



(一)树种调查
在岩质海岸树种调查方面,王有方(1995)对浙江台州市海岛植被资源及其利用进行了研究。该市有海岛678个,其中长期住人岛36个、季节性住人岛42个、无人岛607个,总面积275.44 。通过调查,基本掌握了海岛植物区系的特征及种类组成,共有维管植物1210种,分别属170个科650属,其中蕨类植物25科38属54种;野生裸子植物1种,栽培裸子植物7科21属42种;野生被子植物112科485属936种,栽培被子植物25科105属177种。小科、极小科比例高,小属、极小属成分丰富,说明海岛植物区系成分的多样性和复杂性。对486个野生种子植物属的分布区类型进行统计分析,结果表明海岛植物区系地理成分复杂,与世界各地各种区系成分有广泛的联系,同时也说明该岛具有较典型的中亚热带植物区系性质,且略偏向南亚热带区系的趋势。
。通过调查,基本掌握了海岛植物区系的特征及种类组成,共有维管植物1210种,分别属170个科650属,其中蕨类植物25科38属54种;野生裸子植物1种,栽培裸子植物7科21属42种;野生被子植物112科485属936种,栽培被子植物25科105属177种。小科、极小科比例高,小属、极小属成分丰富,说明海岛植物区系成分的多样性和复杂性。对486个野生种子植物属的分布区类型进行统计分析,结果表明海岛植物区系地理成分复杂,与世界各地各种区系成分有广泛的联系,同时也说明该岛具有较典型的中亚热带植物区系性质,且略偏向南亚热带区系的趋势。
高智慧等(1994)进行了浙江省舟山群岛几种主要树种的生长调查,结果表明香樟、湿地松、柏木3个树种在该地区生长良好,胸径、树高、蓄积的年生长量均大于马尾松及黑松,林分生产力较高,可与杉木等速生丰产林的生长相媲美。同时,这3个树种的胸径生长呈正态分布,为稳定的群落结构。因此,在岩质海岸发展这3个树种有很大的潜力,可作为替代马尾松及黑松在茎岩质海岸造林的优良树种。
唐正良等(1996)针对园林栽培中普遍偏重外来植物、绿化效果不良、难以体现海岛特色的问题,在植物资源调查基础上,筛选出了浙江海岛观赏植物1149种,根据植物习性将其划分为十大类别,即:一二年生花卉(153种)、宿根花卉(295种)、球根花卉(35种)、兰科花卉(20种)、观赏蕨类(98种)、水生花卉(50种)、观赏乔木(126种)、观赏灌木(216种)、观赏竹类(35种)和蔓木类(121种),并分别阐述了资源现状、观赏特性和园林用途,提出了开发利用意见。在调查中还发现,海岛观赏植物具有明显的水平分布规律,在温岭交陈以北属华东植物区系,植被丰富且保存相对较好;其南部属华南植物区系,资源数量不及北部岛屿。另外,就土壤而言,红壤区域是观赏植物最富集的区域,其次为粗骨土,其他土类上较贫乏,但滨海盐土上分布着一些专性植物。
“六五”和“七五”期间,在国家科委牵头组织下,由国家科委、国家计委、国家农委、总参谋部和国家海洋局等单位组成全国海岸带和海涂资源综合调查领导小组,对全国海岸带和海涂资源进行了综合调查,内容涉及气候、水文、土壤、植被、林业等12个专业几十个项目。调查结果显示,我国岩质海岸防护林树种主要有黑松、樟子松、马尾松、油松、火炬松、刚松、台湾相思、麻栎、蒙古栎、槲栎、苦槠、石栎、白栎、红椎、丝栗栲、青冈、侧柏、柏木、木荷、窿缘桉、朴树、糙叶树、黄檀、蚊母树、枫香、响叶杨、桤木、灯台树、冬青、山槐、化香、毛竹、刚竹、淡竹、苦竹等。
王礼先等(2000)也提出了我国岩质海岸各岸段主要造林树种,如在辽东、山东半岛沙岸间岩岸丘陵山地上,主要造林树种有黑松、刺槐、白榆、麻栎、栓皮栎、油松、侧柏、辽东栎、紫椴、枫杨、赤松、落叶松、旱柳、毛白杨、沙兰杨、北京杨、加杨、楸树、香椿、臭椿、国槐、板栗、核桃、苹果、山楂、紫穗槐、胡枝子、黄栌等;在浙南、闽北岩质海岸山地丘陵区,主要造林树种有:木麻黄、落羽杉、池杉、水杉、香椿、臭椿、朴树、檫木、喜树、樟树、苦楝、鹅掌楸、刺槐、泡桐、白玉兰、乌桕、黑松、柏木、柳杉、杉木、板栗、银杏、竹类、文旦、柑橘、桂花、麻栎、栓皮栎、马尾松、枫香、木荷、黑荆树、米槠、苦槠、青冈栎、红楠、闽楠、丝栗栲、格氏栲、胡枝子、山苍子、杨梅及杂灌木类等。
陈胜(2005)研究了沙质地沿海防护林优良适生树种木麻黄、湿地松和厚荚相思的生长特性及生态适应性。结果表明,木麻黄目前仍然是沿海防护林海岸前沿不可替代的造林树种,湿地松抗风能力最强,而厚荚相思最能适应沿海沙质地干旱瘠薄的恶劣立地条件,且生长迅速,对土壤改良效果良好。韩玉洁等(2010)以上海市主要沿海防护林树种为对象,调查了2年生林分32个主要造林树种的适应性,建立涵盖生长适应性、抗逆性、景观价值等多因子的综合评价指标,采用综合评价法,对该区域防护林树种进行了评价筛选。结果表明:试验树种分为4类,表现良好的树种4种,表现尚好的树种5种,表现一般的树种5种,不适宜树种18种。在水盐协同胁迫逆境下,推荐黄连木、侧柏、乌桕、中山杉、弗吉尼亚栎、墨西哥落羽杉、红叶椿、美国皂荚8个乔木树种,胡颓子和海滨木槿2个灌木树种。王洪、张金池等(2010)调查了苏北泥质海岸主要防护林树种生长特性,结果表明:杨树生长速生期为6~11a,早于水杉的16~21a,银杏在0~10a内生长缓慢。宽冠型树种树冠在成熟龄后,仍有缓慢生长,而窄冠型树种则保持稳定。株行距越近于正方形越有利于宽冠型树种的生长,在胸径、生物量生长速度方面,林农复合明显快于片林。
(二)树种选择
为了提高沿海防护林的稳定性和生物多样性,改变沿海造林树种单一的现状,国内外学者在林分改造和树种选择方面进行了深入研究。
蒋妙定、高智慧等(1995)对浙江岩质海岸主要造林树种进行了调查研究,通过对树种分布、生长过程、生长量的分析,认为湿地松、木麻黄、桉树、香樟、枫香、柏木6个树种生长良好,林分生产力大,可逐步代替黑松和马尾松。在该海岸区发展湿地松潜力很大,立地条件较好的局部地段,可栽培杉木,培育中、小径材,还可发展经济林如杨梅、柿子、板栗、枇杷、竹子等,以形成生态经济型防护林体系,既能控制水土流失、提高水源涵养能力,又能创造一个良好的生态环境。另外,康志雄等(1995)通过对浙江省岩质海岸生态型经济林样地调查及资料收集,应用生态经济学原理,系统研究了生态型经济林的设计原理、方法及树种选择等,推荐杨梅、竹类、柚类、柿等作为亚热带岩质海岸生态型经济林的营造树种。这些调查研究工作,为沿海防护林体系建设提供了科学依据。
