



一、淡水水体
淡水水体包括泉、河流、溪流、湖泊。一类为流动水(流动水环境),如泉、溪流和河流,另一类是不流动水(不流动水环境),如湖泊、池塘和湖沼。淡水环境有非常不同的组成和相应特征的微生物群落。
1.泉
泉的种类多样,有冷泉、热泉、矿泉(含硫、镁)、酸泉、放射性泉等。泉水中的微生物主要是细菌和藻类。一般在光合作用群体(光合细菌和藻类群落)占主体的泉环境中,其范围为102~108细胞/ml。在泉的浅层及岩石表面因有充足的光,无机初级生产者的数量可达106~109细胞/ml。在泉水中由于营养物特别是DOM(dissolved organic matter)含量低,异养细菌数量通常相当低(10~106细胞/ml)。在泉水中有机物最初源于光合群体,可以想象泉水中DOM和异养细菌的浓度开始是非常低的。但随着其他有机物的输入(源于其他生物和陆地输入),有机物浓度会进一步提高。此时异养菌数和光合藻类的数量会进一步增加。
2.溪流与河流
溪流和河流是源于山泉、降雨的流动水体,其物理化学特征(如温度、流量、速度和化学组成等)取决于流经地域的地理和气候条件,例如流经陡峭山区的溪流有快的流速和较低的水温,而流经平原的河流流速较慢和有较高的水温。河流有的较浅,但有的地方有50米以上的深潭。溪流和河流的流量高度依赖于降雨量和季节的变化。
溪流中含有主要的生产者群落,特别是当光能穿透到溪流的底部的地方,光合作用群体范围为106~108细胞/ml,它们大多数以吸附群落的形式存在,少数也存在于流动水体中,但由于恒定的水运动,它们不是空间上的稳定群体。溪流中的异养群体一般处于较低水平。大多是好氧或兼性厌氧菌。
河流较溪流积累更多的DOM,DOM的增加限制了光的透性,因此光合作用群体的增加受到限制,而异养群体随着于DOM的增加而增加。河流大部分水体整体上通气良好,异养群体中好氧和兼性厌氧菌占优势,其浓度范围大多位于104~109细胞/ml,随DOM浓度增加而增加。在污水注入区域的下游异养群体比其上游增加2~3个数量级。河流中的生物膜和沉积物中的群落是河流中的稳定群体。
溪流和河流的微生物群体大多来自土壤,因此河流和土壤的微生物群落同源,其结构有相似的地方。
3.湖泊
全世界的湖泊种类多样,物理化学特征各异。湖泊深度从数米到1 000米以上。湖的表面变化也非常大,从数平方米的小水塘到100 000km2的巨大湖泊。一般认为湖泊是静止或非流动的,但湖水仍有流入和流出,风造成扰动,温度造成混合,所有这些造成了湖泊的动态环境。许多湖泊有特征性化学组成,如盐湖(高盐度)、苦味湖(富含MgSO4)、Borax湖(富含Na2B4O7)、Soda湖(富含NaHCO3)等。
