



一、低温生境及微生物
在地球生物圈中,80%以上面积常年温度低于5℃。低温生境主要为深海(约占地球面积的75%)和南北两极地区,还包括高山、冰川和冻土域。此外,还有一类昼夜和季节性低温区域。人为的低温环境包括一些制冷设备和冷藏装置。
1.自然环境中的嗜冷微生物
深海水温常年低于5℃,从深海水中已分离到多种嗜冷微生物。深海海底有大量的沉积物,其深度可以达到1 000米。已经证实在海底500米以下的沉积物中有嗜冷细菌种群,并发现了独特的嗜冷细菌。由于海底的巨大压力,从这种环境中分离的嗜冷细菌多数也是嗜压或耐压菌。
南极由于其独特的地理及气候特征,孕育了丰富的低温微生物资源。已经从南极冰、雪、水、土壤及岩石样品中分离到各种类型的微生物,其中包括大量的嗜冷和耐冷微生物,如从南极土样中筛选到具有溶菌和利用琼脂的嗜冷型黏细菌,从南极湖底分离到一株无细胞壁的专性厌氧嗜冷细菌,它对青霉素具有抗性,能改变其细胞形态。此外,从湖中还分离到一株无细胞壁的螺旋状嗜冷古细菌。南极蕴藏着巨大的冰川,其中封藏着大量的微生物。通过对取自南极3 590m处的一支冰芯(冰龄约1万年)的分析证实该冰层是一有微生物活性的生态系统,从中分离出多种微生物。应该指出的是,即使是在上述常年低温的环境中,分离到的微生物大多数是耐冷菌,嗜冷菌只占很小比例。
在昼夜和季节性低温环境中,能分离到几乎所有类型的微生物。生活在这种环境下,嗜冷微生物对温度波动具有较强的耐受性,只在温度适宜的条件下才生长繁衍。
在低温环境中,革兰氏阳性细菌为优势类群。常见的有假单胞菌属、无色菌属、黄杆菌属、产碱杆菌属、噬细胞菌属、产气单胞菌属、弧菌属、锯杆菌属、埃希氏菌属、变性菌属和嗜冷杆菌属。然而革兰氏阳性细菌中的真细菌、芽孢杆菌和微球菌却比较少见。虽然许多土样和水样的16srRNA分析都显示古生菌为低温生境中的优势菌群,但是迄今分离到的菌株数量很少,主要为产甲烷和极端嗜盐的古生菌。嗜冷酵母主要是假丝酵母属和球拟酵母属的一些种类。嗜冷丝状真菌主要包括青霉属、分枝芽孢菌属、Phoma和曲霉属中的一些种类。最为熟知的嗜冷微藻是一种雪藻(Chlamydomonas nivalis),这种藻产生鲜红的孢子,从而使得其栖息地与白雪背景形成鲜明对照。
2.人为环境中的嗜冷微生物
