



古人类学家使用多种方法来探究新发现化石的科学意义。他们把人类化石归入到特定的、可再分类的类群,探明了它们与其他化石和现生类群的关系并重构化石人类的生活习性。
分类与分类法
西方科学界采用瑞典博物学家林奈在1758年创立的分类体系来给生物分类。种是这一体系的基本单位,由一群形态相似、相互能持续繁殖后代的动物组成。单一动物个体都可归入一个物种,相似物种构成一个“属”,相似的属构成“族”,相似的族构成“科”,依次向上类推到“界”。现代人、智人都属于智人种,直立人属,人族。
“命名法”是分类学的一个分支,专门规定怎样用林奈分类体系给生物命名。命名是有一套规范的编码,科学家给一个新发现的物种命名都要遵守这一编码。这一编码规定了新种或新属的名称规则。商业产品的名称被禁止采用,例如,不允许使用Burgerking(汉堡王)ipodensis作为新人种的双名制名称。“双名制”确保现有的分类不被新分类所取代,可以避免产生混淆。
研究者确定出一个新物种时,需要选择一件化石作为这个种的“模式”标本。通常的选择是最初发现的一件保存相对完好的化石标本,而这件标本无须是这个种类中最典型的成员。模式标本的重要性在于它承载着不可改变的分类单元名称。因此,举例来说,如果尼安德特人的模式标本被发现不同于其他所有的尼安德特人标本,那么,就需要给其他标本另外一个新的种名。尼安德特人的名称不能脱离它的模式标本,模式标本的名称不能再变化。如果研究者最终决定应给予某一标本一新种名,那么这件标本就是一个新种的代表。命名法有优先命名的规则,即如果两件模式标本被发现是同一个种,那么种名只能沿用先命名的那件标本名。
就像“种”是一个类群,在林奈体系中所有类别都是类群;而研究者描述某“类群”时,他们通常指的是一个“物种”。描述“物种”在等级的包容性越来越强(即物种的群体越来越大)的体系中是如何排列的科学被称为“分类学”,即字面上的“分类体系”。分类分析是确定人类化石应该被置于哪个分类单元的过程。首先,研究者必须确定新发现的化石是否属于一个现有的人类类群。只有当他们确信它不能被归入到一个现有的物种时,才考虑给这个新物种确定一个新种名。同样的原则也适用于所有林奈的分类阶元,因此,如果确信新物种不能被现有人类的属所容纳时,研究者应当另外建立一个新属,依此逐渐向上类推。
下面介绍的分类学分析方法与其他方法都是基于对一件化石形态的详细评估。化石的形态包括外部形态和内部结构。形态可以是肉眼看到的、整体的大结构,也可以是微观的、需要在各类显微镜下才能看到的小结构。研究人员不仅对化石的大小和形状作了详细的定性分析,而且也试图提取标本的测量数据进行定量分析。定量描述的最简单的形式是测量化石上两个解剖标志间的距离,即线性测量。目前,研究者借用医学影像的激光束和其他技术,能够比过去更准确地提取到化石外部形态的和内部的细微结构。圣路易斯华盛顿大学的古人类学家格伦·康罗伊(Glenn Conroy)和医学影像学专家查尔斯·维尼尔(Charles Vannier),率先使用电脑断层成像(CT扫描)研究了出自非洲南部汤恩(Taung)的一件化石人类颅骨的内部结构。随后,乌得勒支大学的医学影像学专家弗朗斯·宗尼维尔德(Frans Zonneveld)和伦敦大学古人类学家弗雷德·斯普尔(Fred Spoor)改进了这些方法,他们现在能够提供内耳的结构信息。研究人员利用这些数据确定人类化石的种类,重建其体态和听觉。
