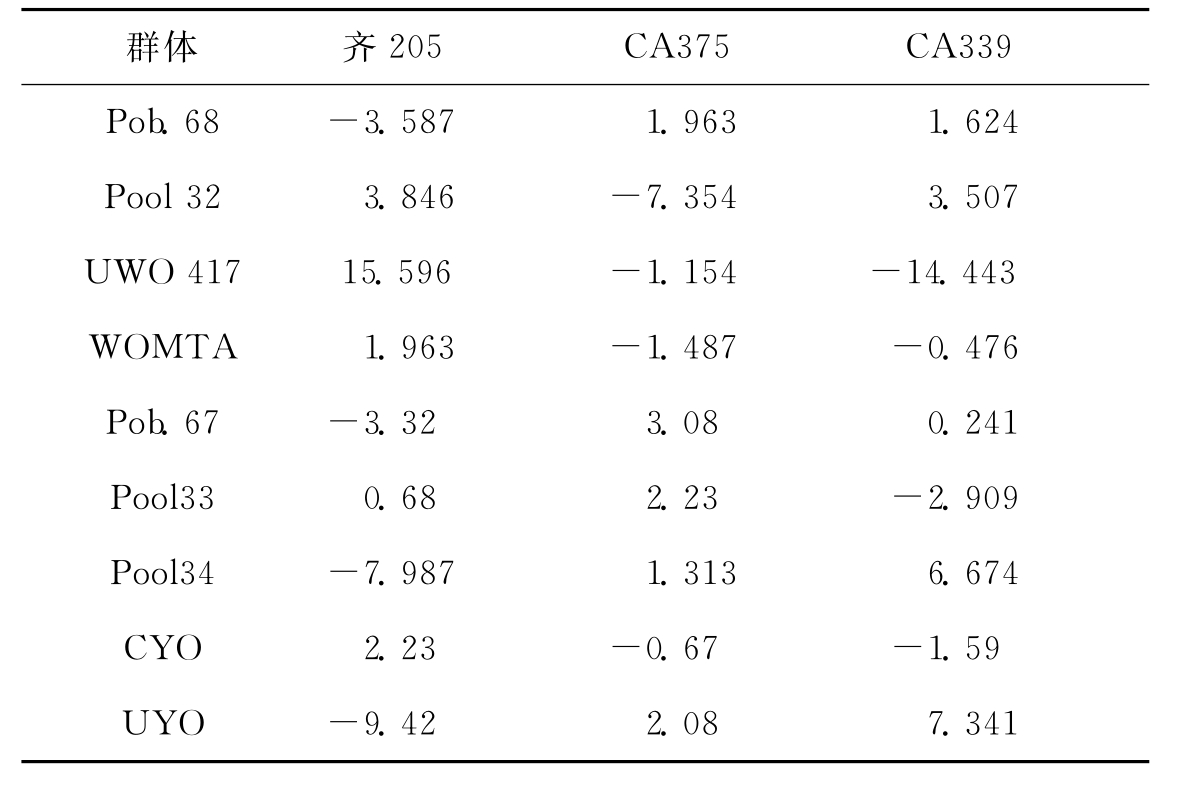
截止目前,通过利用控制双亲的早抽花丝混合选择法改良了5~6轮亚热带QPM基因库Pool33和Pool34,获得了温带适应群体中群13和中群14。同时对从CIMMYT引进的QPM群体Pob.67,Pob.68,Pob.69和Pob.70进行了2~3轮选择,基本适应了黄淮海玉米区的生态条件。利用NCII遗传交配设计,研究了我国QPM核心种质(CA335、CA339和齐205)与22份CIMMYT QPM自交系及其种质群体的杂种优势关系(表2-1),在此基础上通过多2~3次混粉合成了66份半外来种质。通过分析我国QPM种质基础,认为中群13和中群14是一对杂种优势模式,其中CA375和齐205可作为划分QPM种质杂种优势群的测验种,指导种质改良。我国QPM育种的技术路线可概括为:利用o2基因和胚乳修饰基因分别提高胚乳中的赖氨酸含量和籽粒硬质度;其次引进新的QPM杂种优势群,提高杂种优势利用水平,开发和创新我国适用的杂种优势模式。育种目标可归纳为在保持全籽粒赖氨酸含量0.4%左右的基础上,以半硬质或硬质胚乳为选择标准,改善籽粒的物理性状;提高选育品种的适应性和抗病性,尤其是对穗腐病的抗性;选育的杂交种产量水平与生产上的优良品种持平或增产5%~8%。在注重QPM新品种选育的同时,亦应加强配套高效栽培技术和饲料配方技术的研究,使QPM新品种选育、高产栽培和饲料加工相结合,实现育、种、养、加一体化体系。
表2-1 9个CIMMYT QPM群体与我国3个QPM
测验种的特殊配合力效应(李明顺2000)
1980~1990年我国选育出了一批籽粒赖氨酸含量0.4%左右的硬质或半硬质杂交种,如中单9406、中单9409、鲁玉13、鲁单206等。
2.高油玉米。美国于20世纪40年代后期最早开始高油玉米育种工作,先后创建了高油群体IHO、ASK、RYDH、Syn.D.O、YUSSS、DSTU,其中ASK群体遗传基础广泛,经过27轮选择,含油量达到了21.2%,成为目前高油玉米育种的主要种质资源。但直到70年代才育成一批高油自交系品种。由于除油分外的其他性状不如普通玉米,所以高油杂交种推广面积较少。
20世纪80年代初,中国农业大学宋同明教授从美国引进了高油玉米种质,开始了我国高油玉米遗传改良研究。经过近20年的潜心工作,创造性地发展了高油玉米种质资源,至1999年,已对北农大高油群体(BHO)完成了15轮选择,含油量达到13.9%;对引进的高油群体RYD和Syn.D.O分别进行了9轮和7轮选择,含油量分别达到15.2%和13.1%;对从IHO-C80与ASK-C23杂交F3得到的重组高油群体AIHO完成了10轮选择,含油量超过18%(宋同明2000)。
玉米籽粒的含油量受微效多基因控制,而且表现出较高的遗传力,所以轮回选择是创造高油玉米新种质的有效方法。在现阶段,利用高油玉米群体选育自交系是最有效的途径。在育种目标上,高油玉米的含油量应保持在6%~8%,农艺性状优良,抗主推地区的主要病害,产量应与生产上的推广品种相近或增产5%~8%。
