



第1节 染色体畸变
染色体畸变(chromosome aberration)是指体细胞或生殖细胞内染色体发生了异常改变。在配子形成期或合子期(约受精后24小时内)发生的染色体畸变,将导致个体全身所有的细胞均带有畸变的染色体,即形成各种常见的单体型、三体型、部分单体型、部分三体型等染色体病患者;在胚胎早期的卵裂及桑椹胚期(受精后3~4天内)发生的染色体畸变,将导致个体发育成含有不同百分比核型的染色体病嵌合体患者;其后由于3个胚层的分化,某一胚层发生的染色体畸变,将累及由该胚层发育而来的相关器官系统。胎儿出生后,在各种内外环境因素的影响下,某组织的体细胞发生突变,通过有丝分裂,它可将该突变传给其子细胞而形成克隆,有可能导致相应部位的肿瘤发生。
染色体畸变可分为数目畸变和结构畸变两大类,其中数目畸变又可分为整倍性改变和非整倍性改变。结构畸变主要有缺失、重复、插入、易位和倒位等。当一个个体的细胞有两种或两种以上不同核型的细胞系时,这个个体就被称为嵌合体(mosaic)。无论数目畸变,还是结构畸变,其实质是涉及染色体或其某节段上基因群的增减或位置的转移,使遗传物质发生改变,导致染色体异常综合征,或称染色体病。
一、染色体畸变发生的原因
染色体畸变可以自发地产生,称为自发畸变(spontaneous aberration);也可通过物理的、化学的和生物的诱变作用而产生,称为诱发畸变(induced aberration);还可由亲代遗传而来。造成染色体畸变的原因是多方面的,主要包括化学因素、物理因素和生物因素,这些因素与导致基因突变的因素基本相同。一般而言,引起染色体畸变的剂量或能量要大大超过引起基因突变的剂量或能量。
(一)化学因素
1.药物
某些药物特别是一些抗肿瘤药物、保胎及预防妊娠反应的药物均可引起人类染色体畸变或产生畸胎。如环磷酰胺、氮芥、白硝安(马利兰)、氨甲喋呤、阿糖胞苷等抗癌药物可导致染色体畸变;抗痉挛药物苯妥英钠可引起人淋巴细胞产生多倍体细胞。
2.农药
许多化学合成的农药可以引起人类细胞染色体畸变。如农药中的除草剂、杀虫的砷制剂等都是染色体畸变的诱变剂;敌百虫类农药、有机磷农药,可使染色体畸变率增高。
3.工业毒物
工业毒物如苯、甲苯、铝、砷、二硫化碳、氯丁二稀、氯乙烯单体等,都可以导致染色体畸变。长期接触这些有害毒物的工人,其染色体的畸变率比一般群体要高。
4.食品添加剂
