



4.2.3 分子古生物学特征
4.2.3.1 生物大化石特征
生物大化石分析显示(图4.2),该泥炭岩芯主要以Sphagnum为主,特别是在深度8~9cm到30~31cm。Sphagnum的种(和亚种)主要包括S.magellanicum,S.s.acutifolia和S.s.cuspidata。其中S.magellanicum最丰富,S.s.acutifolia在深度23~24cm到32~33cm处占优势。单子叶被子植物主要出现在深度4~5cm到8~9cm和30~31cm到37~38cm。在深度13~14cm和22~23cm处出现少量的(<10%)Ericaceae。在深度6cm处出现了20%的不能鉴定的植物碎片。
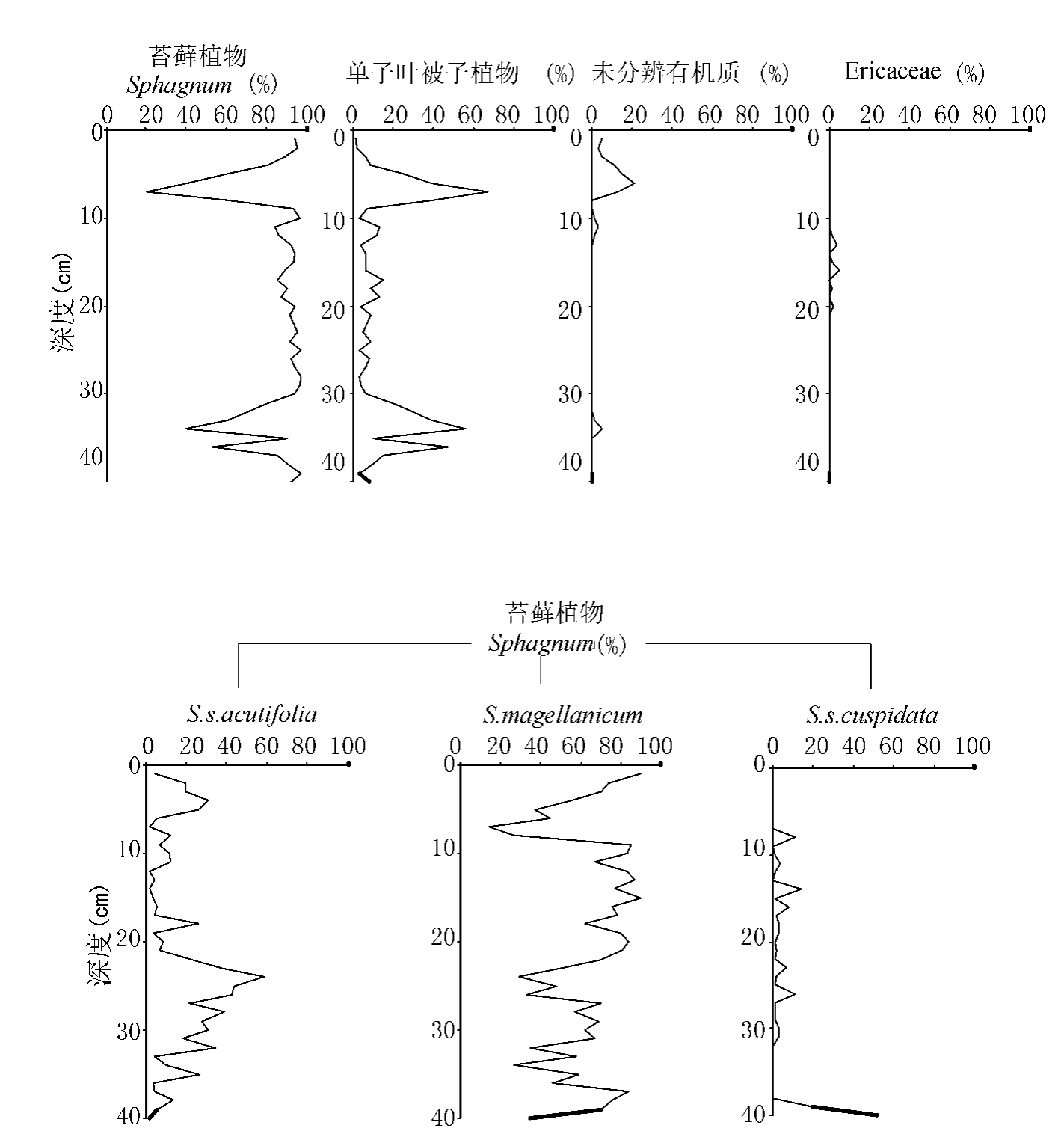
图4.2 泥炭剖面生物大化石分布
4.2.3.2 分子化石种类和分布特征
在这段40cm厚的泥炭岩芯中,大部分分子化石的分布(包括酸、醇、酮、酯等)没有多大的变化,但正构烷烃的分布及其单体碳、氢同位素表现出明显的规律性变化,可以用来划分乃至对比本区的泥炭地层。下面首先介绍一下检测出的分子化石特征。
1.烃类 正构烷烃碳数分布为C21~C35,以C23和/或C31为主峰,并具有很明显的奇偶优势(图4.3),这种分布形式主要与泥炭植物的蜡有关。在整个岩芯中,最明显的变化特征是其主峰化合物的变化,即C23与C31的相互取代。低碳数正构烷烃(C14~C20)占总正构烷烃的含量不到3%,它们可能来源于细菌(Otto等,1994)和藻类(Blumer等,1971)。
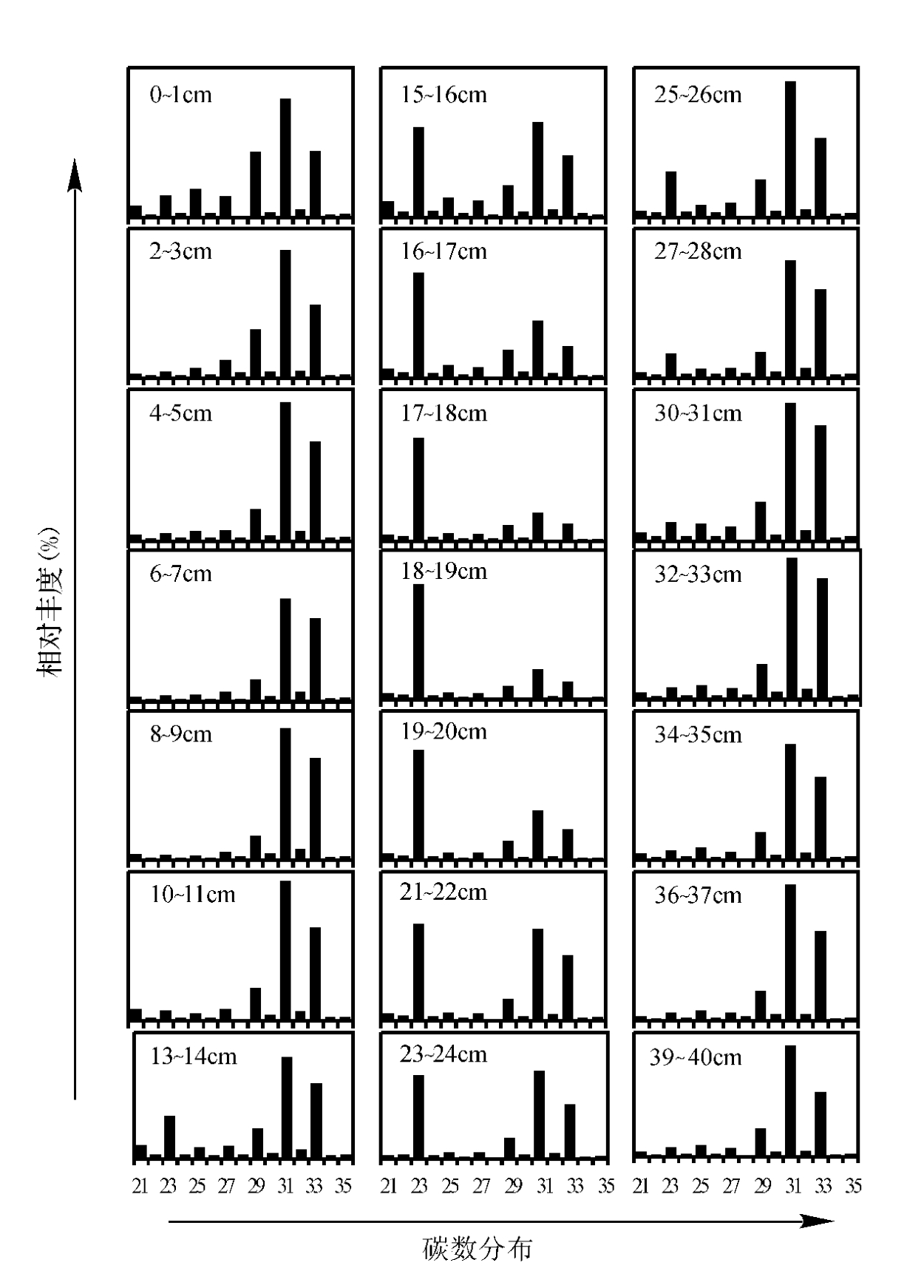
图4.3 不同深度正构烷烃的分布
与正构烷烃一起,还有一小部分正构烯烃被检测出来,碳数为C19~C29。长链正构烯烃曾在湖相沉积物(Giger和Schaffner,1977)、南极土壤(Matsumoto等,1990)和其他泥炭(Dehmer,1993)中出现,它们可能来源于维管植物、微观藻类和蓝细菌(Dehmer,1993)。