研究者必须确保化石的测量能够准确反映出骨骼或牙齿在形成石化前的大小和形状。如果每天暴露于热和冷交替环境一段时间,骨骼和牙齿就会产生裂缝。岩石碎屑进入到裂缝里,就增大了骨骼或牙齿的尺寸。同样,如果一件化石骨骼在石化前后若裸露于地面干燥和多风的环境下,风挟带着沙粒会产生“喷沙”效应,从而会去除皮层质骨的部分外皮层。这样的侵蚀作用就会使骨骼化石的尺寸变小。一件新发现的化石的测量数据和非测量性状可与现有的化石类群进行相似度比较。亲缘关系相近的现生动物(如人科中的现代人类与非洲猿)通常被用来作为模型,以帮助决定在一新化石种类中可以允许有多少变化。纽约大学研究灵长类的专家克里夫·朱利(Cliff Jolly),花了30年时间研究不同狒狒群之间的分界;他认为,狒狒和其近亲在某些方面是研究人类进化更好的参照样本。他同时指出,狒狒不仅比黑猩猩和大猩猩分布广泛,而且在近期的进化史中的模式和时机方面都与人类的相似。
化石碎片的复原
几百万年的人类化石很少能处于完好的状态。头骨和面骨尤为脆弱,容易受到有蹄动物的践踏或洞顶落下的岩石挤压而破碎,有时剩下的只是颅骨的一小块碎片。在少数情况下,也许会发现多块碎片;但如果这些碎片太小,复原就是一个挑战。拼合工作就像完成一个有许多天空插片但没有云彩的三维拼图,而且也没有图画可参照。一种选择是竭尽全力地手工拼合它们,但即使是熟悉头骨每一个细节的专业解剖学家也要花费数百个小时。
苏黎世人类学研究所的玛西娅·庞塞·德·李昂(Marcia Ponce de León)和克里斯托福·朱里库佛(Christoph Zollikofer)都是新兴研究领域——“虚拟人类学”——的专家。他们利用计算机的优势和软件设计的进步发明了一种替代手工复原人类化石的方法。化石经过激光扫描,在计算机屏幕上呈现出一个“虚拟”的化石形态。研究者可以沿任何方向移动和旋转每一件碎片,以找到相对应的部分。该软件还可通过镜像成像根据一侧颅骨信息获得另一侧缺失的对称部分的图像。朱里库佛和李昂最近已经使用这一方法虚拟重建了可能属于早期人类的撒海尔人乍得种的颅骨。同类软件与CT扫描一起使用可清楚地看到隐藏在颅骨深处的结构如上颌窦、内耳的骨管或牙根等。
年龄和性别的鉴定
即使有完整或者接近完整的骨架,确定人类化石遗存的性别和发育年龄仍然可能相当困难。当这些遗存都是很小的颅骨碎片时,鉴定的难度就更加大。化石个体停止生长的年龄比较难精确确定。牙齿的发育程度可以帮助确定未成熟个体的年龄;一旦所有的牙齿都萌出,牙根形成,就不能用牙齿确定其年龄了。
骨头和牙齿的大小与形状,肌肉留下的深浅痕迹以及骨盆的大小和形状(虽然骨盆碎片在人类化石中非常稀少)是确定化石个体性别常用的方法。基本假设是,因为许多非人类灵长类动物中,雄性的体格基本都大于雌性,那么,在早期人类中,男性的体格也同样应大于女性。这是性别二态性的一个方面,二态性指与性别有关的个体间的所有差异。然而,当你处理一个包含信息量很少的化石记录时,个体的大小并不总是性别判定的可靠标尺。
如果把现代人类的性别二态性简单推演至早期人类身上,就会增加研究的复杂性。例如,在现代人类中,许多骨盆的性别二态性的出现是两种需求折中的结果,即要满足两足直立行走的需要,也要满足生育大脑袋婴儿要有较宽骨盆的需要。但是,相同的二态性未必适用于小脑袋的早期人类,他们与现代人类两足直立行走的方式不同,早期人类的骨盆的性别二态性可能会表现为一种独特的模式。