要正确选择造林树种,做到因地制宜、适地适树,就必须了解树木的生态特性。据研究,在我国沿海地区耐干旱而不耐水湿的树种有马尾松、黑松、枫香、柏木、夹竹桃、山槐、合欢、柃木、化香等;较耐干旱又较耐水湿的树种有乌桕、木麻黄、苦楝、棕榈、旱柳等。耐瘠薄的树种有马尾松、黑松、相思树、柃木、化香、胡颓子等;尚能耐瘠薄的树种有木麻黄、麻栎、黑荆树、刺槐、桑树、枫杨、枫香、木荷、侧柏、女贞、湿地松、火炬松等。适于酸性土壤生长的树种有杉木、柳杉、马尾松、金钱松、油茶、木荷、毛竹、化香、木荷、铃木等;适于微酸性至微碱性土壤的树种有麻栎、枫香、侧柏、枫杨、泡桐、桉树、黑荆树、板栗、香椿、棕榈、油桐、漆树、悬铃木、枣树、柑橘、枇杷、水杉、池杉、水松、落羽杉、柿树等。这些树种都可以在我国岩质海岸选用。
沿海沙质海岸立地条件差,从50年代起木麻黄引种获得成功,而后迅速得到推广,很快形成了一条绿色长城。由于树种单一,病虫危害突出,防护效能逐渐减弱。近十几年来,有些地区采用湿地松来更新改造第一代木麻黄防护林。徐俊森(1995)通过调查研究,发现海岸带沙土上营造的湿地松普遍生长良好,不仅从立木蓄积量上比同等条件的木麻黄高出20%~40%,而且有较强的抗风、抗病虫害、耐旱以及耐瘠薄等特性,是海岸带造林较为理性的优良树种。陈建星(2000)进行木麻黄与榕树混交作沿海防护林带的试验,研究结果表明,榕树适应性强,胸径生长比木麻黄快89.5%,盖度大135%,平均寿命长10倍,达到改变沿海防护林带树种结构单一,提高林带的防护效能的目的。林武星等(2001)在福建东山县海岸后沿沙地引种厚荚相思、马占相思、大叶相思、刚果12号桉、肯氏相思、木麻黄、纹荚相思、毛绢相思、湿地松等树种在木麻黄基干林带下套种更新、风口沙地造林和带状采伐更新试验。研究结果表明,在海岸前沿风口沙地厚荚相思造林成活率高,受风害轻微,对沙荒风口不良生境适应力强;在海岸木麻黄基干林带更新中,厚荚相思保存率高,生长迅速;在林带后沿沙地厚荚相思成活率和保存率高于其他相思类树种,树高、直径及材积生长量最大,降低风速明显,林内凋落物多,提高土壤肥力显著。
李绍忠(1995)进行了辽宁省泥质海岸带防护林树种选择的研究,认为在辽河三角洲土壤含盐量0.7%~1.1%的重盐碱地上,理想的造林树种是柽柳,在干渠轻盐碱地粉沙壤土上,可栽植群众杨、新疆杨、三北1号杨、中林46杨,在水肥充足土壤上可栽植辽宁杨(Populusx Liao ningnsis)。WILLIAM H.等(2005)也对美国南部沿海抗盐碱树种baldcypress (Taxodium distichum)进行了种源试验,筛选出最优树种。
郗金标等(2004)分析了限制沿海防护林树种选择的5个方面的障碍因素,阐述了沿海防护林的树种选择原则,分别就山东沿海防护林建设中可选的乔木、灌木、草本植物资源作了介绍,并指出了提高沿海防护林造林成活率的2种造林方法:容器苗造林和根际覆盖技术。孔强、陈秋夏等(2011)对不同立地条件下东方杉等36个沿海防护林树种的生长生态特性进行了研究,结果表明,几个高抗性树种的高生长速率排序为:邓恩桉(1.18cm/a)>木麻黄501(1.16cm/a)>东方杉(1.08cm/a)>无柄小叶榕(1.07cm/a)>乌桕(0.92cm/a);径生长速率排序为:无柄小叶榕(1.58cm/a)>邓恩桉(1.35cm/a)>木麻黄501(1.15cm/a)>东方杉(1.06cm/a)>乌桕(0.93cm/a)。
沿海与内陆地区相比,温度、光照等条件差异不十分明显,而以风力的差异最大,因此沿海荒岛、荒滩、荒涂造林绿化,必须选择抗风力强的树种为先锋树种。据研究,以下几个方面可作为选择抗风树种的依据:①一般常绿树种抗风能力比落叶树种强,因为大多数树种落叶期恰好与大风干旱期吻合,常绿树种冬季仍正常生长,枝和顶芽又有叶片保护,不易出现枯芽、枯枝和枯梢,如黑松、女贞、香樟、木麻黄、桉树等常绿树种极少发现冬季枯梢现象;②枝梢粗壮与冬芽饱满的树种,抗风力较强,如马褂木、海南石梓、悬铃木等冬季基本上不出现枯梢;③根系发达、材质坚韧的树种,其抗风力比根系浅、材质松软的树种强。
(三)引种驯化
为积极引进优良树种,在我国基岩质海岸还开展了树木引种驯化工作。山东青岛于1974~1988年,先后3次从国外引进火炬松进行引种试验,通过种子育苗、田间造林等试验,测得19年生火炬松林分树高、胸径和材积年平均生长量分别为同龄日本黑松的177.8%、196.7%和829.4%,且未发现松干蚧。表明火炬松在青岛引种是成功的,但在种源上应侧重引入北亚热带和抗寒种源。浙江台州的木麻黄种源试验表明,不但不同气候带种源苗期生长有显著差异,而且即使同一个县,丘陵山地和海涂盐碱地的种源,其苗期生长也有很大差异,后者的生长量和耐寒性大大超过前者。接着把海涂盐碱地的种源北移推广,在浙江中部沿海取得了成功。浙江沿海的大鹿山岛自然条件恶劣,即使玉环县适应性最强的乡土树种马尾松也不能适应。后来总结经验,对引进树种从苗期开始就让其经受大风、干旱等恶劣环境的锻炼,增强了树种的适应性,引进的200多个树种有70%引种驯化取得成功,在岛上发育良好。陈艳珍等从1970年开始在沿海河滩地上引种日本黑松,并进行了持续20多年的育苗、移栽、造林试验。结果表明,日本黑松在造林成活率、年生长量和适应沿海地理环境方面均相当或优于油松和樟子松,适宜在沿海地区大面积引种造林。
李云等(1996)指出,我国红树林位于东亚红树林分布区的北缘,天然分布于海南岛最南端的榆林港(约18°09′N)至福建省的福鼎(27°20′N)的沿海滩涂上。由于受气候因子的影响,特别是气温和水温下降的影响,随着纬度的升高,红树植物的种类和林分高度逐渐减少和下降。为了绿化沿海滩涂,增加沿海防护林第一道防线的种质资源,提高林分质量和防风防浪效能,沿海人民及有关部门对红树林引种驯化进行了尝试和实践。1957年,浙江省瑞安市从福建引种秋茄获得成功;进入20世纪80年代后,我国政府先后在海南、广东、广西和福建沿海成立了7个红树林自然保护区;20世纪90年代以来,红树林引种驯化研究被列入国家科技攻关和支撑项目。
福建东山赤山国有防护林场自1987年起,引进5个相思类树种:大叶相思(A.auriculaeformis)、毛绢相思(A.holosericea)、肯氏相思(A.cunninghamii)、马占相思(A.mangium)、厚荚相思(A.crassicarpa)试种于滨海沙地,观察研究其适应能力与生长效果。陈胜等(1997)通过10a观测研究认为,相思树普遍能够适应滨海风沙地的恶劣条件,生长较好;厚荚相思生长迅速,干形较通直,高生长优势明显,可望成为继木麻黄、湿地松之后,营造海防林以及低效林更新改造的又一优良树种。在滨海沙地纹荚相思小面积引种试验结果表明:纹荚相思能够适应海滨沙地的恶劣条件,早期速生性表现特别突出,既有良好的防风固沙作用,又有可观的木材产量,是营造沿海防护林的优良阔叶树种。