三萜类化合物在泥炭类脂物中很少出现,一旦出现往往是一些不饱和化合物(Dehmer,1993)。在本泥炭中出现的非藿烷结构的三萜烯化合物主要是蒲公英-14-烯(taraxer-14-ene)和蒲公英-20-烯(taraxast-20-ene),还有少量的奥利-12-烯和乌散-12-烯。以前曾报道在泥炭、褐煤和海相沉积物中出现蒲公英-14-烯(Brassell等,1980)。它的可能前身物,如蒲公英-14-烯-3-醇和蒲公英-14-烯-3-酮也曾在许多海相沉积物中发现过(Brassell,1980;Brassell和Eglinton,1983;Volkman等,1987)。这些非藿烷结构的三萜烯化合物主要来源于泥炭中的高等植物,它们可能的生物先体,如蒲公英醇(taraxerol)在植物体中分布很广,这个C30的醇类被证实是Sphagnum spp.植物中普遍存在的一种三萜类化合物。
本泥炭中检测出来的三萜烷主要是C31αβ-29-甲基藿烷和少量的C31 ββ-29-甲基藿烷。17β(H),21β(H)构型的藿烷常被认为与细菌有关(Ourisson等,1979),通过对藿酸和藿多醇的还原而形成。αβ这种地质构型的藿烷被认为是由ββ生物构型藿烷的异构化形成的,但目前还难以解释它们出现在没有明显热成熟作用的环境中。以前曾经报道过C31αβ藿烷出现在泥炭中(Quirk等,1984;van Dorsselaer等,1977),甚至在泥炭腐殖化作用的最早期出现。这个化合物在本泥炭岩芯最表层的样品中出现。曾认为17α(H),21β(H)-一升藿烷是由于在酸性的泥炭环境中,通过对细菌藿四醇的氧化和随后的脱羧反应,并同时由于细菌作用在C-17位的异构化而形成的(van Dorsselaer等,1977;Ries-Kautt和Albrecht,1989)。这种异构化作用在泥炭的酸性环境中是很容易实现的。本泥炭岩芯该化合物的单体碳同位素特征也支持这个化合物来源于微生物。
2.酮和酯 本泥炭检测出的α-正构脂肪酮碳数分布为C25~C35,主峰在C29或C31,有很强的奇碳优势。该组分中的三萜类化合物主要是蒲公英酮(taraxerone)和奥利-13(18)-烯-3-酮。