分层是较深湖泊的重要特征。依形态(深度、宽度、沿岸地学特征、流速等)和化学参数(温度、pH值、氧含量等)的不同被分为沿岸区(littoral zone)、漂浮层(neuston layer)、浅水区(limnetic zone)、深水区(profundal zone)和湖底区(benthic zone)(图1-4)。沿岸区为湖泊边沿,阳光能穿透到底部,生长着茂盛的沉水、挺水植物。漂浮层的水化学特征及微生物参见本章第五节。浅水区是离开沿岸区的开放水域的表层水体,这个区有较强的光照。深水区位于浅水区的下方,光照强度少于阳光的1%(光补偿点)。湖底区位于湖泊直达沉积物的水体的最下部。湖的最下层是沉积物,也称为底层生境。温度对于湖泊非常重要,由于季节变化导致水温变化还会使较深湖的上下层水体发生翻转混合。从温度变化的角度可分为三个区;上层区或表水层(epilimnion)变温层,下层区或下层滞水带、均温层(hypolimnion)和温跃层(thermocline)(图1-5)。由于水在4℃时密度最大,温度导致的密度分层发生在中间的温跃层。在夏天上层水被太阳加热,水温高,溶氧量高,有较高的初级生产力。而下层区正好相反,低温和低营养水平,缺乏光照。这种分层使湖泊水体相对静态,但当秋冬来临时,上层水水温降低直至达到下层水的温度及相应的比重,此时温跃层被打破,并使上下二水层混合。在寒冬上层水结冰,温度位于0~4℃之间,而下层水仍然维持4℃或更高,此时温跃层仍形成,不发生混合。在春天当湖的冰雪融化,上下层水又达到同样的温度,混合再度发生。实际上这二层水的翻转和混合使下层水再充氧和上层水的矿物营养再次得到补偿。较浅的湖泊可只具有深水湖泊的部分特征。
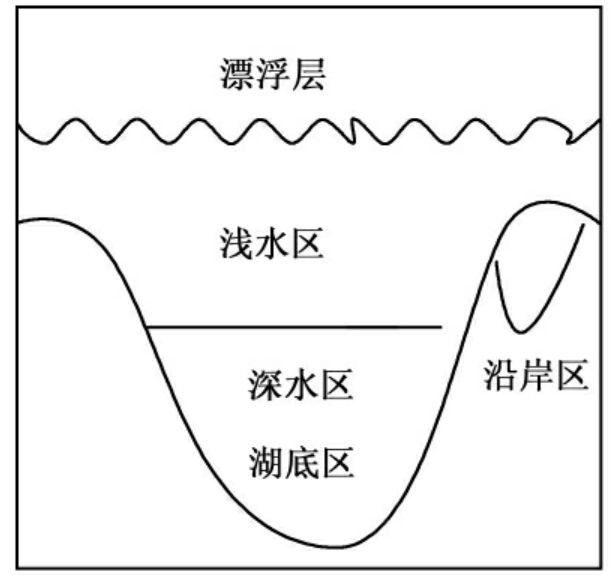
图1-4 深水湖泊水体的分层
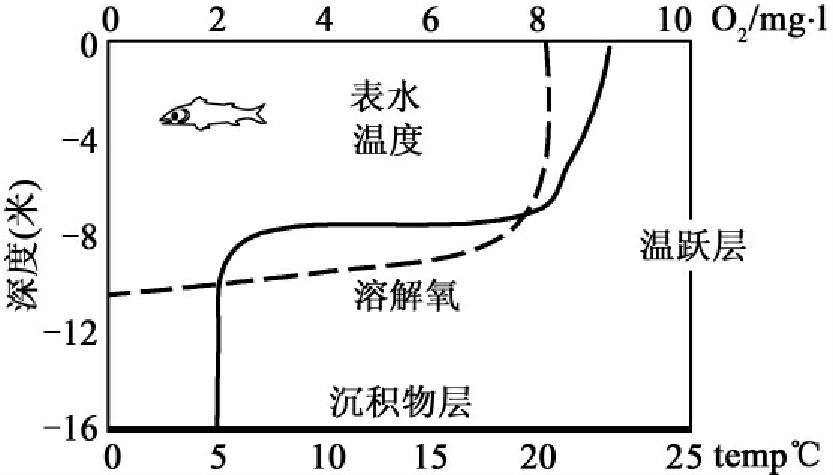
图1-5 从温度变化划分湖泊层次示意图