目前,中国农业大学已将有关高油玉米向美国转让专利。近年来相继选育出含油量7%~9%的高油玉米杂交种,如高油115、春油298等,已用于生产和饲料产业。
3.其他特用玉米。我国的糯玉米、甜玉米和爆裂玉米育种相对来说起步较晚,但为适应市场需求,发展迅速。近年来,相继育成一批玉米杂交种应用于生产,如中糯1号、苏糯1号、垦粘1号、白糯1、2号等;甜玉2号、甜单8号、金银99、东甜3号等。优质品种的育成,为我国种植业结构调整奠定了基础。
(五)生物技术育种开始启动
生物技术育种是进入20世纪80年代中期以来发展起来的,在DNA水平上实施作物改良计划的理论和方法,主要包括基因工程育种技术和分子标记辅助育种技术。基因工程研究是克隆目标基因,同时利用转基因技术培育转基因玉米品种。这些外源基因可能是来自其他生物,也可能是在实施室中人工合成的。分子标记技术的研究目标是鉴别控制性状的基因,获得紧密连锁的分了标记,并与常规育种技术相结合建立分子标记辅助育种技术。与常规育种方法相比,分子育种技术真正实现了对目标基因的操作。
玉米成为生物技术研究的主要对象有两个明显的理由。从全球范围来看,玉米具有极其重要的经济价值;另一方面,商品玉米生产广泛使用杂交种,种子公司每年能够从销售种子中获取高额利润。
1.基因工程技术。虽然常规技术仍然是玉米育种的主要方法,但是生物技术的迅速发展为育种家提供了新的技术手段。用转基因技术培育的转基因玉米杂交种(抗虫和耐除草剂品种)已经在发达国家投入生产使用。抗虫转基因玉米具有天然的抗虫性,农民就可以节省购买杀虫剂的投入,而且对环境有好处。美国种子公司Novartis和Mycogen已经获准在美国生产和销售Bt玉米。关于Bt玉米可能对环境和人类造成的有害影响已经在许多国家引起广泛关注和争议。玉米基因工程研究的第二个方向是耐除草剂品种选育。选择性除草剂只毁灭杂草而不伤及农作物,在现代玉米生产中发挥了重要作用。在生产上使用耐除草剂转基因玉米必须长期依赖该公司所生产的广谱除草剂,这是发展中国家不愿意接受转基因玉米的一个重要原因。1999年全球转基因玉米种植面积为280万公顷。在美国,1/3的玉米种植的是耐除草剂或抗虫的转基因品种。
在我国,转基因育种技术在玉米上的应用相对于水稻、番茄等作物来说,起步较晚,取得的成果相对较少。现已报道的主要有中国农业大学培育的Bt玉米(王国英,1995);1998年进入中间试验,1999年开始在北京进行环境释放,预计4~5年后有望进入商品化生产阶段。
2.分子标记技术的应用。目前,分子标记技术主要有两大类,即以分子杂交为基础的(如RFLP等)和以PCR扩增为基础的标记(如SSR、RAPD、STS、AFLP等)。分子标记技术在玉米育种上的最重要应用是划分杂种优势群。截止目前,美国、德国和CIMMYT利用RFLP或SSR标记技术分别研究了200个、200个和400个左右主要玉米自交系的遗传多样性。中国农科院作物所首先利用RFLP和SSR标记研究了我国29份骨干自交系的杂种优势群划分,并筛选出扩增带型清晰、遗传多态性丰富的61对SSR引物;进而初步将160份我国主要玉米自交系划为6个类群,即四平头、旅大红骨、Lancaster、Reid、PA和PB,并将未知系谱关系的自交系划分到相应类群中;划群结果与育种家按系谱分析结果基本吻合。研究结果为我国玉米育种及种质改育提供了重要理论依据和技术基础。