赤桉原产澳大利亚,是目前世界上引种栽培广泛的桉树之一,我国南方各省都已在低丘、台地引种赤桉。周毅等(1994)在沿海前沿围垦草甸盐渍沼泽土上营造赤桉防护林,探究在沿海前沿赤桉防护林幼林的生长表现。
陈玉军、廖宝文等(2003)将速生红树植物无瓣海桑成功向北引种到粤东沿海和福建九龙江口,其生长表现良好,分别向北推移了约3.5个和4.5个纬度带,越过3~4个冬季后能够正常生长发育。董必慧(2005)进行了江苏沿海湿地美国白蜡树引种造林。林武星、叶功富(2009)论述了福建省沿海防护林树种现状和问题、台湾防护林建设状况和主要树种特性、引种台湾海岸树种的必要性,提出了台湾海岸树种在福建沿海防护林中的应用技术。
我国东南沿海属海洋性季风气候区,光照充足,雨量充沛,总的说来是适宜森林生长的,然而海岸带尤其是岩质海岸的立地条件也有其特殊性,并且往往存在着某些限制森林生长的因素。因此,为做到因地制宜、适地适树,必须掌握海岸带森林生长条件的特殊性,对沿海造林地进行立地类型划分和评价。
林文棣(1988)认为,我国海岸带森林的立地分类应在海岸的自然地理背景、地域分异特点以及自然区划理论的基础上,找出影响海岸立地各种不同尺度的地域分异规律,从综合生态因素中提出对立地条件起主导作用的因子,采取逐级区分的方法,将林地的立地差异足以影响到树种选择和造林技术发生改变的界线区分开来,而将其基本一致的地段归在一起,按带(亚带)、区、级、组、型5个等级,对海岸造林地进行立地类型的逐级分类。其中,带(亚带)和区属于大尺度的区划,级、组和型属中、小尺度的区划。
我国海岸造林地立地带按纬向热量的差异,可分为南温带、北亚热带、中亚热带、南亚热带、北热带、中热带和南热带7个立地类型带。同一立地类型带中又可根据水湿条件差异、气候干燥度的不同,将7个立地类型带划分出12个亚带。在立地类型带(亚带)内,可按海岸动力地貌类型即岸段作进一步划分,岸段就成为第二级立地分类单位,称立地类型区。我国沿海地区共可划分出55个立地区。在同一个岸段之内,中地形地貌单元如山地、丘陵、台地、阶地、冲洪积平原、河口三角洲、海积平原(含泻湖平原)等,控制着林地的生态环境,也影响到造林树种选择和造林技术,可用来划分第三级立地类型,称立地级。在同一个中地形地貌单元之下,对海岸造林树种选择和造林技术起主要限制作用的因素,例如在低山丘陵区,以基岩和母质的性质、土层厚度、石砾含量、地形、气候等为主要限制因子,就成为划分第四级立地类型即立地组的依据。在同一立地组之内,林地肥力等级的差别,是划分第五级立地类型的依据。这样,就形成了我国海岸造林地的立地分类系统。
王德安等(1996)通过对青岛市岩质海岸宜林地立地条件和林木生长的分析,选择中地貌、坡位、坡向、土壤厚度和石砾含量作为划分立地类型的主要因子,并结合实践经验,将该海岸段划分出2个立地类型区、7个立地类型组、24个立地类型。在浙江岩质海岸,高智慧等(1997)分析了树木生长与立地因子的关系,认为大地貌(不同类型的岩质海岸)、坡位、土层厚度和石砾含量是影响树木生长的主导因子,将浙江岩质海岸宜林地划分为3个立地类型区、9个立地类型组、30个立地类型,为适地适树造林提供了科学依据。在江苏沿海地区,胡海波、张金池等(1991)分别对海岸造林地立地类型进行过划分,并对其作了分析评价,这些都为沿海防护林体系建设提供了可靠依据,使林业生态工程建立在切实可行的基础上。陶吉兴、余国信(1994)将浙江省“沿海立地区”划分成4个立地类型区、7个立地类型组和26个立地类型。
(一)生理生态特性
环境条件如气候、土壤等对林木生长有很大影响。Dale V.H.等研究认为,气候变化对美国太平洋西北沿岸花旗松林的生长有一定影响。研究人员用计算机模拟规划了两种气候条件下的林分生长方案:常见气候条件和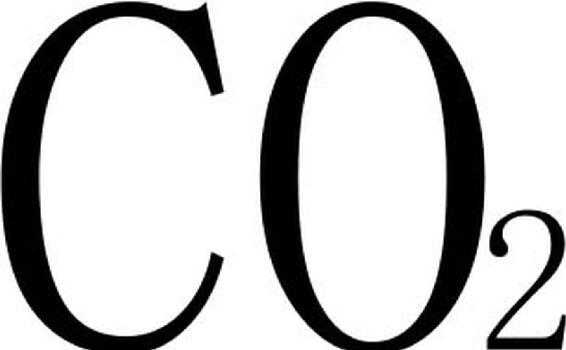 浓度加倍的温暖气候。该模型估计,气温会引起树种组成、叶面积和林分密度的变化,但地上生物量无变化。Peterson C.E.等在该地的研究结果是,花旗松天然林分的平均材积和增长率与夏季高温呈负相关,而与非生长季的高温呈正相关。因此,冷湿的夏季和温暖的冬季可使该地针叶林具有较高的生产力。
浓度加倍的温暖气候。该模型估计,气温会引起树种组成、叶面积和林分密度的变化,但地上生物量无变化。Peterson C.E.等在该地的研究结果是,花旗松天然林分的平均材积和增长率与夏季高温呈负相关,而与非生长季的高温呈正相关。因此,冷湿的夏季和温暖的冬季可使该地针叶林具有较高的生产力。
陈顺伟等(2001)研究了杜英等11个树种对盐雾胁迫的反应及其生理特性。通过模拟盐雾胁迫观察及结合叶片叶绿素含量、游离脯氨酸含量、SOD活性、MDA含量等生理指标的测定,指出不同组织耐盐雾性强弱分别为成熟叶>嫩叶>芽,7个参试树种的耐盐雾性强弱综合排序为杨梅、木荷、杜英、二次结实板栗、湿地松、枫香、马尾松。
苏祖荣(1999)进行了沿海防护林木麻黄的抗盐能力及滨海盐碱地的降盐技术研究,结果表明,1年生木麻黄在土壤含盐量为0.6%时,造林成活率可达60%以上;最大抗盐量为0.767%,木麻黄成林最高抗盐量为3.141%;抗盐锻炼等因素可提高木麻黄的抗盐能力;大畦深沟、开沟起埝、高堆法等整地方式可使含盐量下降50%~68.7%。
冯泽幸等(1996)研究比较了几个树种对模拟海风的抗性,结果表明,木麻黄和夹竹桃为高抗树种,粗枝木麻黄、细枝木麻黄和湿地松属中抗树种,山地木麻黄抗性较差。游离脯氨酸与抗“海风”力的强弱无明显规律。陈少雄等(1995)开展了抗风育种、抗风栽培措施的研究,对不同株行距配置的桉树幼林抵抗台风的能力进行了检验,证明不同密度、不同龄级的桉树在遭受风害的等级上有明显差别。李国旗等(1999)在对实验数据分析的基础上,计算和比较了杨树、水杉、刺槐和泡桐的风压应力的大小和分布,结果发现树种的受害程度与其树冠形状密切相关,从而探讨其受灾原因,为海堤防护林营林树种的选择和经营管理提供理论依据。