某些食品的防腐剂和色素等添加剂中所含的化学物质也可以使人类染色体发生畸变,如硝基呋喃基糖酰胺AF-2、环已基糖精等。
(二)物理因素
在大自然空间存在的各种各样的射线可对人体产生一定的影响,但其剂量极微,因此影响不大。但大量的电离辐射对人类具有极大的潜在危险。例如放射性物质爆炸后散落的放射性尘埃、医疗上所用的放射线等,对人体都有一定的损害。工业放射性物质的污染也可引起细胞染色体的改变。细胞受到电离辐射后,可引起细胞内染色体发生异常,畸变率随射线剂量的增高而增高。最常见的畸变类型有断裂、缺失、双着丝粒染色体、易位、核内复制等,这些畸变都可使个体出现临床症状。射线既可影响体细胞又可影响生殖细胞,如果一次照射大剂量的射线,可在短期内引起造血障碍而导致个体死亡。长期接受射线治疗或从事放射工作的人员,由于微小剂量的射线不断积累,也会引起体细胞或生殖细胞染色体畸变。有实验证明,受照射的卵细胞,其染色体不分离的频率明显高于未受照射组。同时还发现,这一现象在年龄较大的小鼠中更为明显。还有报道指出,受到过电离辐射的母亲生育唐氏综合征患儿的风险明显增高。
(三)生物因素
导致染色体畸变的生物因素包括两个方面。一是由生物体产生的生物类毒素所致;二是某些生物体如病毒本身可引起染色体畸变。霉菌毒素具有一定的致癌作用,同时也可引起细胞内染色体畸变,如杂色曲霉素、黄曲霉素、棒曲霉素等;病毒也可引起宿主细胞染色体畸变,尤其是那些致癌病毒,其原因主要是由于影响DNA代谢;当人体感染某些病毒,如风疹病毒、乙肝病毒、麻疹病毒和巨细胞病毒时,就有可能引发染色体的畸变。如果用病毒感染体外培养细胞将会出现各种类型的染色体异常。
(四)母亲年龄
高龄孕妇所生子女,其细胞中某一序号染色体有三条的情况要多于一般孕妇。母亲年龄越大(大于35岁),生育唐氏综合征患儿的危险性就越高。母亲生育年龄只是环境因子在体内累积作用的表现形式,这与生殖细胞老化及合子早期所处的宫内环境有关。一般认为,生殖细胞在母体内停留的时间越长,受到各种因素影响的机会越多,在以后的减数分裂过程中,容易产生染色体不分离,导致染色体数目异常。
二、染色体数目异常及其产生机制
人体正常生殖细胞(精子或卵子)所包含的全部染色体称为一个染色体组(chromosome set),因此,精子和卵子为单倍体(haploid),以n表示,分别含有22条常染色体和1条性染色体(X或Y);受精卵为二倍体(diploid),以2n表示,包括22对常染色体和1对性染色体(XX或XY)。以人类二倍体数目为标准,如果体细胞的染色体数目(整组或整条)增加或减少,则称为染色体数目畸变。包括整倍体改变和非整倍体改变两种形式。