分子标记技术可用于建立品种的DNA指纹图谱。利用DNA指纹图谱可以很精确地把遗传差异很小的植株区别开来。因此这项技术在植物新品种保护领域特别有应用价值。目前我国已初步建立了一套以SSR标记为主的玉米新品种分子测试技术体系,主要包括快速提取籽粒或叶片DNA→通过分析DNA指纹图谱数据,选择特异性SSR引物进行PCR扩增(或多重PCR)→测序凝胶电泳→扩增片段读取(通过凝胶识别软件)→数据处理及统计分析→杂交种纯度鉴定。这套检测技术尚需进一步程序化、标准化。应用DNA分子检测技术可以较快速、准确地鉴定玉米杂交种的纯度。它可以用作品种真实性鉴定、纯度检测以及新品种保护的仲裁技术依据之一。
采用分子标记技术对玉米重要经济性状进行基因定位和分子标记辅助育种已成为当今玉米遗传改育的重要研究方向。目前在我国正在研究的课题包括玉米对病毒病、茎腐病、弯孢菌叶斑病的抗性基因定位、耐旱、耐酸性状的分子遗传分析、玉米杂种优势的遗传机理、玉米细胞质雄性不育恢复基因Rf3的基因定位、玉米胚乳突变基因su2和du基因定位等。分子标记辅助育种技术研究亦处于起步阶段,许多育种单位都在开展此项研究工作,如中国农业大学、中国农业科学院、华中农业大学、中国科学院遗传所等,在技术储备方面已经取得一定进展。石永刚等(1997)找到了与玉米S组育性恢复基因Rf3相连锁的两个标记,一个距离为4.8厘米的RFLP标记,一个相距为2.3厘米的RAPD标记。李立家(1998)利用RFLP标记将两个玉米大斑病抗性基因Ht1分别定位于第2染色体上,此外在第7染色体上也发现有同源序列。杨文鹏(1998)利用具有o-6基因遗传背景的玉米材料筛选出与o-6位点连锁的RFLP标记。这对于优质蛋白玉米材料的筛选及育种,以及o-6基因的分离、克隆具有重要意义。中国农科院作物所(2002)对玉米矮化叶病抗性基因进行了QTL分析,分别在第1、3、5、6、10染色体上找到了QTL及相应连锁标记,并确定3、10染色体上存在主效位点。此外,中国农业大学找到了与抗玉米肿囊菌青枯病基因距离2厘米以下的SSR分子标记;中国科学院遗传研究所分别将禾谷镰刀菌和肿囊腐霉菌引起的青枯病抗性基因Rfgl和Rpil基因定位到第6和第4染色体上,并找到了与其连锁的分子标记;山东大学确定了玉米粗缩病抗性至少由3个遗传位点决定,分别位于不同染色体上,其中第6染色体上的遗传位点起关键作用。该位点与探针umc21、umc46和umc152c紧密连锁。对玉米重要经济性状的分子标记和基因效应分析,为分子标记辅助选择和聚合育种技术研究提供条件,也推动了我国玉米分子育种技术的开展。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











