



第二部分植物病毒
植物病毒是杆状体或球形体。以杆状病毒来说,在电子显微镜制备中长度很不一致,它们很可能是正常病毒粒子的碎片或它们的聚集体。有的杆状体比较纤细而弯曲,并且还有分叉,这些应该说是线条体。表2示几种植物病毒的大小。
表2 几种植物病毒的大小

植物病毒的主要组成成分是蛋白质和核糖核酸(RNA)。这两个组成部分的比率在不同的病毒中有很大的区别(烟草花叶病毒含5.2%~6%的RNA,芜菁黄化花叶病毒含38%的RNA),但是每种病毒所含RNA的绝对量是一致的。
近几年来用化学或物理方法把完整的病毒粒子降级为亚结构。如能获得这些亚结构的某些化学的或生物学的鉴定,则对病毒的结构与功能的关系将会有进一步的了解。这方面的研究工作在几种病毒中已取得了显著的成就,烟草花叶病毒是其中之一(Hart,1956)。
近几年来有关烟草花叶病毒的重要发现具有深远意义的(Herriott,1961)是裸体的或自由的RNA有感染性(Gierer and Schramm,1956;Fraenkel-Conrat,1956)(所谓裸体的或自由的RNA指从病毒分离出来而提纯的RNA)。巳分开的RNA和蛋白质在适当的实验条件下混合后可再建成为有感染性的完整病毒(Fraenkel-Conrat and Williams,1955;Fraenkel-Conrat,1957),同时蛋白质部分也能聚集成杆状体(Schramm,1943)。从外形看,这种蛋白质聚集的杆状体和完整病毒粒子相似,几乎不能区别,但由于缺乏RNA,因而无感染性。

图60 从粒子中轴的平面切成一半的一短段烟草花叶病毒的结构示意图,显示蛋白质亚单位的螺旋形排列(螺旋体每三转为49个亚单位),螺旋槽及其螺旋顶端延长于粒子的平均半径之外和空的中轴核心(半径为207A)。在半径为407A的螺旋隧道中隐藏着核酸(RNA)(Franklin,Klug and Holmes,1957)
植物病毒是杆状体或球状体,两者都有RNA中髓,外由蛋白质所包围,而蛋白质则由相同的多肽链(亚单位)分子所组成。

图61 烟草花叶病毒的结构示意图,显示没有蛋白质外壳的一部分核酸链(当然在实际情况下,没有蛋白质,核酸是不可能维持它的构型的)。每个盘状体代表一个蛋白质亚单位(Klug and Caspar,1960)
关于植物病毒的感染过程现在的知识还是很不够。虽然实验已提供证据,RNA是植物病毒的遗传性物质(Fraenkel-Conrat,1956),但进入宿主细胞的遗传性物质是否只有RNA或尚有其他物质还不了解。在植物病毒感染过程中值得注意的是在病毒感染植物中的异常蛋白质问题。感染植物中除病毒外还存在着异常蛋白质。所谓异常蛋白质是指具有和病毒有关的抗原性蛋白质,而它们又不存在于健康植物中。对这一类蛋白质的知识比较多的是烟草花叶病毒感染的植物。其他病毒感染的植物中也有类似的物质产生[烟草坏死病毒(Rothamsted株)、芜菁黄化花叶病毒、甜菜黄化病毒等]。
1945年在烟草花叶病毒感染的植物叶部发现了含有血清学特异性相同的而沉淀率有差别的各种组成部分(Bawden and Pirie,1945),其中较小的粒子缺乏感染性。在芜菁黄化花叶病毒感染的植物中同样获得了两种分子量和密度不同的粒子(Markham,1953)。这两种粒子的大小、形状以及它们的结晶体外表和结构完全相同,主要区别在于一种含有38%的RNA,并且有感染性,另一种没有RNA,又无感染性。后者类似于噬菌体的“幻象”或空壳体。
近年来对烟草花叶病毒感染的植物研究证实了早期的观察(Bawden and Pirie,1945),发现在感染植物中含有沉淀率很低的蛋白质组成部分(Jeener,Lemoine and Lavand'home,1954,称它们为可溶性抗原)。这些蛋白质和病毒在血清学上有关,如Taka-hashi氏的X蛋白质(Takahashi and Ishii,1952)、Commoner氏的所谓非病毒蛋白质“B3、B6”和“A4”等(Commoner,1952,1953,1954)。它们共同的性质是不含有RNA和缺乏感染性,以适当的化学方法处理(以硫酸铵沉淀或控制pH值下降),它们能聚集成为较大分子,再进一步聚集能形成和病毒粒子类似的蛋白质杆状体(Rich,Dunitz and Newmark,1955),并且把pH值适当地下降能促使它们以酝晶形态完全沉淀下去。这种酝晶体(Para-crystal)在形状上和病毒的酝晶体没有任何区别(Jeener and Lemoine'home,1954)。
异常蛋白质在感染植物中出现的时间,根据各学者的报告有所不同(Rich,Dunitz and Newmark,1955;Franklin and Commo-ner,1955;Newmark and Franklin,1956),但它们一般都出现于烟草花叶病毒粒子出现之前,并且它们在形状上、血清学反应上、X射线结晶的样式上以及氨基酸的组成上都与病毒粒子相类似,因此,它们是病毒在宿主植物中合成过程中的中间物,也就是病毒的先质,那就是说,在病毒合成前首先合成这些蛋白质,然后包围RNA髓核聚集为完整的病毒粒子。
植物病毒,杆状的或球形的,在一定条件下都能形成结晶体。事实上,早在1904年Iwanowski氏研究烟草花叶病时,制备花叶病叶的染色切片中就注意到加入一种酸性固定剂后在叶细胞内形成一种“条纹物质”(Stanley,1941)。现在看起来,这所谓条纹物质就是烟草花叶病毒的结晶体。杆状体病毒一般产生酝晶形态,(二次元或平面体)缺乏有规律的立体网状。烟草花叶病毒的酝晶体是病毒第一次获得的结晶体(Stanley,1935),现在植物病毒的结晶体巳获得10余种之多。
一、各种植物病毒