顾沈华等(2010)用典型抽样方法,以冠径/树高为胁地作用评价指标、以冠长/树高为防风能力指标、以枝梢折断率、风倒(折)率为抗风能力指标,对浙江省嘉兴市17个主要沿海基干林带造林树种的防护效能进行评价,结果表明:池杉和水杉胁地最小,合欢、香樟、重阳木和垂柳胁地最为严重;落羽杉、水杉、池杉和银杏防风功能最强,枫香、香樟、喜树和黄山栾树防风功能最弱;池杉、山杜英、枫香、乐昌含笑、落羽杉、香樟和银杏抗风能力最强,美洲黑杨的抗风能力最弱。应松康、张晓勉、高智慧等(2010)在对浙江省沿海基岩质海岸防护林7个主要林分类型的土壤渗透性和理化性质取土测定的基础上,将不同林分类型土壤渗透过程进行拟合,入渗过程大体符合卡斯恰可夫模型,拟合度R2为0.824~0.977,通过初渗率、平均渗透速率、稳渗率、渗透总量等6个土壤理化指标的关系分析,总孔隙度、非毛管空隙度、有机质含量与土壤渗透性极显著正相关,土壤容重则呈极显著负相关;聚类分析表明:北江荛花檵木混交林土壤渗透特性最强,湿地松木荷混交林、湿地松枫香混交林、化香纯林、湿地松纯林土壤渗透性中等,无林地和枫香纯林土壤渗透性较差。
(二)改善土壤生境
在基岩质海岸防护林下土壤生境发生很大变化,土壤理化性质、生物活性得到改良。据陈顺伟等(1995)研究,亚热带岩质海岸不同林分土壤含水率存在显著差异,气象因子与各林分土壤水分相关显著性大小排序为:荒地>湿地松>枫香>湿地松>杨梅,表明荒地土壤水分受气候因子影响最大,在林分中由于林冠、地被物及根系的作用,土壤水分不仅受气候影响,还受到森林植被的调节作用,因而与气候因子相关性较小。而且,营造杨梅林和混交林可提高土壤水源涵养能力,这为沿海岩质海岸造林树种选择提供了科学依据。
在防护林改善土壤理化性质方面,Maily D.对塞内加尔沿海木麻黄林分进行了研究,发现土壤pH由7.3降至6.5,凋落物厚度由4.2cm增至8.0cm,Ca、Al、Mg、Fe、K和P分别增加6.9kg/ ·a、3.7kg/·a、1.8kg/·a、1.3kg/·a、0.49kg/·a和0.25kg/·a。随着年龄的增长,全P和交换性Ca下降,N和有机质分别增加30kg/·a和61.5kg/·a。梁珍海等(1994)以苏北沿海主要防护林树种刺槐和水杉为研究对象,以农田和滩涂作对照,从土壤盐分含量、时空分布规律等方面,揭示了苏北沿海防护林对土壤脱盐及其脱盐稳定性的影响,表明在具备一定水利设施基础上,造林和农业耕作都能加速土壤脱盐过程,但在稳定脱盐效果,防止旱季“返盐”等方面,林业措施更为有效。胡海波等(1994)从土壤物理和化学性状两方面对江苏中部沿海泥质海岸防护林改良土壤的功能进行了探讨,结果表明,防护林能减少土壤容重,增大孔隙,显著提高含水量,促进土壤形成良好的团粒结构且林分年龄越大的结构性越好,同时它也能提高土壤肥力,加快土壤熟化。
·a、3.7kg/·a、1.8kg/·a、1.3kg/·a、0.49kg/·a和0.25kg/·a。随着年龄的增长,全P和交换性Ca下降,N和有机质分别增加30kg/·a和61.5kg/·a。梁珍海等(1994)以苏北沿海主要防护林树种刺槐和水杉为研究对象,以农田和滩涂作对照,从土壤盐分含量、时空分布规律等方面,揭示了苏北沿海防护林对土壤脱盐及其脱盐稳定性的影响,表明在具备一定水利设施基础上,造林和农业耕作都能加速土壤脱盐过程,但在稳定脱盐效果,防止旱季“返盐”等方面,林业措施更为有效。胡海波等(1994)从土壤物理和化学性状两方面对江苏中部沿海泥质海岸防护林改良土壤的功能进行了探讨,结果表明,防护林能减少土壤容重,增大孔隙,显著提高含水量,促进土壤形成良好的团粒结构且林分年龄越大的结构性越好,同时它也能提高土壤肥力,加快土壤熟化。
B.SEELY(1998)对Waquoit Bay 沿海防护林不同土壤质地上N的转化和流失进行了研究。于雷等(1998)在辽河三角洲对海堤重盐土柽柳防护林定位观测研究表明,柽柳林具有改善土壤理化性能效应,不仅能降盐碱,而且能改善土壤养分,主要根系层土壤容重降低,孔隙度和田间持水量显著增加。
张金池、臧廷亮等(2001)也认为,森林能有效改善土壤理化性质,在0~60cm内,湿地松、马尾松、日本扁柏、毛竹四种林分土壤容重分别降低0.037g/ 、0.067g/、0.082g/和0.103g/,非毛管孔隙度提高0.88%、1.68%、2.59%和2.85%,总孔隙度提高1.36%、2.56%、3.86%和3.87%。
、0.067g/、0.082g/和0.103g/,非毛管孔隙度提高0.88%、1.68%、2.59%和2.85%,总孔隙度提高1.36%、2.56%、3.86%和3.87%。
谭芳林等(2003)选择东山1代、2代和惠安1代、2代、3代林分为研究对象,分别采集土壤进行对比研究。结果表明,木麻黄多代连栽后:①随着栽植代数的增加,沙地土壤的pH表现为降低趋势,而土壤的水解性总酸度、交换性总酸度、交换性氢和交换性铝含量等指标随着木麻黄连栽代数的增加而增大;②沙地土壤中水解性氮、铵态氮、有效磷、速效钾、全碳和全氮的含量随着代数的增加而降低,而且木麻黄连栽对表层土壤中上述养分的降低更为显著,但木麻黄连栽对缓效钾的影响并不明显;③土壤中阳离子交换总量逐代增大,这主要与交换性钙和水解性总酸度逐代增大有关;同时,对沿海沙地湿地松林地土壤养分含量及酶活性研究,结果表明,在1代木麻黄的采伐迹地上栽植湿地松,土壤pH随着湿地松林龄的增大而降低,到26a时土壤呈较强的酸性,湿地松可能导致沿海沙地土壤的酸化。湿地松林地土壤中养分含量随林龄不同而表现出不同的变化趋势,但与2代木麻黄纯林相比,湿地松林分在26a时的养分含量总体表现为降低,其中有效磷、速效钾和全氮含量的降低更为明显;阳离子交换总量反映了土壤的交换性能和肥力水平。分析表明,在1代木麻黄的采伐迹地上栽植湿地松对20~40cm土层阳离子交换总量的影响不大,对0~20cm土层中阳离子交换总量的影响主要表现在6龄之后,即随着湿地松林分的生长,该土层中阳离子交换总量逐渐升高,到26a时比木麻黄2代纯林相应土层高21.6%。
土壤微生物也反映了其生物活性水平,通过对亚热带岩质海岸土壤微生物数量分布及与酶活性、理化性质关系的研究,得到的结果是土壤中各类微生物极显著相关,其数量关系是细菌>放线菌>真菌;不同林地土壤微生物数量差异很大,经济林土壤微生物多,马尾松和杨梅成林反而较少;随着深度增加,土壤微生物减少,但在同一层次中各类微生物所占的比例大多比较接近;土壤微生物数量与酶活性有一定的相关性,但相关系数不高,而与土壤理化性质密切相关。土壤微生物数量可以反映土壤肥力水平的高低。