种及其鉴定方法
生物物种概念是最广泛使用的对物种的科学定义,由哈佛大学已故杰出的进化生物学家恩斯特·迈尔(Ernst Mayr)提出。生物种是指“与其他群体在繁殖方面相隔离的一个自然群体”。当你有机会观察现生动物并了解其交配情况,就很容易对生物种加以区分。但是,这种方法显然不能用于鉴定化石种。不过,因为相同种的成员之间才能够交配繁殖,异种之间不能交配繁殖,所以同种之间的相似性比异种之间更为密切。这样,在缺少交配习性信息时,我们可以通过化石的外观、结构和基因组成(如有DNA保存)来确定化石的种类。
然而,要用这些方法来确定化石的种类还存在着一些问题。第一个问题是,我们发现的人类化石中并没有完整的生物躯体。我们习惯把生物体的组成分为两类,一类是软组织,如肌肉、神经、动脉等;一类是硬组织,如骨骼和牙齿。人类远祖化石仅限于硬组织的遗迹,而且许多都是骨头和牙齿的碎片,因此问题是,当仅发现了几块磨损的牙齿或一块颚骨或部分股骨时,古人类学家如何鉴定这些化石的种类。
第二是时间问题。每个物种都有一段生命史,即有一个形成、生长和消亡的过程。物种要么灭绝而没有留下任何直接的后裔,要么成为一个或多个新“继承种”的共同祖先。化石哺乳动物的种一般能延续到1至2百万年。经历这么悠久的历史,物种的外形不可能保持不变。偶然的变异和气候变化引起的形态改变都将引起物种的改变。但只要种的个体仅与同种的个体交配繁殖,则该种还能保持它的独特性。即使科学家花费一生的时间观察一个现存物种,也只能是这一物种自存在以来的一个瞬间。因此,在博物馆藏品中看到的,百年来采集的现代种的骨架并不是一个模式种,你无法参照它来鉴定一份采集于数十万年前的遗址中的化石样品。
此处用长跑比赛来比喻很恰当。一件化石就像长跑比赛中的一张静止的画面。长期存在的物种可能在其历史中被数次取样。古人类学家需要找出办法鉴别他们观察的是同一赛跑中的几张照片,还是几个不同比赛的照片。在研究人类进化时这意味着应先观察现代人类和高等灵长类动物的骨骼,然后参考这些现生类群的形体大小和形状的变量来推定在单一物种化石中应容许多大的变异。如果变异少于现生类群,那么,有充分的理由来断定,在这批化石中代表的是一个种。对于涉及时间跨度大的化石标本,古人类学家在宣布其变异太大而不能包含于一个单一物种之前,会试图对化石样本中可能的变异数量进行有根据地预测,但要声明,这仅仅只是一种预测。
要确定在早期人类化石中有多少个种则更加困难,因为人类包括化石人类的生物学变异持续存在。因此,化石类群的分界往往是科学推断和争鸣的结果,新化石的发现或新分析方法的引入往往意味着须重新分类,或须重新考虑原有分类和标准的适用性。只有当有充分的理由相信新化石种不属于现有的物种时,那就应该建立一个新种。而要建立一个新属,则需要有更强有力的证据。
物种的形成
一些研究者认为新种是整个种群逐渐演变的结果;这种理论被称为“种系渐变”,相应的物种形成模式被称为“进化”。另一部分人认为,物种的形成是一些生物亚群体由于地理上的隔离而在进化过程中发生突变的结果;这种解释被称为“间断平衡进化”。在后一种解释中,突变之间的长间隔期中形态演化应没有持续的趋势,只是形态学上的“随机”波动;物种形成的这一模式被称为“分支演化”;“静态平衡”一词用来描述物种形成过程中的形态的稳定时期。几乎所有研究者现在都接受,进化过程中的大部分的形态变化都发生在物种的形成时期。