图62 一个烟草花叶病毒粒子和粒子的一小碎片(Williams,1953)

a.普通摄影,没有投影;b.金属投影;c.聚集的X-蛋白质(Williams,1954)
图63 烟草花叶病毒(×150000)

图64 马铃薯X病毒(Potato X virus)(Harrison,1959)

图65 马铃薯Y病毒(Potato Y virus)(Schramm,1954)

图66 烟草激响病毒(Tobacco rattle virus)(Harrison,1959)

图67 天仙子花叶病毒(Heubane mosaic virus)(Harrison,1959)
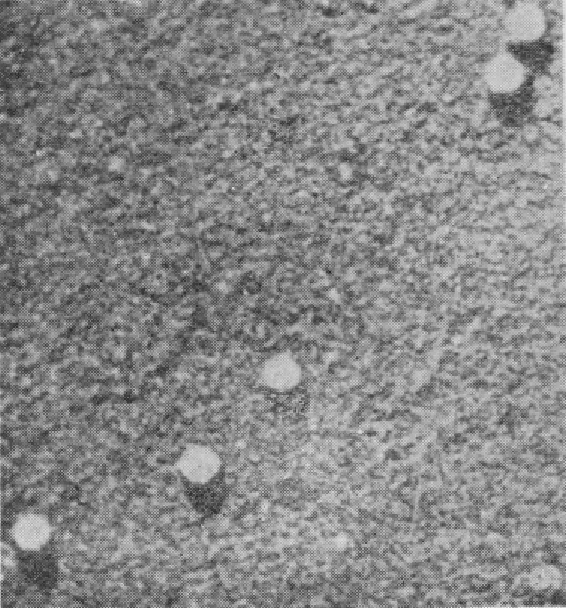
图68 番茄丛矮病毒(Tomato bushy stunt virus)(Williams,1953)

图69 烟草环斑病毒(Tobacco ring spot virus)(Williams,1953)

图70 马铃薯黄矮病毒(Potato yellow dwarf virus)(Harrison,1959)

图71 番茄斑萎病毒(Tomato spotted wilt virus)(Harrison,1959)

图72 苜蓿伤痍癌病毒(Clover wound tumor virus)(Harrison,1959)

图73 南方菜豆花叶病毒(Southern bean mosaic virus)(高尚荫,未发表)。

图74 番茄丛矮病毒(Tomato bushy stunt virus)(左)(×150000)(右)(×100000)(Williams,1954)

图75 芜菁黄化花叶病毒(Turnip yellow mosaic virus)(×500000)(Markham,1959)
二、植物病毒的结构

图76 烟草花叶病毒的结构(×125000)。(左)完整的病毒杆状体;(中)局部降级的粒子,显示RNA;(右)没有RNA的粒子横切面,注意中间的小孔(Stanley,Lauffer and Williams,1959)

图77 烟草花叶病毒用酚处理后蛋白质部分降解的各种不同程度的状态。丝状体是RNA(Corbett,1964)。

图78 烟草花叶病毒(×150000)。用碱作短时间的处理后,一部分蛋白质从病毒杆状体脱去。联系四段杆状体的细丝是暴露出来的RNA(Schramm,1955)

图79 重聚集的烟草花叶病毒蛋白质(Nixon and Woods,1960)。(上)显示中轴小孔和循环节约20~257A的纵面条纹,这等于烟草花叶病毒螺旋体螺距237A(×270000);(下左)垂直状立着的杆状体的一短段。围绕圆圈的隆起状体的数目是约16。从X射线分析所推测的螺旋体每转的亚单位数是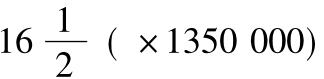 ;
;
(下右)上图的放大,显示亚单位(×1350000)