在土壤生物活性方面,胡海波等(2001)对亚热带岩质海岸主要防护林类型土壤酶活性特征进行了研究,指出在3种磷酸酶中,酸性磷酸酶活性大于碱性、中性磷酸酶活性。3种磷酸酶与总磷酸酶活性密切相关,但以碱性磷酸酶相关性最高,中性磷酸酶次之,酸性磷酸酶最小。蔗糖酶、磷酸酶、蛋白酶、脲酶4类土壤酶活性自上而下均逐渐减少。不同森林类型土壤酶活性差异显著,林龄越大酶活性越高;经济林受人为干扰,土壤酶活性居中低水平,应增施有机肥料、尽可能减少对林地的干扰,采用合理的经营管理措施。
胡海波等(1998)研究了苏北淤泥质海岸防护林土壤磷酸酶活性的特点以及几种酶活性的分布规律。结果表明,在滨海潮土中磷酸酶以碱性磷酸酶为主,占总磷酸酶活性的62.6%,因而可用碱性磷酸酶活性反映整个磷酸酶活性;土壤酶活性在剖面和水平方向上均存在差异;在林地中,林龄越大其土壤酶的活性越高;农田的土壤酶活性与林粮间作的幼林相似。谭芳林等(2003)选择东山1代、2代和惠安1代、2代、3代林分为研究对象,对木麻黄连栽对沿海沙地土壤养分及酶活性的影响进行试验。结果表明,沙地土壤中脲酶的活性没有明显的变化,而磷酸酶、多酚氧化酶和过氧化物酶的活性则随代数的增加而降低。在1代木麻黄的采伐迹地上栽植湿地松对脲酶活性的影响似乎不太明显,但磷酸酶、多酚氧化酶和过氧化物酶活性随湿地松林龄的增大而降低。与木麻黄2代纯林相比,各湿地松林分土壤中磷酸酶活性均较低,但多酚氧化酶和过氧化物酶活性在11a之前表现为略高或与之相当,11a之后,二者的活性在湿地松林分中较低,尤其在表层土壤中。总体而言,在1代木麻黄防护林的采伐迹地上栽植湿地松有使土壤磷酸酶、多酚氧化酶和过氧化物活性降低的趋势。
田保状、乔勇进等(2005)通过对沙质海岸防护林体系前沿的草带、灌木带、基干林带、经济林种植带、农田林网、梯田地埂水土保持林和山地丘陵水源涵养林等7部分的土样养分含量测定,结果表明:不同区域立地土壤及层次肥力存在显著差异,全氮在整个海防林体系中含量很低;速效氮在草带、灌木带和基干林带含量较低,而在其他带含量则较高;速效磷在基干林带、经济林种植带和农田林网土壤中含量较高;速效钾含量除草带外总体变化不大;有机质含量普遍较低。李春艳、李传荣、许景伟等(2007)研究了东营泥质海岸不同林分下土壤养分、微生物数量、土壤酶活性及分布特征,结果表明:①土壤微生物、土壤酶和土壤养分主要分布在0~20cm的土层,20~50cm的较少;②不同林分微生物数量差异显著,白蜡林微生物数量最多,柳林、枣园数量较少,柽柳林数量最少;③不同林分土壤酶的活性表现出差异性,各种酶之间过氧化氢酶活性最高;④土壤养分含量差异显著,柳林土壤养分含量相对较高于其他样地,白蜡和桑园土壤养分含量次之;⑤林地土壤微生物数量和土壤酶活性与土壤养分密切相关。
李玥、张金池等(2010)以上海市沿海防护林为研究对象,选择6种不同树种的防护林带,采集0~10cm、10~20cm、20~40cm、40~60cm四层土样为研究材料,运用典型相关分析法,对防护林地土壤养分因子、微生物因子和酶活性因子中每两组变量间的相关性进行了分析。结果表明:三组变量土壤养分、微生物、酶活性中,每两者之间均有显著的典型相关变量存在,而且基本能够代表变量总体相关信息;土壤养分和土壤微生物间的相关主要由全氮、速效磷含量与微生物生物量氮、微生物生物量碳和微生物生物量磷引起;土壤养分与土壤酶活性间的相关性主要由全氮、有效磷、水解氮含量与脲酶、蛋白酶活性的相关性引起;土壤微生物与土壤酶活性间的相关性主要是由微生物生物量氮、微生物生物量磷与脲酶、蛋白酶、碱性磷酸酶活性的相关性引起;不同林地不同土壤层次的养分、微生物及酶活性在各对典型变量上的聚集趋势可为防护林建设过程中的树种选择与土壤健康诊断提供一定的依据。刘骄、黄义雄、叶功富等(2011)通过采用平均木法对福建海坛岛主要防护林树种即木麻黄、台湾相思、黑松、湿地松的生物量进行测定,并通过凋落物的收集和土壤养分的分析了解这4种树种的生长习性以及养分丰缺情况。结果表明:①生物量总量从大到小依次为湿地松(270.84t/)、黑松(125.72t/)、台湾相思(75.99t/)、迹地更新不良木麻黄(56.02t/);②从凋落物的研究情况可知9月是四树种共同的生长月,凋落物归还量很高,而对于木麻黄和台湾相思而言,另一个凋落物归还量的高峰期是5月;③对于四种树种林下土壤养分的测定,可以看出台湾相思林下土壤养分相对丰富,而总体来说四种树种的林下土壤营养物质相对缺乏,影响到防护林的生物量进而影响其防风固沙、保持水土的性能。高智慧等(2011)根据沿海基岩质海岸不同的植被类型典型地段,在浙江省三门县浬浦镇草头村选择了6个试验样地和1个无林地作对照进行人工促进植被恢复对基岩质海岸防护林的土壤生态特性影响研究,结果表明:①沿海岩质海岸防护林土壤含水率普遍较低,并随着土层增加相应减少。经人工促进植被恢复的化香纯林土壤含水率最高,北江荛花檵木混交林土壤比重最小,二样地总孔隙度和毛管持水量增加,其他各样地间差异不明显。②沿海岩质海岸防护林土壤养分含量差异较大,试验样地的土壤全氮、水解性氮、全磷、速效钾及有机质含量均明显高于对照地,其中土壤全氮、水解性氮、全磷及有机质含量均以经人工促进植被恢复的北江荛花檵木混交林最高,土壤速效钾含量以化香纯林最高。
国外的研究认为红树林生态系统在热带沿海地区营养循环和能量流动中,起着重要的作用(Odum and Heald,1974; Cintrón and Schaeffer Novelli,1983; Camilleri and Ribi,1986; Flores Verdugo et al.,1987; Lacerda et al.,1993; Silva,1996; Silva and Mozeto,1997)。
对于工业化给红树林带来的污染,国外众多学者对此进行过研究和探讨。他们认为红树林能够积聚含硫化物的有机物质,从而可以长期地储存重金属污染物(Harbinson,1981; Harbinson,1986a; Silva,1996; Lacerda,1997)。野外调查和试验研究都表明,红树林生态系统对重金属的捕捉是非常有效和迅速的(Montgomery and Price,1979; Harbison,1981; Harbison,1986a; Harbison,1986b; Tam and Wong,1993)。