(一)整倍体改变
如果染色体的数目变化是以一个染色体组(n)为基数,整倍地增加或减少,则称为整倍体(euploid)改变,超过二倍体的整倍体,被称为多倍体(polyploid)。
在2n的基础上,如果增加一个n,则染色体数为3n,即三倍体(triploid);若在2n的基础上增加两个n,则为4n,即四倍体(tetraploid);以此类推。如果在2n的基础上减少一个染色体组,则称为单倍体。
在人类中已知有三倍体和四倍体的胎儿,但只有极少数三倍体(图7-1)的胎儿能存活到出生,存活者多为2n/3n的嵌合体。在流产的胎儿中三倍体是常见的类型。一般认为,三倍体胎儿易于流产的原因是在胚胎发育过程中,细胞有丝分裂形成三极纺锤体,因而造成染色体在细胞分裂中后期时的分布和分配紊乱,最终导致子细胞中染色体数目异常,从而严重干扰了胚胎的正常发育而导致流产。四倍体比三倍体罕见,往往是四倍体和二倍体(4n/2n)的嵌合体,活婴罕见,在流产的胚胎中可见。单倍体和四倍体以上的多倍体个体尚未见报道。
整倍体改变的机制主要有双雌受精、双雄受精、核内复制和核内有丝分裂。
1.双雄受精
一个正常的卵子同时与两个正常精子受精称为双雄受精(diandry)。由于每个精子带有一个染色体组,所以当两个精子同时进入一个卵细胞时,就将两个染色体组同时带入了这一卵细胞,所形成的合子则含有三个染色体组(三倍体),可形成69,XXX,或69,XXY,或69,XYY三种类型的受精卵(图7-2)。
图7-1 三倍体核型
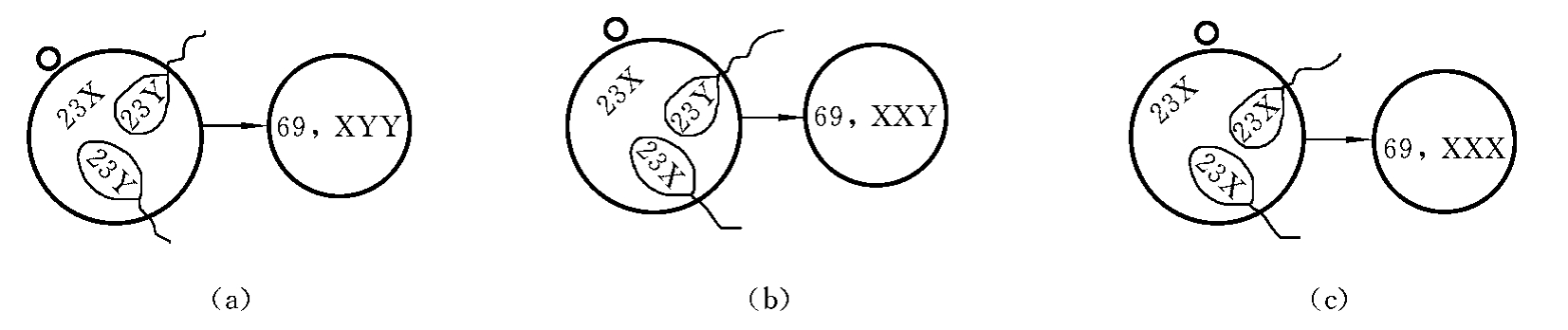
图7-2 双雄受精
2.双雌受精
一个二倍体的异常卵子与一个正常的精子受精,从而产生一个三倍体的合子,称为双雌受精(digyny)。在卵细胞发生的第二次减数分裂过程中,次级卵母细胞由于某种原因未形成第二极体,因此,应分给第二极体的染色体组仍留在卵细胞中,使该卵细胞成为异常卵细胞。当它与一个正常的精子结合后,就将形成含有三个染色体组的合子(三倍体),可形成69,XXX或69,XXY两种核型的受精卵(图7-3)。
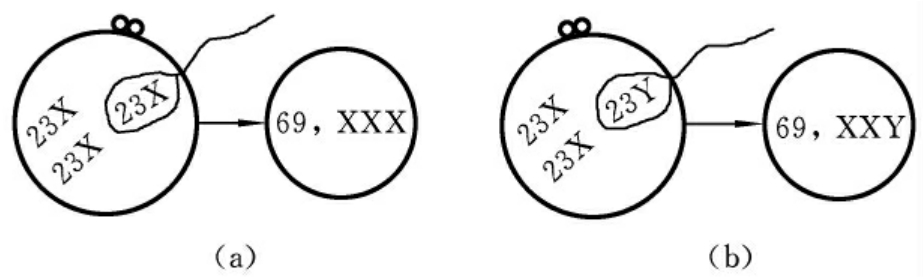