Morison和Bick(1967)曾报道过泥炭中的α-正构脂肪酮,碳数为C17~C35,有很强的奇碳优势,主峰主要是C25和C27。在一个Sphagnum泥炭中,Lehtonen和Ketola(1990)发现了C17~C35α-正构脂肪酮,但主峰化合物为C17,而且低碳数分子(C17~C23)丰度随腐殖化程度的增强而升高。本泥炭中缺少这些低碳数分子,可能与所分析的岩芯因埋藏浅而腐殖化程度较低有关。
泥炭中α-正构脂肪酮有多种可能的来源:①泥炭植物的表皮蜡;②正构烷烃的微生物氧化;③正构脂肪酸的β-氧化和脱羧作用;④长链α-正构脂肪酮向短链的转化。本泥炭中高丰度的C29和C31α-正构脂肪酮可能来源于正构烷烃的氧化以及泥炭植物的蜡,而不太可能来源于正构脂肪酸,因为后者的主峰化合物为C26和C24,而对应的C25和C23α-正构脂肪酮的含量很低。
酯化合物主要是C16脂肪酸酯,碳数分布为C38~C52,主峰化合物为C40 和C42(15~16cm深度以下)或C48和C50(15~16cm深度以上),有很明显的偶碳优势。
3.醇和甾醇检测出最主要的甾烯醇和甾烷醇主要有:24-乙基胆甾-5-烯-3β-醇,24-乙基-5α-胆甾-3β-醇,24-乙基胆甾-5,22-二烯-3β-醇,24-乙基-5α-胆甾-22-烯-3β-醇,24-甲基胆甾-5-烯-3β-醇和24-甲基-5α-胆甾-3β-醇。甾醇的分布反映了泥炭有机质主要来源于高等植物。
正构脂肪醇碳数为C20~C34,有很明显的偶碳优势,C22为主峰化合物。与正构烷烃相似,这些化合物相对较难受到生物的改造(Ficken等,1998),然而,它们在本泥炭岩芯中的分布还是与生物大化石不吻合,酯的水解作用可以释放出一些正构脂肪醇,这可能使后者的分布复杂化了。
4.酸酸的组分很复杂,但主要是正构脂肪酸,α,ω-二元羧酸和ω-羟基酸。正构脂肪酸碳数分布为C16~C34,主峰为C24或C26,具很明显的偶碳优势。最浅部样品出现了两个主峰(C16和C24)的双峰型分布,前主峰C16的丰度随深度变化很快降低。但总的来说,脂肪酸在整个岩芯中变化不大。