C.A.R.Silva(1998,The Dynamics of Heavy Metals through Litterfall and Decomposition in A Red Mangrove Forest)对Zn、Mn、Fe这3种重金属元素在巴西东南红树林森林生态系统中的循环进行了研究,认为在热带沿海地区,红树林生态系统作为一个有效的生物屏障,能够减少重金属进入土壤和海洋的数量,并对在红树林内生态系统内循环的重金属进行了量化。
Miguel等 (2003,Seasonal Dynamics in Dissolved Organic Carbon Concentrations in a Coastal Water-Table Aquifer at the Forest-Marsh Interface)对美国加利福尼亚南部森林和沼泽交界处的地下水中可溶性有机碳的季节动态进行了研究。
J.J.Zhu等(2002,Salt Distribution in Response to Optical Stratification Porosity and Relative Windspeed in A Coastal Forest in Niigata,Japan)在日本Niigata岛进行了森林的垂直结构和风速对盐分分布的影响的研究。结果发现海盐的分布和林冠内的风速都与森林的垂直结构紧密相关,通过建立线性回归模型,可以根据森林的垂直结构来预测海盐的分布以及海防林林冠内风速的变化。这一结构可以用来分析海防林植被的含盐量和评价海防林的过滤盐分功能。Marianne K.等(2003)为了研究洪水对植物根的影响,在美国东南沿海2个相邻沼泽地内对4种橡树根的变化和洪水对它们的影响进行了研究。
(三)调节气候及提高产量
森林具有显著的生态功能,可调节气候,改善环境,主要表现在降低风速、调节气温和提高空气湿度等方面,在沙质海滩还能起到防风固沙的作用。1983年,广东省在陆丰县东海岸林场和惠东县港口林场对海岸防护林带的防风效应进行了测定,结果是林内风速显著低于林外,尤以幼林内分枝低,降低风速效能更强。当林外风速接近4m/s时,幼林内风速不到林外的15%,而中壮龄林分分枝较高,林内风速约为林外风速的40%~50%。防护林密度不同,林内降低风速的效应也有差异,密度越大,降低风速的效应越明显。海岸防护林还有调节气温、土温和空气湿度、土壤湿度的功能,林内气温比林外裸地低,最大温差出现在午后13~14时,9年生林分温差达4.2~5℃。林内空气湿度和土壤湿度比林外高,林带附近高于空旷地沙滩。
由于改善了气候条件,为农作物生长创造了一个良好的生态环境,提高了农作物产量。在辽宁省新金县,沿海防护林可使平坦耕地粮食增产5.0%~18.2%,坡耕地增产48.6%~119.0%。浙江省温岭市防护林网内的柑橘,在一般年份增产11.7%,1979年夏高温干旱,产量提高18.8%~30.6%,说明在遇到气象灾害时,防护林的增产效果尤为明显。
董晓敏、周学东(1994)从蒸发过程的物理机理出发,在具有湿润海洋性气候特征的江苏北部沿海地区,对防护林区域1974~1991年的月蒸发量进行时、空对比分析,得出随林木的生长成林对蒸发的影响,肯定了苏北沿海防护林体系调节气候的积极效应。周学东、董晓敏等(1995)利用多元统计回归方法,消除气候特征量自身随时间的演变和空间分布的差异,以及一些非线性因素的影响,从而揭示了江苏北部沿海森林环境的时、空变化对区域性气候产生的影响,得出防护林具有落叶期升温降温、叶生长期降温增湿的效应,所在区域的风速和蒸发量有全年明显减小趋势的分析结果。黄承标等(1999)对广西北热带山口林场1.5年生和4.5年生两种林龄窿缘桉防护林内及空旷地的主要气象要素进行定位对比观测。结果表明两种林龄的防护林防风效果显著,并都有一定的降温增湿作用。
王贵霞、许景伟等(2004)对胶南沿海黑松等幼龄人工混交林带的气候和土壤生态因子进行了测定,与空旷地相比较,结果表明:混交林带具有明显的调节气温和地温,减弱风速,增加空气相对湿度的作用;具有良好的改善土壤结构作用,能增大土壤孔隙,减少土壤容重,增大土壤的饱和持水量和强化入渗的功效。采用特尔斐法对其效益进行定量化评价,得出该混交林带的气候效益指数为-0.2975,改良土壤效益指数为-0.093,综合环境效益指数为-0.39。王宗星、虞木奎等(2010)为分析上海市郊沿海防护林的防护效应,于2009年夏季对人民塘水杉基干林带前后不同观测点的风速、气温和空气相对湿度等气候因子进行了观测,并对该林带前后1km处的防台风效应进行了比较;另外,以滨海森林公园的红叶椿林分为观测对象,比较分析了林带内不同垂直高度处的防风效应。结果表明,水杉基干林带具有明显的防风效果,林带内至林带后65m范围内的风速较林带前均有不同程度的下降,以林带后25m处的风速最低;在“莫拉克”台风登陆期间,水杉基干林带在较大尺度上具有明显的降低风速的作用;水杉基干林带在一定程度上能降低局部气温、提高空气相对湿度,以林带内的气温最低、空气相对湿度最高,从而达到改善林带后小气候的作用。红叶椿林分内不同垂直高度处的防风效应有一定的差异,其中垂直高度2m处的风速最大,而树冠处(垂直高度5m)的风速最低。林宇(2011)应用回归分析的方法,分析主要气候因子对沿海防护林卷荚相思树高、胸径生长的影响。结果表明,气候因子对沿海防护林卷荚相思树高生长的影响为年日照时数>年最冷月平均气温>月平均气温>年最热月平均气温>月均相对湿度;气候因子对沿海防护林卷荚相思胸径生长的影响为年日照时数>月平均气温>年最热月平均气温>年最冷月平均气温>月均相对湿度。其中,年日照时数、年最冷月平均气温、年最热月平均气温、月平均气温与卷荚相思树高和胸径的生长量有极显著的正相关关系。
我国沿海岩质港湾海岸广泛分布,地势起伏的裸露丘陵山地,降水时会形成大量地表径流并引起土壤侵蚀;天旱时又出现缺水的景象,季节性缺水是我国沿海气候的重要特点,常给防护林体系建设带来不利影响。森林可发挥涵养水源和保持水土的作用,防止河道、水库淤塞,并能长期源源不断地流出清洁的淡水。在海岸岛屿的丘陵山地上,由于淡水奇缺,森林涵养水源、保持水土的功效更为显著和重要。
(一)涵养水源
(二)保持水土