图80 烟草花叶病毒的X-蛋白质,垂直状堆积圆盘构型,注意由16个亚基建成(Markham,Frey and Hills,1963)

图81 烟草花叶病毒粒子的堆积圆盘构型(Markham,Hitchborn,Hills and Frey,1964)

图82 没有核酸的烟草花叶病毒蛋白质亚单位在不同pH值的溶液中。(上)pH10.0;(中)pH6.0,和正常的病毒蛋白质相类似;(下)pH5.2,接近等电点,杆状体聚集(Schramm and Zilling,1955)

图83 (上)烟草花叶病毒;(下)重聚集的烟草花叶病毒(Gierer,1960)

图84 在不同pH值中的烟草花叶病毒。(上)pH5.2,接近烟草花叶病毒的等电点,杆状体聚集成较长的杆状体;(中)pH8.6;(下)pH10.0(Schramm,1954)

图85 (上)感染着烟草花叶病毒的烟草中的异常蛋白质;(下)正常的烟草花叶病毒(Commoner,1953)

图86 甜菜黄化病毒(Beet yellow virus)。a.b.丝状体病毒显示周期性结构;c.丝状体,中心区域显示空的中心。丝状体的成形圈表示其柔软性(Horne,Russel and Trim,1959)

图87 烟草花叶病毒在等电点pH3.5,显示正常病毒杆状体聚集成丝状体。注意右角上的一个正常病毒杆状体(300×15)(高尚荫,未发表)
三、植物病毒的结晶体

图88 含有烟草花叶病毒结晶体的烟草植物细胞(左)冻结干燥前的标本;(中)冻结干燥后的标本;(右)从细胞内抽出的一个结晶体(Luria,1953)

图89 上图的一个结晶体以水溶解后释放出烟草花叶病毒粒子(Luria,1953)。

图90 烟草坏死病毒整块结晶体(Labaw and Wyckoff,1956)

图91 同上(Hercik,1959)

图92 在感染着烟草花叶病毒的烟草毛细胞内的两个病毒结晶体(Bawden and Sheffield,1939)
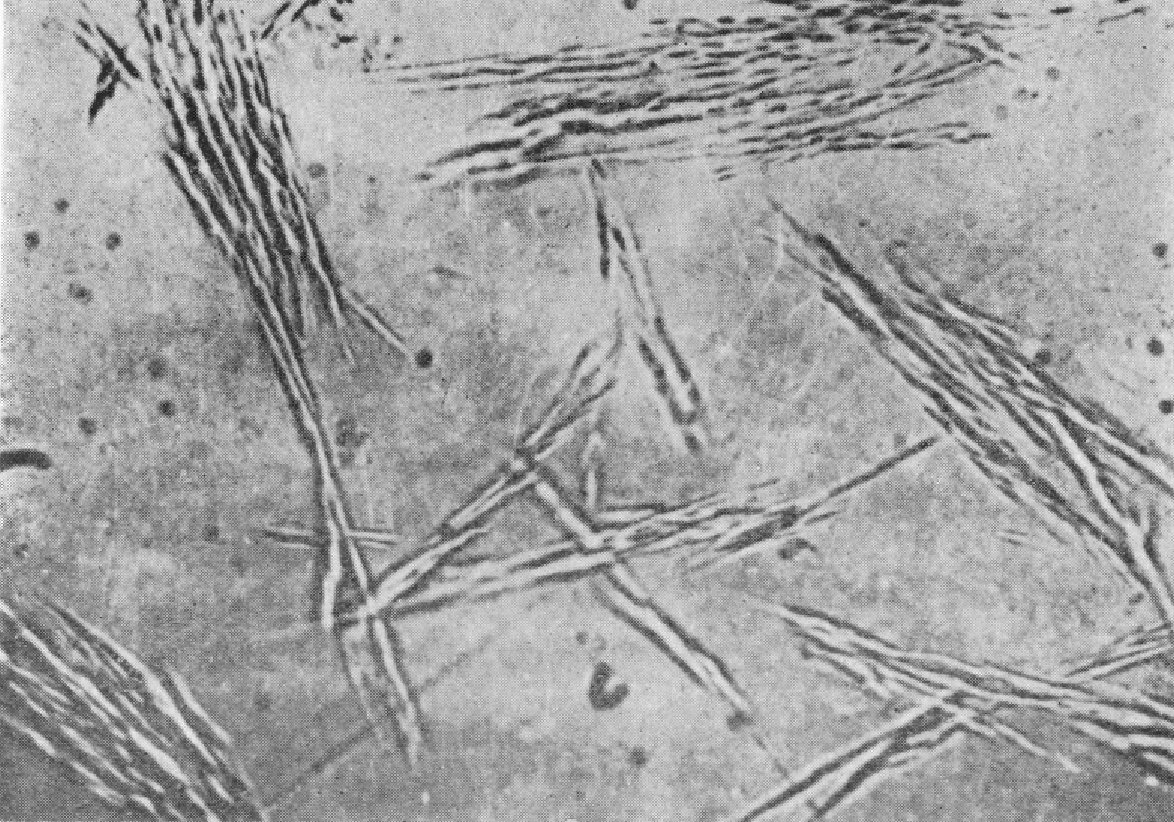
图93 烟草花叶病毒结晶体(Schramm,1954)