图7-3 双雌受精
3.核内复制
核内复制(endoreduplication)是指在一次细胞分裂时,DNA不是复制一次,而是复制了两次,而细胞只分裂了一次。这样形成的两个子细胞都是四倍体,这是肿瘤细胞常见的染色体异常特征之一(图7-4)。
4.核内有丝分裂
核内有丝分裂(endomitosis)是指在正常的细胞分裂时,染色体正常复制了一次,但到分裂中期时,核膜仍未破裂,也无纺锤体的形成,因此,细胞分裂未能进入后期和末期,没有细胞质的分裂,结果细胞内含有四个染色体组,形成了四倍体。
归纳来说,三倍体的形成原因可为双雌受精或双雄受精;四倍体形成的主要原因是核内复制或核内有丝分裂。

图7-4 核内复制
(二)非整倍体改变
如果体细胞中的染色体不是整倍数,而是增加或减少了一条或数条,则称为非整倍体(aneupliod),这是临床上最常见的染色体畸变类型,包括超二倍体(hyperdiploid)和亚二倍体(hypodiploid)。
1.亚二倍体
当体细胞中染色体数目少了一条或数条时,称为亚二倍体,即在2n的基础上,减少了一条或几条染色体,可写做2n-m(其中m<n)。若某对染色体少了一条(2n-1),细胞染色体数目为45,即构成单体型(monosomy)。临床上常见的有21号、22号和X染色体的单体型,核型为45,XX(或XY),-21;45,XX(或XY),-22和45,X。核型为45,X的个体往往是由于X染色体的丢失所致,具有这种核型的个体,多在胚胎期流产,少数存活。由于缺少一条X染色体,具有性腺发育不全等临床症状。
2.超二倍体
当体细胞中染色体数目多了一条或数条时,称为超二倍体。在超二倍体的细胞中某一同源染色体的数目不是2条,而是3条、4条……。
若某对染色体多了一条(2n+1),细胞内染色体数目为47,即构成三体型(trisomy),这是人类染色体数目畸变中最常见、种类最多的一类畸变。例如,在常染色体病中,除了第17号染色体尚未有三体型的病例报道外,其余的染色体三体型均有报道。少数常染色体三体型病例可以存活至出生,甚至可以活至成年,这表明人类增加一条额外的常染色体显然比少一条常染色体的临床症状要轻。但是由于染色体的增加,特别是较大染色体的增加,将造成基因组的严重失衡而破坏或干扰胚胎的正常发育,故绝大部分常染色体三体型核型只见于早期流产的胚胎。少数三体型病例可以存活至出生,但多数寿命不长,并伴有各种严重畸形。