α,ω-二元羧酸碳数分布为C20~C28,主峰为C22,有明显的偶碳优势。其分布在整个岩芯中几乎很稳定,但在16~18cm深度,C22,C24和C26表现出相近的丰度。
ω-羟基脂肪酸碳数分布为C20~C28,主峰为C22或C26,有明显偶碳优势。一个比较特征的分布是高碳数ω-羟基脂肪酸,尤其是C26,在13~14cm以下深度,其丰度明显增加。曾在一个高度腐殖化的Sphagnum泥炭中报道过类似的C26ω-羟基脂肪酸丰度增加的现象(Lehtonen和Ketola,1993)。
脂肪酸在生物体中广泛分布,能够完成许多功能。羟基脂肪酸是在高等植物中分布最广泛的脂肪酸,但在真菌中很稀少。高碳数的ω-羟基脂肪酸(>C20)是高等植物的标志(Killops和Killops,1993)。α,ω-二元羧酸可能主要来源于泥炭高等植物,它们在泥炭植物中以聚合物酯的形式存在(Eglinton和Hamilton,1967;Karunen等,1983;Lehtonen和Ketola,1993)。然而,由于脂肪酸在沉积环境中容易受到微生物的改造,在利用其分布来判断生物源时应谨慎。例如,ω-羟基脂肪酸可以经微生物作用,转化成α,ω-二元羧酸。在本泥炭中,ω-羟基脂肪酸和α,ω-二元羧酸具有相似的碳数分布,以及在表层样品中,α,ω-二元羧酸的CPI值很低,这些都说明了一部分α,ω-二元羧酸可能由微生物氧化ω-羟基脂肪酸而形成。
4.2.3.3 单体分子化石的碳同位素组成
稳定同位素被广泛认为是重要的气候替代指标。由于有关CO2进入植
物叶子和随后的光合作用过程中的同位素分馏效应有很好的理论基础(Farquhar等,1989),加上有很多实验确定了环境因子对植物体δ13C的影响(Madhavan等,1991;Broadmeadow和Griffiths,1993;Saurer等,1995),植物有机体的稳定碳同位素比值被广泛用来探讨生物生长的环境条件,如温度和大气CO2含量等。利用气相色谱-燃烧-同位素比质谱仪(GC-CIRMS)测出的单体分子化石的δ13C值是一个综合的信息载体,它们反映了生物所利用的碳源的同位素值以及碳固定和生物合成过程中的同位素分馏效应,相应地,它们记录了环境的信息(Hayes,1990;Ficken等,1998)。