森林植被以茂密的多层树冠截留降水、深厚的死地被物吸收降水、盘根错节的根系固持土壤,再加上土壤非毛管孔隙大,几乎不产生地表径流,即使有少量的地表径流也只能在死地被物中缓慢流动,因而在森林保护下地表径流和土壤侵蚀量都很小。据浙江嵊县上东水土保持试验站测定,有森林植被覆盖的山坡地要比各种不同开垦种植的地面水土流失量少得多。在200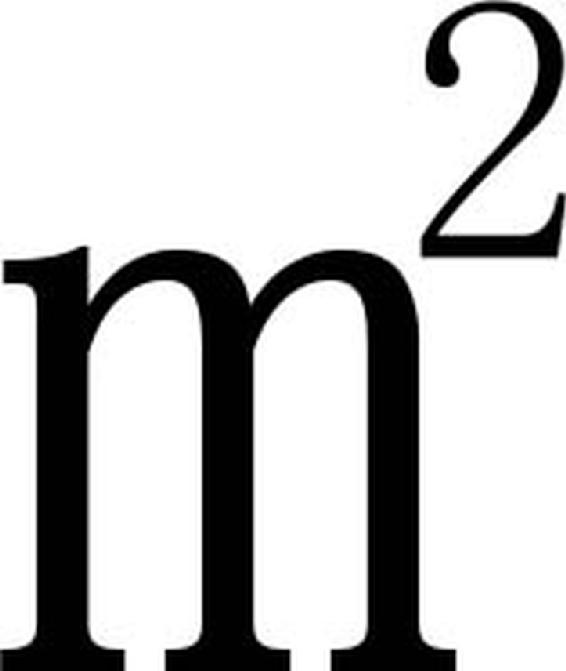 的径流小区上,经历一场289mm的暴雨后,旱坡耕地流失土壤1383.3kg,顺坡茶园986.2kg,而覆盖度分别为80%、60%和95%的杉木幼林、灌木林及松树幼林,其土壤流失量分别为50.1kg、80.3kg和0.0kg,森林植被保持水土的效果十分显著。高智慧等(1999)通过设立径流小区和多元回归分析方法,对亚热岩质海岸不同植被类型的水土保持效益进行了对比研究,结果表明即使林分处于幼龄阶段,不同植被类型的地表径流也比对照低9.05%,侵蚀模数比对照低32.29%。
的径流小区上,经历一场289mm的暴雨后,旱坡耕地流失土壤1383.3kg,顺坡茶园986.2kg,而覆盖度分别为80%、60%和95%的杉木幼林、灌木林及松树幼林,其土壤流失量分别为50.1kg、80.3kg和0.0kg,森林植被保持水土的效果十分显著。高智慧等(1999)通过设立径流小区和多元回归分析方法,对亚热岩质海岸不同植被类型的水土保持效益进行了对比研究,结果表明即使林分处于幼龄阶段,不同植被类型的地表径流也比对照低9.05%,侵蚀模数比对照低32.29%。
张金池等(1996)研究了苏北海堤刺槐、水杉和柳杉防护林的防护效益。认为森林植被可有效地减低地表径流,对地表径流过程的影响,主要表现在减洪和滞洪方面;研究苏北海堤主要防护林防蚀能力得出:土壤侵蚀量柳杉林<水杉林<刺槐林<无林区,年土壤侵蚀模数分别为922.2888t/·a、1231.2822t/·a和2608.4538t/·a。无林区发生土壤侵蚀的临界降雨量为5~8mm,有林地则为8~10mm;对照区大于50mm降水引起的土壤侵蚀量占全年土壤侵蚀量的80.5%,有林地占90%,表明雨量较大,尤其是大暴雨和特大暴雨往往引起较为严重的土壤侵蚀。雨量、最大某时段的降雨强度与土壤侵蚀量相关密切,平均雨强度次之,与降水间隔期相关极差。另外,在主分量分析基础上确定了影响土壤侵蚀量的主要降水特征因子,结合各林分冠层截留量,得出各地段坡面土壤侵蚀量预报模型。
为了揭示森林防蚀固土的机理,张金池等(2001)研究了细根生物量、年生长量、年死亡分解量和细根周转的变化,测定树木根系对土壤抗冲、抗蚀和渗透性的强化作用以及水土保持效益,筛选出了既适合当地立地条件,又具有较强水土保持功能的树种,为今后指导沿海防护林体系建设提供了科学依据。结果表明,岩质海岸防护林根系强化土壤抗冲性的机理在于根系通过≤0.5mm和0.5~1.0mm两种径级细根的直接网络作用。与无根系土壤相比,湿地松、马尾松、日本扁柏、毛竹四种林分≤1mm的细根在0~60cm土层内提高土壤抗冲值平均分别为0.400、0.414、0.458和0.526,且强化值与径级≤1mm的根量服从线性回归关系。根系强化土壤抗蚀性的实质,是径级≤1mm的细根提高了土壤中水稳性团聚体的含量,上述4个树种≤1mm的细根能使>0.25mm土壤水稳性团聚体含量分别提高3.66%、8.83%、19.53%和28.8%。岩质海岸防护林能显著提高土壤渗透性,其强化值与根系中≤0.5mm和0.5~1.0mm的根量呈极显著的相关性。各防护林类型可降低地表径流量和土壤侵蚀量,杨梅和桃形李混交林、湿地松、日本扁柏3种林分地表径流比对照分别减少22.53%、35.12%和39.02%,土壤侵蚀量分别减少29.73%、58.28%和62.45%,最后建立了地表径流和土壤侵蚀量的回归方程,估测精度在80%以上。
余坤勇(2009)根据美国USLE土壤侵蚀模型,实现了对福建省漳浦县沿海防护林水土保持的动态监测,并根据土壤侵蚀所流失有机质的经济损失实现了对沿海防护林防护效益的评价。研究结果表明:沿海防护林能明显地提高水土保持效益,减少经济损失。黄晓芳(2010)根据美国USLE土壤侵蚀模型,实现福建省漳浦县沿海防护林水土保持动态监测,并根据土壤侵蚀所流失的有机质的经济损失来实现沿海防护林的防护效益的评价。
(三)土壤侵蚀强度分级
沿海岩质海岸降雨量大、地势起伏不平,土壤侵蚀的潜在危险性大。若植被受到破坏,土壤侵蚀将十分严重,不仅使环境恶化,而且还危及工农业生产及交通、水利等事业。因此,在岩质海岸需弄清水土流失成因、类型、特点,对土壤侵蚀强度进行分级,以便因地制宜地采取相应的预防措施。张金池等(2003)曾对苏北云台山区水土流失强度遥感分级方法进行过探讨,通过模式小班调查、建立判读标志和遥感图像目视判读等方法,进行图斑面积勾绘和各图斑向量确定。运用多元统计方法,首先对60块模式样地水土流失强度等级进行划分,然后根据数量化方法Ⅱ对170块遥感判读样地进行归类,并统计不同侵蚀强度的面积。此外,还按立地条件和水土流失程度的差异,为各类型区提出了树种选择、经营管理和开展水土保持工作的建议,具有较高的理论和实际应用价值。
我国岩质海岸尤其是临海一面坡,土壤贫瘠、干燥,常年风大,夏季又易遭受台风袭击和山洪危害,造林难度很大。几十年来,各岸段林业生态工程建设虽经艰苦努力但成效不大,造林树种单一、结构简单,有些地方甚至导致严重的水土流失。因此,为提高造林成活率和保存率,促进树木生长,提高森林的生态、经济和社会效益,开展营造技术研究具有重要的理论意义和应用价值。
(一)造林技术
张敦论等(1999)针对山东沙质海岸土壤保水性能差的特点,进行高分子吸水剂对沙质海岸土壤物理性能及造林成活率的影响的研究。结果表明,施用高分子吸水剂能显著增强土壤的吸水和持水能力,提高黑松造林成活率。高分子吸水剂施入量以10~20g/株为宜。
蒋妙定等(1996)对岩质海岸湿地松造林技术进行研究,总结出“挖穴造林、适当密植、掌握时机、深栽紧打”的技术要领,另外指出湿地松适生苗造林应选1年生良种壮苗,营造混交林,栽植时既可用钙镁磷肥打浆蘸根,亦可直接施放于穴内,这样可显著提高造林成活率,促进林木生长。陈顺伟等(2001)通过对岩质海岸防护林造林技术的研究,提出了采用阔叶树以60cm见方、针叶树以30cm见方的穴状整地;落叶阔叶树截干、ABT蘸根造林;针叶树磷肥蘸根、集团造林,以及容器苗造林等造林配套技术,确保造林保存率达90%以上。