三体型以上的统称为多体型(polysomy)。多体型常见于性染色体中,如性染色体四体型(48,XXXX;48,XXXY;48,XXYY)和五体型(49,XXXXX;49,XXXYY)等。如果患者细胞中一对同源染色体同时缺失,即减少了一对同源染色体(2n-2),称为缺体型(nullosomy),人类缺体型目前尚未见报道。
(三)非整倍体的产生机制
多数非整倍体的产生原因是生殖细胞在减数分裂过程或受精卵早期卵裂中,发生了染色体不分离或染色体丢失。
1.染色体不分离
在细胞进入分裂中后期时,如果某一对同源染色体或姐妹染色单体彼此没有分离,而是同时进入一个子细胞,结果所形成的两个子细胞中,一个将因染色体数目增多而成为超二倍体,另一个则因染色体数目减少而成为亚二倍体,这个过程称为染色体不分离(non-disjunction)。
染色体不分离可发生于配子形成时的减数分裂过程中,称减数分裂不分离(meiotic non-disjunction);也可发生于受精卵的卵裂早期或在体细胞的有丝分裂过程中,称有丝分裂不分离(mitotic non-disjunction)。
(1)减数分裂不分离。染色体不分离发生在第一次减数分裂,使得某一对同源染色体不分离,同时进入一个子细胞核,所形成的配子中,一半将有24条染色体(n+1),另一半将有22条(n-1)。与正常配子受精后,将形成超二倍体或亚二倍体。若在第二次减数分裂时发生染色体不分离,所形成的配子的染色体数将有以下几种情况:1/2为n、1/4为(n+1)、1/4为(n-1)。它们与正常配子受精后,得到相应的二倍体、超二倍体、亚二倍体(图7-5)。
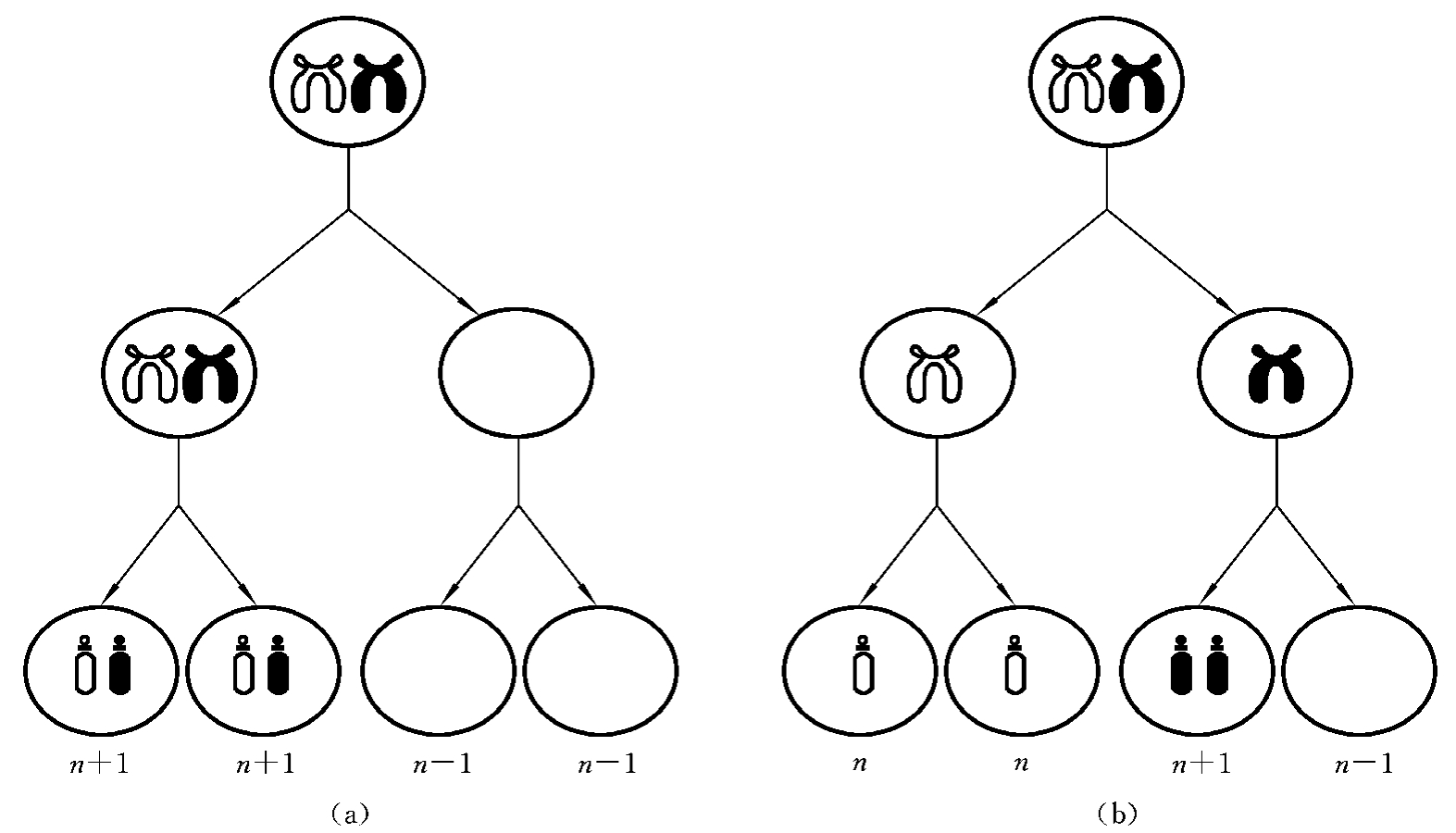
图7-5 减数分裂中染色体不分离
(a)同源染色体不分离 (b)姐妹染色单体不分离
(2)有丝分裂不分离。卵裂早期,某一染色体的姐妹染色单体不分离,可导致产生由两种细胞系或三种细胞系组成的嵌合体。染色体不分离发生在第一次卵裂时,则形成具有两个细胞系的嵌合体,一个为超二倍体细胞系,一个为亚二倍体细胞系。染色体不分离发生在第二次卵裂(图7-6)时,即形成具有三个或三个以上细胞系的嵌合体(46/47/45)。染色体不分离发生得越晚,正常二倍体细胞系的比例越大,临床症状也相对较轻。
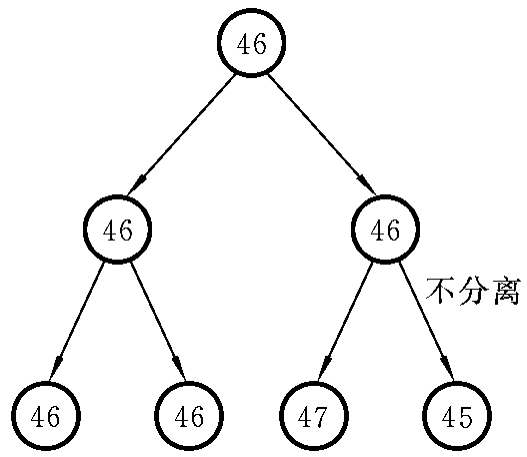