冷藏食品可延长储存时间,同时也为嗜冷微生物的生长繁衍提供了营养丰富的生境。已经证实许多冷藏食品的变质和毒化与嗜冷菌和耐冷菌有关,如乳制品的变质与假单胞菌属、不动杆菌属、产碱菌属、产色细菌属和黄杆菌属的一些细菌有关,而肉制品的污染常与乳酸杆菌属和Brochothrix属的某些种类相关。此外,一些耐冷细菌,如梭菌属、耶森氏菌属和产气单胞菌属的某些菌株还产生毒素,可导致人畜食物中毒。
单细胞生物不能调节自身的温度,因此,嗜冷和耐冷微生物必须改变其细胞组分和结构,主要是细胞膜、蛋白质、酶分子组成和结构,以适应在低温条件下生长的需要。
3.质膜
当生长温度改变时,微生物细胞质膜最重要的应答反应是改变其磷脂的脂肪酸组成。细胞正常发挥功能要求其质膜中的脂质有较大的流动性。当温度下降时,双层脂质中的脂肪酸链的排列发生变化,从流动的无序态变为有序的结晶排列。在低温环境中,嗜冷微生物可通过改变其质膜中脂肪酸的组成,从而降低质膜从无序到有序的相变温度。这类改变涉及脂肪酸的下述一种或多种变化:①不饱和性增加;②平均链长下降;③甲基支链增加;④反式支链/顺式支链的比例上升;⑤sn-1和sn-2位置上脂酰链的异构变化。最常见的改变是饱和度和链长,甲基支链的数量和构型改变绝大多数发生于革兰氏阳性细菌。多数细菌仅含单不饱和脂肪酸,并且其含量随温度下降而增加。多不饱和脂肪酸可提高质膜的流动性。
通过去饱和酶可增加脂肪酸的不饱和性,这是改变质膜组成的一种快速方式。相比之下,脂肪酸链长和甲基支链的数量及结构改变则需要重新合成,因而是一个相对缓慢的过程。
4.蛋白质结构与酶活性
嗜冷微生物产生的酶在低温下具有很高的催化活性,这种高催化效率与其蛋白质结构的柔性相关。柔性大的酶蛋白易于产生具有催化效能的构象变化,因而使其与底物集合的活化能降低。通过比较嗜冷微生物与相应嗜温微生物的同种蛋白质序列,已鉴定出一些与嗜冷酶柔性相关的结构基础,包括盐桥和氢键减少、芳香基相互作用减弱、疏水簇减少、脯氨酸含量降低、分子表面亲水性增强等。不同的酶可以不同的方式来提高其结构的柔性。
另一方面,嗜冷微生物产生的酶一般对热不稳定,在室温条件下容易失活。酶的稳定性、活性和柔性是紧密相关的,嗜冷酶的高催化活性要求其分子具有较高的柔性,而分子柔性的增加又往往降低其稳定性。事实上,酶的稳定性、活性和柔性三者之间的关系是错综复杂的。酶的活性中心只占整个酶分子的很小部分,其中只有少数几个氨基酸残基参与底物的结合和催化作用,而酶的稳定性在很大程度上是靠其非活性中心的氨基酸序列来维持的。由此可知,在低温环境中生活的微生物有可能产生既有高催化活性又有较高热稳定性的嗜冷酶,即具有局部柔性的酶。采用定点突变已经获得一种枯草杆菌蛋白酶,它的热稳定性有明显提高,但仍保留了原有的催化特性。