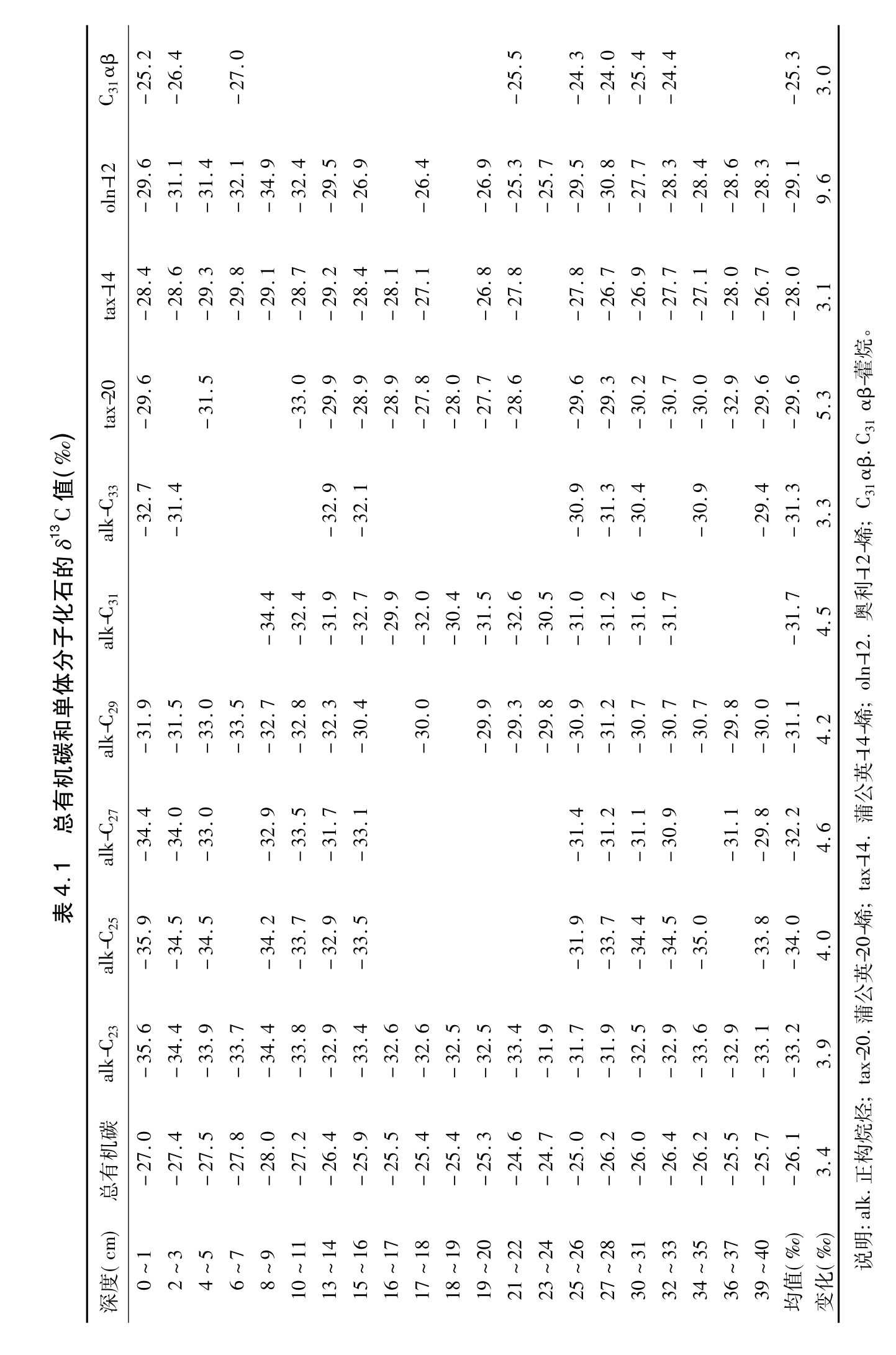
1.总有机碳的δ13C总有机碳的δ13C值变化于-24.6‰~-28.1‰(表4.1),显示了一种C3植物源特征(Galimov,1985)。在22~23cm深度以上(210Pb年龄为公元1870年),总的趋势是其值越来越小(-24.6‰~-28.0‰)。已经证实,工业革命以来(130年),由于化石燃料的燃烧,大气CO2的δ13C值约降低1.3‰,即从工业革命以前的-6.5‰变到现在的-7.8‰(Friedli等,1986)。本泥炭剖面的总有机碳的δ13C值在该段时间内,比工业革命的影响值还低2.2‰,这可能与温度变化有关,下文将要讨论到这一点。
2.单体烃类的δ13C表4.1还列出了本泥炭岩芯一些特定类脂物的单体
δ13C值。所有化合物都比总有机碳的δ13C更小。这与前人报道的类脂物单体碳同位素值比C3植物总有机质要低是一致的(Rieley等,1993;Collister等,1994)。C23和C29正构烷烃的δ13C值分别变化于-34.2‰~-32.5‰和-33.5‰~-29.3‰。C29正构烷烃比C23更富(2.1‰)13C,这与现代Sphagnum植物体的分析结果是一致的(Ficken等,1998;Nott等,2000)。所有正构烷烃比非藿烷结构的三萜烯化合物更缺乏13C。这些三萜烯化合物在本剖面中的δ13C平均值为-29.9‰,大致比C29,C31和C33正构烷烃的低1.5‰。这与Hayes(1993)的研究结果相吻合,他认为由于它们各自生物先体的同位素差异,萜类化合物比直链状类脂物的δ13C值约低1.5‰。
正构烷烃、三萜烯和藿烷化合物的δ13C值总的变化幅度为3.0‰~9.6‰(表4.1)。其中1.3‰的变化与工业革命以来的化石燃料的燃烧有关,这在C23正构烷烃上很明显地反映出来,其δ13C值从工业革命以来出现逐渐降低的变化趋势。