杨运立等(1994)在福建重盐碱地上采用造林前(3~5个月)整地,选择抗盐碱害的粗枝木麻黄容器育苗造林,并采用浅植高培土造林和保护幼林地杂草植被的抚育等技术,使土壤含盐量下降50%~68.7%,造林成活率由8%提高到85%~88.5%。柯玉铸等(1996)针对木麻黄防护林二代更新困难问题,采用水培繁育新技术,培育速生、抗病木麻黄幼树苗木,利用速生、抗病优树水培苗,在福建长乐、平潭两地进行更新造林试验。发现幼林阶段优树水培比实生苗造林成活率增长14.3%~34.5%,林分保存率增长21.7%~59.7%,树高增长75.7%~88.4%,胸径增长89.4%~165.5%,显示了应用速生、抗病木麻黄优树水培苗进行更新改造是解决木麻黄二代更新困难的一个有效途径。张水松等(2000)在福建省惠安县赤湖林场和东山县赤山林场木麻黄基干林带前沿风口干旱沙地,进行提高木麻黄造林效果配套技术研究。经2年试验研究,在分析风口困难立地造林失败主要原因基础上,从木麻黄抗逆品系选择应用,因地制宜确定造林方式,采用深挖整地、放客土、拌泥浆、提早造林季节、选择大雨天冒雨造林、大苗深栽等抗旱造林配套技术以及旱季培土抚育保墒、浇水保苗和筑沙堤设风障防潮防风工程措施相结合的造林方法,在造林屡次失败的风口沙地将木麻黄造林保存率提高到80%~90%。在造林技术上取得的重大突破,为海岸带风口困难立地国家特殊保护林带工程建设提供了有益的经验。何小广等(2004)认为,淤泥质海岸是浙东南滨海的主要海岸类型,也是我国沿海的淤泥质滩涂集中分布区。这一范围的土壤主要为滨海盐土,分布涂泥和咸泥两个土属,特性与沙土土属差异很大,具有土层深厚、质地黏重(黏粒<0.01mm占90.5%)、自然肥力高的特点。据测定,有机质含量一般为2.19%~4.58%,全氮0.09%~0.15%,全磷0.08%,全钾2.3%,含盐量0.21%~1.58%,碳酸钙5.6%~7.3%,pH8.25~8.97,含盐量较高,海风长驱直入,蒸发强度大,下层土壤盐分随水分上升,极易造成表层土壤返盐返碱,脱盐洗碱过程缓慢,土体收缩率大,成为各类作物生长的限制因子。因此,在造林前均需进行土壤改良。一般绿化造林可采取海涂筑堤围塘、海塘地规划、树种配置、林木引种驯化、苗木培育、整地、适时造林、幼林抚育管护等技术措施。皮宝柱(1999)调查分析、总结了泥质海岸柽柳林营造技术,摸索出一整套重盐碱地造林新方法。即通过人工开沟、自然播种、植苗、插条等方法,使光板盐碱地上长出了翠绿的柽柳,使难以利用的重盐碱地得到了充分的利用,既改善了生态环境、增长了绿色植被,又收到了良好的经济、生态、社会效益。许基全等(1998)针对泥质海岸的特点,提出了筑堤围涂、开挖河渠、合理规划、适地适树、引种驯化、壮苗培育、合理整地、适时造林、适当浅栽、抚育管护等10项造林措施。
为探讨黑松人工林抚育间伐效果,钟鼎谋等在山东刘公岛林场设置4块黑松人工林固定标准地,进行间伐强度为0%、15%、20%和25%的抚育间伐试验,并分别在1974年和1983年以相同处理方式实施间伐。1993年调查结果表明,抚育间伐能显著促进林木生长,林分蓄积量提高12%~28%,以间伐强度为20%和25%的效果较好;间伐使林分结构改善,林内幼苗幼树的生长也明显优于不间伐林分。
林承超等(1995)对福州琅岐岛朴树群落进行了全面调查,分析了朴树群落的种类组成、区系成分、群落结构和生活型组成等群落特征,并将琅岐岛朴树群落下土壤性质与平潭县芦洋农场木麻黄林下土壤性质作了比较分析,提出了福建沿海防护林更新与改造的具体意见,即朴树在福建滨海风沙土上种植是可行的,朴树与木麻黄混交种植将会减轻木麻黄的病虫害,提高木麻黄林下土壤有机质含量,改善土壤结构。
方正贺(2007)分析了福建省沿海防护林体系国债造林难度和影响造林质量的主要因素,提出了提高国债造林质量的对策。陈侃等(2009)进行了北仑区沿海防护林树种选择与造林模式研究,并结合试验林带进行造林模式研究,提出几种优化配置模式。李恭学等(2010)对新围垦滩涂防护林树种选择及造林关键技术进行了研究,依据生态学原理,通过采用一系列生物、工程和管理措施,提高了造林试验树种的成活率,探索出入侵生物水葫芦的应用途径及特困立地造林的有效措施,也为沿海防护林体系建设提供了可借鉴的经验。陈忠(2011)对沿海防护林水杉的造林技术进行了探讨,张尔等(2011)浅述了木麻黄沿海防护林造林技术。
在挪威和芬兰沿海地区,Hame T.等结合地面实地调查,利用卫星资料对寒温带针叶林的生物量(有机质)进行估算,为森林经营和森林资源动态监测提供了依据。为掌握林分卫生状况,及时采取经营管理措施,Busing R.T.等利用林冠层的枯死木来监测林分的卫生状况。红树林作为抗盐碱树种在世界沿海地区广泛种植,国内外学者也对它的恢复和保护进行了深入的研究。F.Rasool等(2002)在巴基斯坦海岸从规划设计、育苗、幼树移植等方面对红树林生态系统恢复进行了探讨。
(二)病虫害防治
森林病虫害是无烟的森林火灾。我国基岩质海岸生态环境恶劣,常暴发森林病虫害,给沿海防护林体系建设带来严重威胁,因此应对病虫害发病规律和防治对策进行深入研究。
在浙江沿海地区,方惠兰等对木麻黄害虫进行调查研究,发现害虫有6目26科73种,其中以星天牛分布最广,危害最严重,在该省发生以1代为主,幼虫在寄主树干木质部隧道中越冬。试验结果表明,用磷化锌毒签防治幼虫较为理想,校正虫口减退率为94.4%。
我国沿海木麻黄林主要病害有青枯病和立枯病,在美国佛罗里达州,由真菌假蜜环菌引起的蘑菇根腐病使木麻黄的死亡率很高。主要虫害有木麻黄毒蛾、黄星天牛和多斑豹蠹蛾,此外皮暗斑螟、木麻黄潜叶蛾和龙眼蚁舟蛾亦很频繁。在国外,黑蛀虫也是木麻黄的重要害虫,在尼日利亚,木麻黄幼苗还常受到白蚁危害甚至死亡。针对木麻黄的病虫害,陈云征认为,主要防治对策有化学防治、生物防治(微生物、线虫、蚂蚁和肿腿蜂等)、抗性育种、营林措施(如营造混交林)及综合治理等方法。
刘清浪等(1999)筛选出蝗虫微孢子虫、蜡状芽孢杆菌及簇孢霉等微生物杀虫剂,并应用于防治棉蝗,感染率达77%以上;应用从星天牛中分离的白僵菌防治星天牛,感染率达77.8%;应用芜菁夜蛾线虫和蠹蛾线虫防治星天牛,感染率达90%以上,为沿海防护林木麻黄病虫害的综合控制提供科学依据。黄金水等(1998)认为苦楝树是星天牛成虫最喜欢取食的补充营养植物,有强烈的诱集作用。以苦楝树做诱饵树集中诱杀沿海木麻黄防护林星天牛是一种很有效的防治方法。
方丽等(2001)对日照沿海防护林17种主要病虫害发生原因进行了全面分析,提出了相应的防治对策。郭祥(2002)阐述了福建沿海防护林主要病虫害的可持续治理对策。吴马愿(2010)对晋江市沿海防护林主要害虫及其防治进行了研究,调查木麻黄害虫的危害部位和生物学特性,提出相应的防治对策,对于保护沿海区域生态环境具有重要的现实意义。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