在某些情况下,物种的形成可能是由于染色体的重排而引起的基因型的大规模变化。研究者认为,这可能是高等灵长类物种形成的潜在机制。
种的形成与种多样化密集期被称为“适应性辐射”,往往是因为出现了一个开拓新环境的机会,或因为其他种群的灭绝意味着适应性机会在现有的环境中成为可能。在类似阶段一些族系往往比其他族系能产生更多的物种,因此,具有较高的“物种丰富度”。
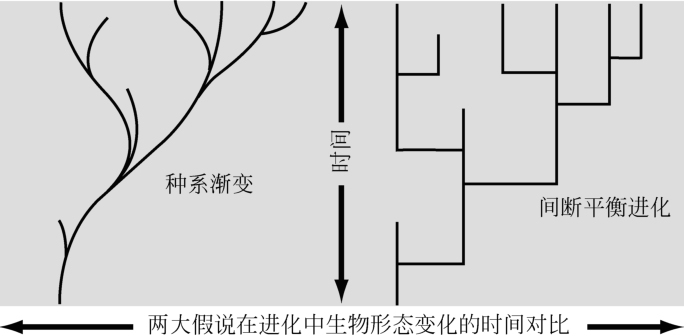
图6 “种系渐变”(左)和“间断平衡进化”(右)两大假说关于进化中生物形态变化的时间对比
所有物种,包括现代人类,最终都将灭绝。不确定的是,灭绝是由物种内在的特性决定的,还是由物种外在的因素如环境的改变决定的,或内外两种因素兼而有之。这些竞争性假说的验证可以通过在实验室里改变果蝇等迅速进化的生物体的环境条件,或观察生物的发育情况来实现,也可以通过把化石与过去气候变化的直接证据相比较来实现。
主分派和主合派
本书使用的分类法倾向于分出相对多的人种,并非所有研究者都赞成分出如此多人种。赞成分出多个人种的学者被称为“主分派”,相反,中意于综合人种的学者被称为“主合派”。两组科学家研究同样的证据,只是各自的解释不同而已。古人类学家对化石人种数量认定的不同源于他们对化石变异解释的差异。强调化石记录连续性的学者一般“主合”,而强调化石记录非连续特点的学者一般“主分”,但说来说去,所有的分类法都是基于假设。如果一些科学家用一种分类法研究了某件标本,其他科学家也能够以不同的分类法重新研究这件标本,只要各自能清晰地把标本归入他们认定的分类体系。
表2 主分派和主合派对化石人类分类假说的对比

续表

(续表2)
支序分类分析
新发现的化石一旦归入一个类群之后,研究者就会进行下一步,应用支序分类学推导这个化石人类与现代人类以及其他化石人类类群间的关系。
“进化支”包含拥有同源祖先的所有生物体。最小进化支只由两个类群组成;最大的进化支包括所有现生的有机体。支序分类分析根据共同的形态性状数量来划分类群,但形态性状需要具有一定独特性。为了更好地了解两个种之间的亲缘关系,所选的形态性状必须是两个或多个类群的共有特征,但也能要反映出类群内的差别,以便能再划分出次一级的组群或较小的分支。举例来说,所有高等灵长类都具有哺乳动物的一些特征,如拥有乳头和温血,但对厘清巨猿间的关系并无助益。另一方面,只在一个类群所具有的形态性状,也不能用来确定与其他类群间的亲缘关系。
两个类群若共有某特定的形态性状则被称为姊妹群。这对姊妹群本身又有自己的姊妹群(例如,大猩猩是黑猩猩和直立人分支的姊妹群等。)表述上述分支关系的图叫“分支图”。这一姊妹群关系也可以用套括号表示,例如,(((人属,黑猩猩属)大猩猩属)猩猩属)。
支序分类分析先假设,如果两个类群有共同的形态性状,那么它们都是继承于前一个共同祖先。这个假设往往论据充分,但也有例外。我们知道,灵长类动物包括高等灵长类经历了趋同进化,不同谱系能各自独立演变出类似的形态。趋同是指在两个物种上出现类似的形态性状,但这一性状不是源于前一个共同祖先,即非同源相似。举例来说,厚牙釉质性状在人类进化过程中很可能不止一次出现,于是成为人类分支中的一个趋同现象。
化石DNA