图94 芜菁黄化花叶病毒结晶体(Smith,1950)

图95 同上。(右)完整病毒的结晶体;(左)没有RNA的蛋白质结晶体(Smith,1950)

图96 在血清反应上有关系的病毒株形成不同状态的结晶体,如烟草坏死病毒。(上)六角形柱状体结晶;(下)薄的菱形片结晶(Bawden,1943)

图97 番茄丛矮病毒结晶体(Bawden and Pirie,1938)

图98 南方菜豆花叶病毒结晶体,各种菱形板和柱状体的变形(Miller and Price,1946)

图99 烟草坏死病毒结晶体(Smith,1950)
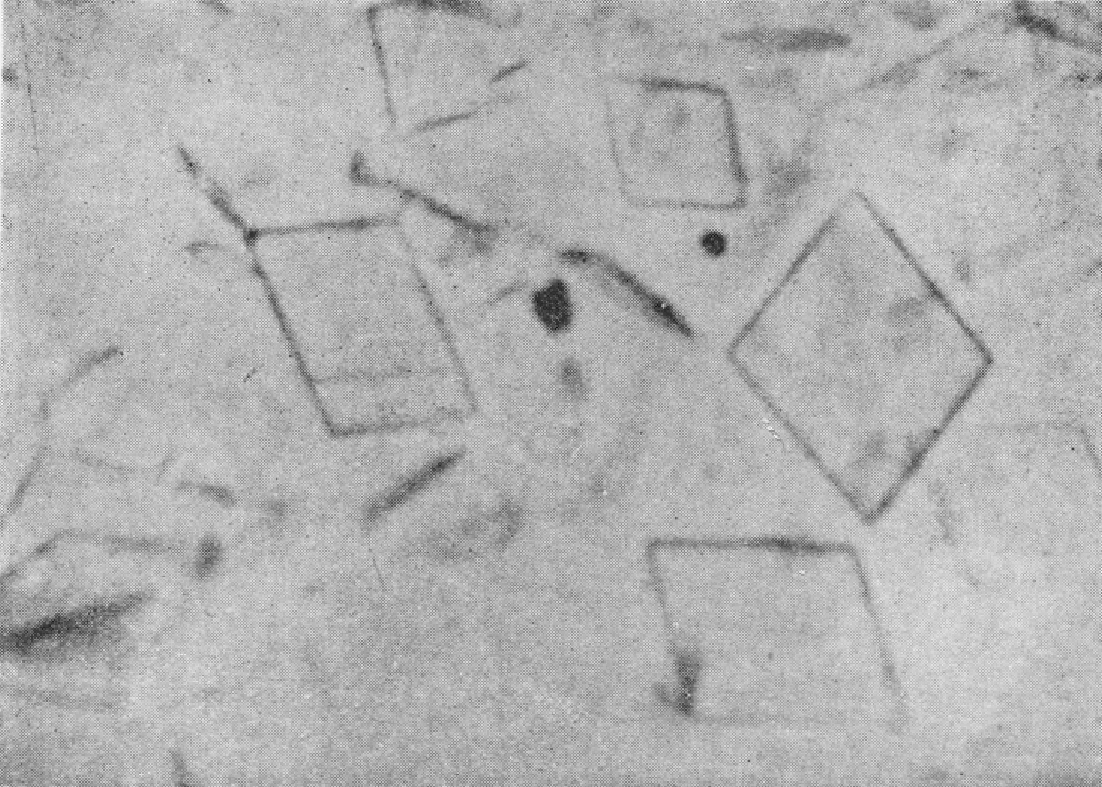
图100 南方菜豆花叶病毒结晶体(Price,1946)
四、其他

图101 烟草花叶病毒,用超声波处理后正常的杆状体断裂成为长度不等的碎片(高尚荫,未发表)

图102 超声波处理的烟草花叶病毒(Pollard,1953)

图103 烟草花叶病毒和其抗血清混合后形成稻草状的凝集。注意粒子平行排列间似有距离,这距离间可能是抗体分子所占的位置(高尚荫,未发表)
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