图7-6 第二次卵裂时染色体不分离
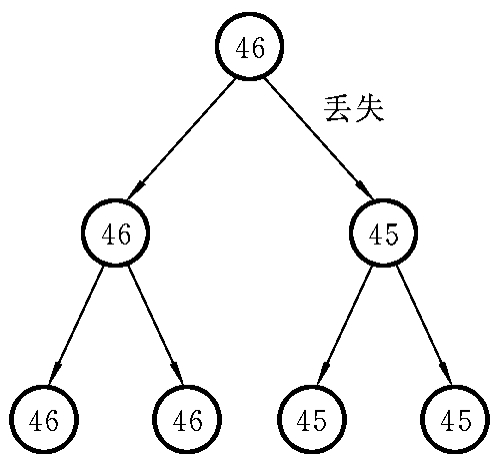
图7-7 染色体丢失与嵌合体的形成
2.染色体丢失
染色体丢失(chromosome lose)又称染色体分裂后期延滞(anaphase lag),在细胞有丝分裂过程中,某一染色体未与纺锤丝相连,不能移向两极参与新细胞的形成;或者在移向两极时行动迟缓,滞留在细胞质中,最后分解、消失,造成该条染色体的丢失而形成亚二倍体。染色体丢失也是嵌合体形成的一种方式(图7-7)。
(四)假二倍体
有时细胞中某些染色体数目发生了异常,其中有的增加,有的减少,而增加和减少的染色体数目相等,结果染色体总数不变,还是二倍体数(46条),但不是正常的二倍体核型,则称为假二倍体(pseudodiploid)。
按照人类细胞遗传学命名的国际体制(ISCN),非整倍体的描述方法为“染色体总数,性染色体组成,+(-)畸变染色体序号”。例如:某一核型中的18号染色体多了一条,可描述为47,XX(或XY),+18;少了一条22号染色体则描述为45,XX(或XY),-22;若是少了一条X染色体,可描述为45,X。
三、染色体结构畸变及其产生机制
导致染色体结构畸变(structural aberration)的基础是染色体发生断裂(breakage)及断裂后的异常重接(rejoin)。如果某条染色体受射线等因素影响后发生了断裂,但随后在原位重接,将不引起遗传效应。如果染色体发生断裂后,未发生重接或未原位重接,这就将引起各种染色体结构畸变,称染色体重排。发生了结构畸变的染色体即称重排染色体(rearrangement chromosome)。
(一)染色体结构畸变的描述方法
人类细胞遗传学命名的国际体制(ISCN)制定了有关人类染色体以及染色体畸变等的命名方法。结构畸变染色体核型的描述方法有简式和详式两种。①简式。对染色体结构的改变只用其断裂点来表示。按国际命名规定,应依次写明染色体总数,性染色体组成,然后用一个字母(如t)或三联字符号(如del)写明重排染色体的类型,其后的第一个括弧内写明染色体的序号,第二个括弧内写明区号、带号以表示断裂点。②详式。除了简式中应写明的内容外,与简式有所不同的是,在最后一个括弧中不是只描述断裂点,而是描述重排染色体带的组成。涉及两条以上染色体异常时,先描述号数小的染色体,若涉及性染色体则先描述性染色体。一般由短臂末端向长臂末端描述,除非号数小的染色体短臂异常。