在泥炭表层样品中,C23正构烷烃的δ13C值(-35.5‰)比现代泥炭的Sphagnum植物体的C23正构烷烃的δ13C值(-37.6‰)要高2.1‰。曾经报道过,泥炭表层样品正构烷烃的δ13C要比生物体中的高2.5‰(Ficken等,1998),植物叶子和土壤中的正构烷烃的δ13C值也相差1‰(Lichtfouse等,1995)。这可能与成岩作用有关,它使更富12C的CO2和CH4散发掉。在近地面的大气中曾经观察到由于地面有机质的腐烂和氧化,其CO2含有更多的12C(Keeling,1961;Münnich和Vogel,1959)。
前人曾经报道过在泥炭中出现大量的C31αβ藿烷(Quirk等,1984;van Dorsselaer等,1977),但有关它的来源问题还远未研究清楚。本泥炭的藿烷化合物只要含量足够高,都作了单体碳同位素分析。在8个样品中C31-17α(H),21β(H)-29-甲基藿烷的δ13C值变化于-24.0‰~-27.0‰,平均为-25.3‰(表4.1)。与非藿烷结构的三萜烯相比(δ13C均值为-29.9‰),C31 17α(H),21β(H)-29-甲基藿烷要富4.6‰的13C,这可能指示了这种藿烷来源于微生物。曾经在一些土壤剖面中观察到藿-22(29)-烯或藿-13(18)-烯比高等植物的正构烷烃要富4‰~5‰的13C(Huang等,1996),这种差异是由异养细菌或蓝细菌造成的。曾经报道过碳水化合物和蛋白质比类脂物要富5‰的13C(Deines,1980),这与本文C31-17α(H),21β(H)-29-甲基藿烷要比三萜烯富4.6‰的数值比较接近。这可能暗示,泥炭表面的微生物可能以碳水化合物和蛋白质为主要碳源,因为随后的细菌生物合成作用造成的碳同位素分馏效应较小。以前曾报道过,土壤异养细菌能够利用这种碳源(Huang等,1996)。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