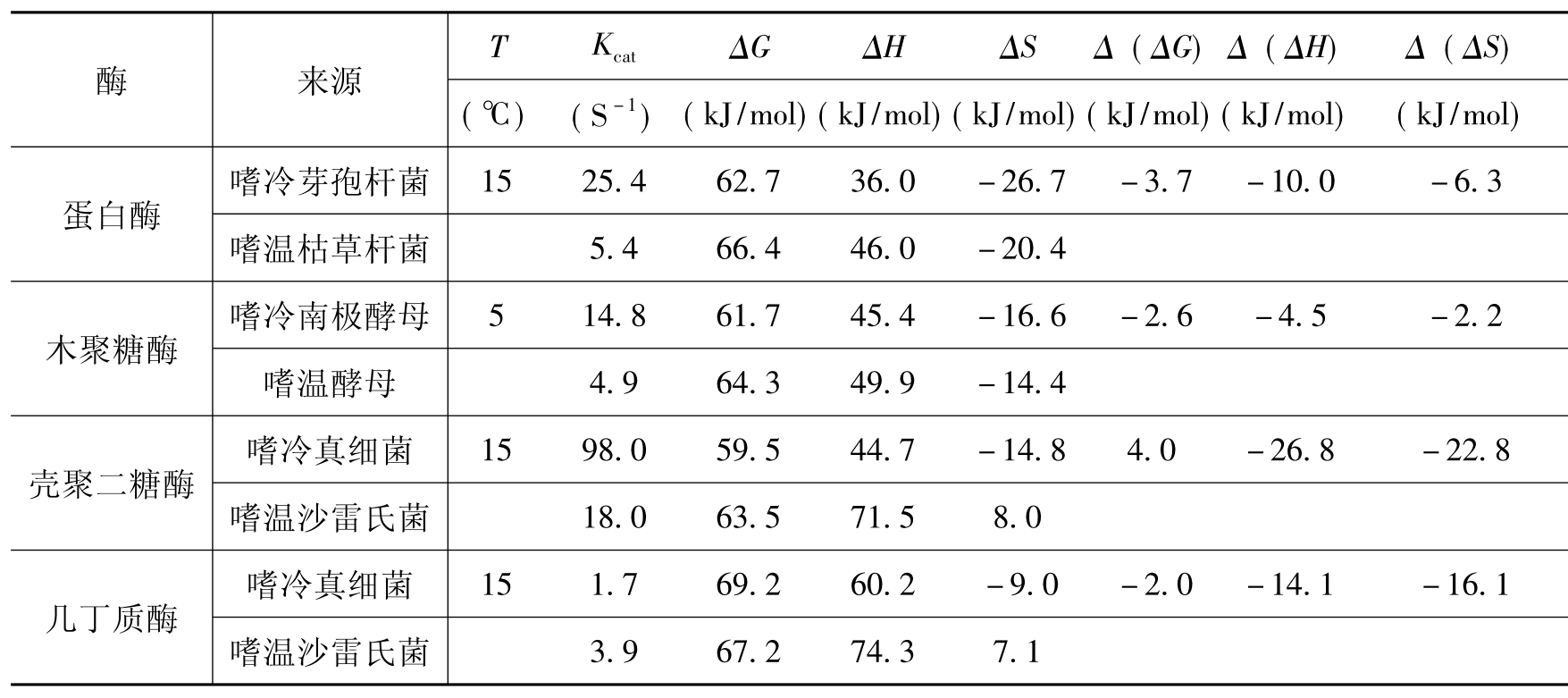
通过测定活化热力学参数可证实嗜冷酶局部柔性的构象。表2-7比较了几种嗜冷和嗜温酶活化的热力学参数,如自由能(ΔG)、焓(ΔH)和熵(ΔS),显示嗜冷酶的主要适应方式是大幅度降低其ΔH,从而获得较高的转换数(Kcat)。这种现象在低温条件下特别明显。然而,嗜冷酶的ΔS却较小,一方面,这意味系统的有序性较高,不利于酶与底物的有效碰撞;另一方面,这表明酶分子保持了某些结构域的稳定性,而这种相对稳定的构象往往对酶催化活性是必需的。
表2-7 嗜冷和嗜温酶的活化参数
5.冷休克和冷适应
当生长温度突然下降(冷休克)时,微生物细胞会诱导产生一组冷休克蛋白(cold shock proteins,CSPs)。CSPs是一组小分子量(7~8kU)的酸性蛋白,它们在微生物对低温的生理适应过程中起重要作用。在某些情况下,CSPs具有转录增强子和RNA结合蛋白的功能。在S.cerevisiae细胞中,温度休克诱导蛋白I是一种主要的CSP。该蛋白合成后被运送到质膜外周,并进行大量的D-甘露糖糖基化修饰。由此看来,在低温适应过程中,这种温度休克诱导蛋白对质膜具有保护作用。
除CSPs外,还发现了一组与嗜冷微生物在恒低温条件下生长有关的蛋白——冷适应蛋白(cold acclimation proteins,CAPs)。与CSPs相比,CAPs的一个明显特征是能在低温生长过程中持续合成。但两者没有绝对界限,有些CSPs本身就是CAPs。
一般而言,蛋白质合成对温度敏感。因此,嗜冷微生物必须有独特的冷适应核糖体和辅因子(如起始和延长因子)。此外,一些嗜冷和耐冷微生物也要经受环境温度的突然下降。冷休克应答已经在许多嗜冷和耐冷微生物中得到证实,包括Trichosporompullubans、嗜冷芽孢杆菌、Aquaspirrillumarcticum和Arthrobacter globiformis。已知冷应答反应(如诱导蛋白的种类和数量)在这些菌株中依赖于温度下降的幅度,但还不清楚其调控机理。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