(二)染色体结构畸变的类型及其产生机制
临床上常见的染色体结构畸变有:缺失、重复、易位、倒位、环状染色体和等臂染色体等。染色体结构畸变只有在显带标本上才能准确识别。
1.缺失
缺失(deletion)是染色体片段的丢失,缺失使位于这个片段的基因也随之丢失。按染色体断裂点的数量和位置,缺失可分为末端缺失和中间缺失两类。
(1)末端缺失(terminal deletion)。指染色体的臂发生断裂后,未发生重接,无着丝粒的片段不能与纺锤丝相连而丢失。如图7-8(a)所示,第1号染色体长臂的2区1带发生断裂,其远侧端(q21→qter)丢失。这条染色体是由短臂的末端至长臂的2区1带所构成。这种结构畸变的简式描述为46,XX(或XY),del(1)(q21);详式描述为46,XX(或XY),del(1)(pter→q21:)。

图7-8 缺失
(a)末端缺失 (b)中间缺失
(2)中间缺失(interstitial deletion)。指一条染色体的同一臂上发生了两次断裂,两个断裂点之间的片段丢失,其余的两个断片重接。如图7-8(b)所示,3号染色体长臂上的q21和q31发生断裂和重接,这两断裂点之间的片段丢失。这种结构畸变的简式描述为46,XX(或XY),del(3)(q21q31);详式描述为46,XX(或XY),del(3)(pter→q21∷q31→qter)。
丢失的片段大小不同将产生不同的生物学效应。大片段的缺失甚至在杂合状态下也是致死的,如果缺失的部分包括某些显性基因,则同源染色体上与这一缺失相对应位置上的隐性等位基因就得以表现,这一现象称为假显性。
2.重复
重复(duplication)是一个染色体上某一片段增加了一份以上的现象,使这些片段的基因多了一份或几份。原因是同源染色体之间的不等交换或染色单体之间的不等交换以及染色体片段的插入等。重复的分子细胞效应比缺失缓和,但如果重复片段较大也会影响个体的生活能力,甚至出现死亡。
3.倒位
倒位(inversion)是某一染色体发生两次断裂后,两断裂点之间的片段旋转180°后重接,造成染色体上基因重排。染色体的倒位可以发生在同一臂(长臂或短臂)内,也可以发生在两臂之间,分别称为臂内倒位和臂间倒位。