分类分析的最新形式是依靠提取和分析DNA(脱氧核糖核酸)来找出人类类群之间的相关性。在一个家庭中,亲缘关系密切的个体如兄弟姐妹具有比亲缘关系远的如堂兄弟更多相同的DNA。类群也是如此,同一类群的个体间比不同类群的个体间平均拥有更多相同的DNA。然而,尽管DNA在我们生命中具有重要性,石化会很快降解核酸。比如,5万年后只有少量的DNA存留下来,而且剩下的只是DNA片段。德国莱比锡马普进化人类学研究所的分子生物学家施温提·柏保(Svante Pääbo),率领他的研究小组首先从人类化石中发现了DNA,在本书第7章介绍尼安德特人时将讨论化石DNA证据的作用。做化石DNA分析的研究者,要特别注意防止和发现其所受的污染。当人们处理化石时,难免会在化石上留下头发和皮肤细胞等潜在的污染源。科学家们必须确保,他们观测的DNA来自人类化石而不是其他途径。在最近的一项研究中,研究者在一件洞熊化石上检测到了20多个不同的现代人类DNA序列。大多数人类化石都被几十甚至几百人接触过,尤其是许多年前发现的化石,因此,在一件现代人类化石中分离出属于化石本身DNA序列的工作会具有很大的挑战性。
等级划分
“趋同现象”使按进化支划分早期人类变得复杂。一种替代方案是把人类类群分成等级。一个等级就是基于动物行为本身而不是它的系统发育关系的分类阶元。举例来说,越野车相当于一个等级,而包括越野车在内的所有由福特汽车公司生产的汽车相当于一个进化支。等级可以是进化支,但并不一定如此。举例来说,食叶猴属于一个等级,但不是一个进化支,因为旧大陆的食叶猴和新大陆的食叶猴都是各自大陆更大猴类进化支的成员。一个进化支必须包含一个共同祖先的所有后裔,而不是其中的一部分。古人类学家更可能就分级而非分支达成一致,但决定系统树分支模式的问题必须探索,即使结果仍有争议。在随后的章节,本书将探讨一些争论的焦点。
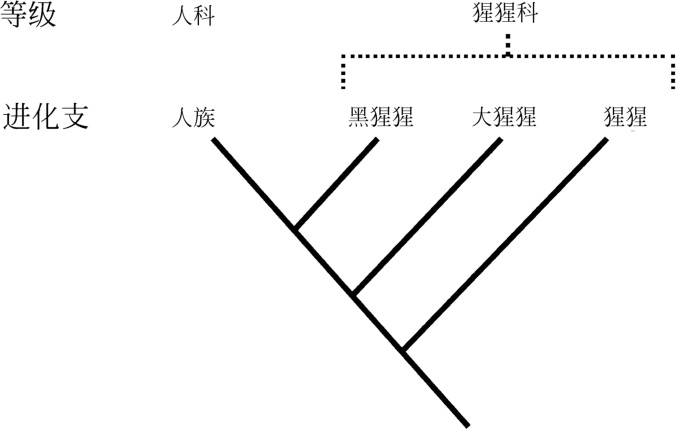
图7 现生高等灵长类分支和对应等级的比较
功能和行为形态学
除了分析化石在进化支图中的位置和种系发育关系之外,古人类学家还利用化石了解人种的适应特点。他们设法重建同一类群的个体的生活方式,然后把这些信息与生活环境证据整合并得出这一物种如何适应环境的假说。与期望了解现生动物一样,研究者也要努力更多了解灭绝动物的生活。它们吃什么?怎样活动?是群体生活,还是独自生活?古人类学家试图通过功能或行为形态学的研究来解答这些问题。
功能形态学是指观察骨骼或牙齿并推测其最擅长且最经常使用的功能。例如,经常在树上攀爬的动物的指骨就需很弯曲,因此动物的指骨弯曲就表明攀爬是其活动的一部分。指关节的形状和手指的长度也都能提供早期人类抓握物体强度的线索。要抓住锤子,需要有力的抓握。同样,使用锋利石制小工具须精确的抓握以及上臂、前臂和手部肌肉的交替配合。类似地,身体全部重量由后肢承担的动物,其大腿骨的形状就与体重由四肢共同承担的动物的完全不同。
功能形态学也有助于重建早期人类的食谱。牙齿的形状反映了所吃食物的类型。齿冠大、齿尖低而圆且覆盖有厚层釉质的牙齿,反映出饮食可能包含坚硬食物,或食物被包裹在某种坚硬外壳如坚果的壳里,需要破壳才能食用。科学家使用显微镜能够观察到肉眼无法看到的牙齿上的微痕。生长在地下的食物如植物块茎含有大量的沙砾,而这会在牙釉质的表面留下许多凹槽。有时牙齿被践踏或受到风沙中硬沙粒的撞击也会有擦痕,但这类的损伤不仅仅在牙齿的咬合面上,也会在牙齿的侧面出现。当通过食物遗留的微痕证据来找寻早期人类的饮食线索时,研究者必须确保没有把动物死亡后产生的微痕与动物存活期间留下的微痕相混淆。