湖泊水体有大量初级和次级生产群体,有最为复杂、相互作用的微生物群落。光合群体主要分布于沿岸区及浅水区。在沿岸区有大量的初级生产者,有较高的初级生产力。浮游群落中藻类是主要的,而蓝细菌是次要的,而附着群落中的优势种类则是丝状和附生藻类。浅水区也能为光合作用生物提供良好的生态环境,也有处于优势地位的大量浮游藻类,而且其组成因光达到水体的深度和波长的差异而显示出梯度性的变化。图1-6显示了代表性光合生物及其光吸收光谱。从上至下是chlorophycophyta、紫球藻属(Porphyridium)(Rhodophycophyta)、聚球藻属(Synechococcas)、蓝细菌、绿菌属(Chlorobium)、绿硫细菌、红假单胞菌属(Rhodopseudomonas)非硫紫细菌。以绿菌属(绿硫细菌)为例,它们比其他光营养菌更能利用长波长的光,也是厌氧生物,需要H2S而不是H2O进行光合作用。这样它们分布在较深水体或沉积物表面,那里只有少量光穿透,很少或没有氧存在,但有H2S可利用。
湖泊中除有光合群体外,还有大量的异养群体,异养群体产生的次级生产量和初级生产量直接相关。一般在透光区次级生产为初级生产的2%~20%。在水体中异养群体的密度随深度的增加而增加,但有三个区明显有较高的异养群体数量。在漂浮层由于积累了大量的蛋白质、脂肪酸而创造了一种富营养条件,从而有大量的异养群体。另一个区是温跃层,形成于漂浮层及上层的有机碎屑会沉积到那里,因而造就大量异养生物的生境条件。第三个是邻接底泥的底层水体,那里有大量沉降和从底泥中释放出来的有机物供微生物利用,但这些微生物主要是厌氧的。
寡营养和富营养湖泊中由于生物可利用营养物的差异造成水体细菌群体数量及垂直分布上的差异(图1-7)。从图中可见寡营养湖泊上层水中蓝细菌是优势菌,而异养细菌的浓度较低,而在有机物积累位于上层和下层之间的温跃层异养细菌数量大量增加。在富营养化湖泊中尽管分布格局基本一致,但光合作用细菌和异养细菌数量均较前者要高,而且由于富营养湖泊有高得多的有机物,造成混浊,阻碍光透过,使光合细菌的位置上移。其生产力(1~30mgc/m3·day)也低于寡营养湖泊(20~120 mgc/m3·day),相反富营养湖泊的次级生产力速率(190~220 mgc/m3·day)却比寡营养湖泊高得多(1~80mgc/m3·day)。
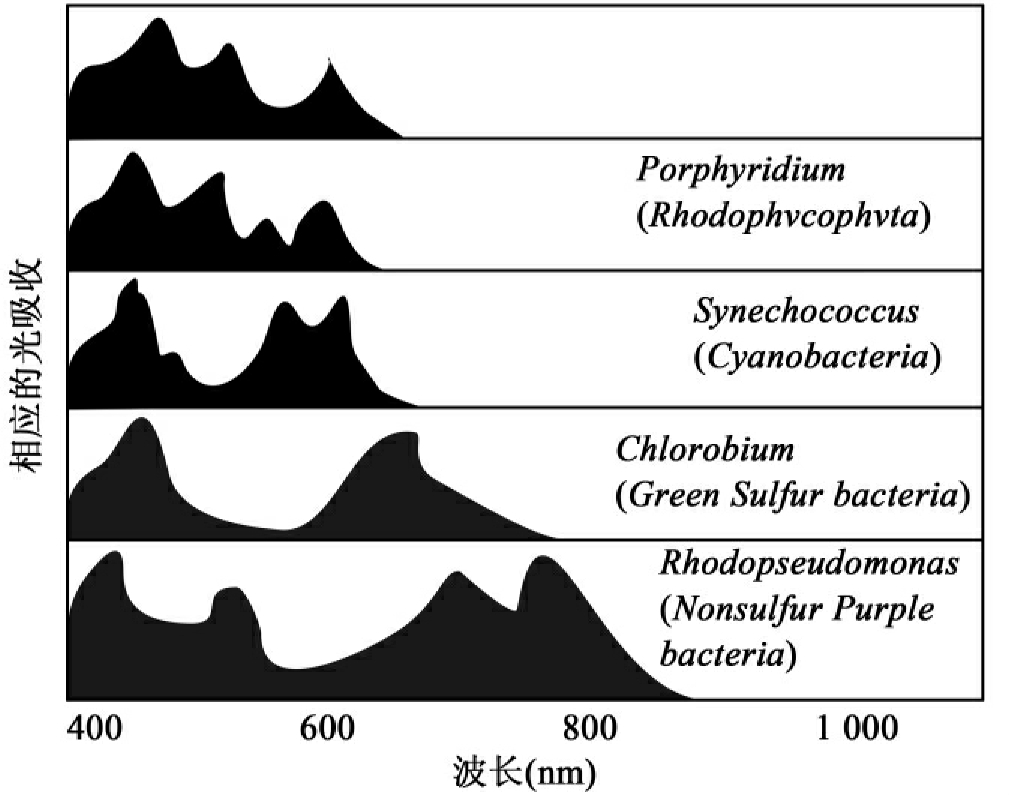
图1-6 不同浮游藻类和光合细菌的光吸收谱

图1-7(A)典型寡营养湖泊中细菌分布示意图。特别注意光合生物群体的分布和浓度(丰富度),也标明了上层水中异养生物浓度较低,但那里蓝细菌占优势。在湖上层和湖下层之间,异养菌大量生长,这与这个带的有机物积累有关。(B)典型富营养湖泊细菌分布示意图,光合和异养群体都存在相当高的比例。