人类食物种类的直接证据来自于稳定同位素分析。这种分析方法首先是测量化石骨骼或牙齿里的氧、氮和碳同位素含量,然后,把化石的食谱模型与已知的现生动物的食谱进行对比。例如,叶食动物有别于草食动物,也有别于主要食肉类动物。布拉德福德大学考古学系的同位素化学家朱莉亚和她的同事利用这种方法证明,发现于斯瓦特克朗斯的距今1.5百万年的傍人化石有稳定的同位素模型,属食肉类型。这一结果引发了研究者对早期认为其以素食为主的观点进行重新思考。
人类化石记录的偏缺
几十年来,古人类学家采集了数千件的人类化石,其时代可追溯到六至七百万年前。虽然这个数字听起来令人振奋,但大多数都集中在人类进化的后一阶段。除了时代偏缺,人类化石记录还存在其他偏向性和薄弱环节。发现并设法弥补这些偏缺的工作是埋藏学的重要课题。骨骼中一些最硬的部分如牙齿和下颌骨被发现的较多,而颅下的脊柱及四肢,尤其是脊椎、手和脚的标本都发现的较少而且保存得不好。骨骼不同部分的相对耐久性(例如,下颌骨普遍较重,且骨密度比椎骨高)可以解释为何身体各部分保存情况不同。较轻的骨头如椎骨很可能在倾盆大雨之后被洪水席卷到湖里,与鱼和鳄鱼的骨骼化石混杂在一起。相比之下,较重的骨骼如头骨和颌骨会沉落到溪流或河流底部的石头缝隙里,与其他陆生动物的重骨骼一起保存在河流的沉积物里。
造成保存状况差异的另一个因素是,许多人类的尸体成为了食肉动物的美餐。豹子喜欢咀嚼猴子的手和足,如果灭绝的大型食肉类动物有类似的偏好,那么人类身体的这些部分就会很少变成为化石。因此,我们掌握的牙齿演化知识要比掌握的手和脚演化知识多。体型大小对类群是否可能形成化石也有很大的影响。大体型类群比小体型类群更容易形成化石;同样,在一个类群中,大个体比小个体成为化石的可能性更大。有充分的理由认为,这些偏缺现象同样存在于人类化石中。
某些环境更利于形成化石,也更容易发现化石。因此我们不能说,某一特定时期或特定地方的化石多就意味着出现在那个时候或那个地方的人类个体就多。只是那个时期或那个地点的环境可能比其他时期或地点更有利于化石的形成。同样,某一特定时间或地点缺少人类化石并不能表示那时那地缺少人类。正如俗话所说,没有证据并不表示不存在证据。类似的逻辑表明,某些类群在其最早化石记录之前可能已经存在,也可能存活超过了化石记录的时代。因此,某类群化石人的最早出现记录(FAD)与最晚出现记录(LAD)可能是对这个类群起源与灭绝时间的有保留的说法。
同样的保留态度也适用于对化石地点地理分布的描述。几乎可以肯定,人类生活的地方远比发现的化石地点要多。古老的环境往往不同于我们现在看到的,现在我们认为不宜生存的一些地方在远古时期未必如此;反之亦然。
最后,并非所有的环境有利于保存骨骼和牙齿。骨骼和牙齿也很少能在强酸性的土壤环境里存留。有很长一段时间,人们认为在古森林环境中不会找到人类化石,因为那里的腐殖酸水平很高,但后来被证明是错误的。在有些地点,考古学家曾期望可以发现石制工具和骨骼一起,而他们只发现了石制工具,骨骼和牙齿在形成化石之前已被分解了。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