除了细菌和藻类群体外,淡水水体的溪流、河流和湖泊中也有真菌、原生动物和病毒。它们相互作用并形成水体的微生物群落和食物网。真菌罕见悬浮生长,多作为浮游藻类、原生动物及其他生物的寄生物,有的可以聚集在固体表面形成真菌菌苔。
淡水环境中病毒能利用细菌、蓝细菌和微型藻作为它们的宿主,数量可能非常庞大,总体上能超过浮游细菌二个数量级,病毒群体密度随细菌群体波动,有的有广泛的宿主,有的为专一性宿主。病毒导致的细菌裂解能使20%~50%的细菌死亡。
原生动物是细菌和藻类的重要捕食者,因而对水体中的微生物群落有重要影响。当细菌和藻类的数量增加为原生动物提供丰富的食物来源时,这时群体增加。如食物源消耗尽,原生动物数量会急剧减少。如原生动物数量减少,细菌和藻类会再度增加。一般每个原生动物每天捕食数百个细菌和藻类,其数量一般比细菌数量少几个数量级。病毒和原生动物都有助于控制细菌和藻类群落的生物量,保持淡水生态系统中群体间的平衡。
底层生境(benthic habiat)是水体底层的沉积物层、水柱和土壤表面间的过渡区。沉积层是一个由有机物、矿物颗粒材料和水组成的疏松复合物。该区复杂的组成成分和特殊的生态环境特征,特别是氧的可利用性造成了微生物区系组成和生化反应过程的错综复杂现象。沉积层中基本营养物(C、N、S)循环和好氧微生物、厌氧微生物转化的结合如图1-8所示。富含有机物的表面微生物的氧化分解活动导致氧的消耗,产生厌氧微环境,这种微环境能支持兼性和严格的厌氧微生物的活性。沉积物中好氧-厌氧界面是异质性最大的特殊生境,这种生境支持着一个生理上最为多样的微生物群落。对C元素我们可以发现这样的转化环,发酵性细菌代谢DOM成有机酸(如乙酸和CO2),严格的厌氧细菌又可以进一步利用乙酸、CO2,并产生甲烷(CH4)。但在好氧条件下甲烷氧化菌又能利用甲烷和其他一碳化合物作为能源,再产生CO2。甲烷营养活性位于沉积物-水界面区以便利用厌氧区释放出来的CH4和水柱中的可利用氧。在沉积物-水界面上也发生N的复杂转化过程,氨化细菌使氨从有机物(沉积物)中释放出来。释放出的氨一方面被微生物同化利用,另一方面又在有氧的微生境中被自养微生物氧化生成硝酸盐,硝酸盐又可以厌氧区被反硝化产生N2释放出来。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