(1)臂内倒位(paracentric inversion)。一条染色体的某一臂上同时发生了两次断裂,两断裂点之间的片段旋转180°后重接。例如1号染色体p22和p34同时发生了断裂,两断裂点之间的片段倒转后重接,形成了一条臂内倒位的染色体(图7-9(a))。这种结构畸变的简式描述为46,XX(或XY),inv(1)(p22p34);详式描述为46,XX(或XY),inv(1)(pter→p34∷p22→p34∷p22→qter)。
(2)臂间倒位(pericentric inversion)。一条染色体的长、短臂各发生了一次断裂,中间断片颠倒后重接,则形成了一条臂间倒位染色体。如4号染色体的p15和q21同时发生了断裂,两断裂点之间的片段倒转后重接,形成了一条臂间倒位染色体(图7-9(b))。这种结构畸变的简式描述为46,XX(或XY),inv(4)(p15q21);详式描述为46,XX(或XY),inv(4)(pter→p15∷q21→p15∷q21→qter)。
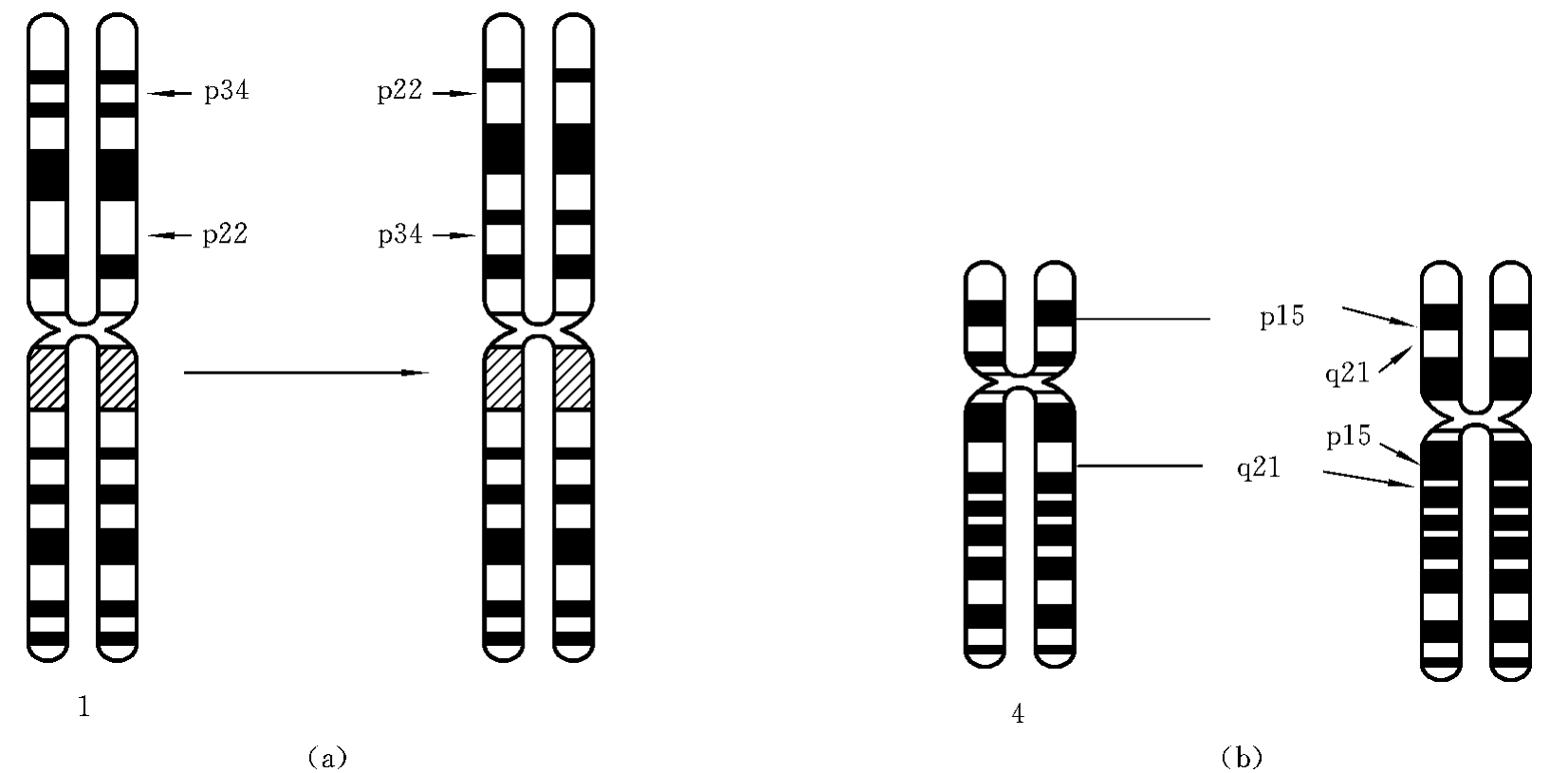
图7-9 倒位
(a)臂间倒位 (b)臂内倒位
倒位染色体在减数分裂同源染色体联会时,如倒位片段很小,倒位片段可能不发生配对,其余区段配对正常;如倒位片段很长,倒位的染色体可能倒过来和正常的染色体配对,形成一个环,称为倒位环(inversion loop)。环内两条非姐妹染色单体间发生单体交换,形成4种类型的染色体。
4.易位
一条染色体的断片移接到另一条非同源染色体的臂上,这种结构畸变称为易位(translo-cation)。常见的易位方式有相互易位、罗伯逊易位和插入易位等。
(1)相互易位(reciprocal translocation)。相互易位是两条染色体同时发生断裂,断片交换位置后重接,形成两条衍生染色体(derivation chromosome)。当相互易位仅涉及位置的改变而不造成染色体片段的增减时,称为平衡易位。如2号染色体长臂2区1带和5号染色体长臂3区1带同时发生了断裂,两断片交换位置后重接,形成两条衍生染色体。这种结构畸变的简式描述为46,XX(或XY),t(2;5)(q21;q31);详式描述为46,XX(或XY),t(2;5)(2pter→2q21∷5q31→5qter;5pter→5q31∷2q21→2qter)(图7-10)。相互易位的杂合体在减数分裂粗线期,由于同源部分的联会配对而形成特征性的四射体(图7-11)。
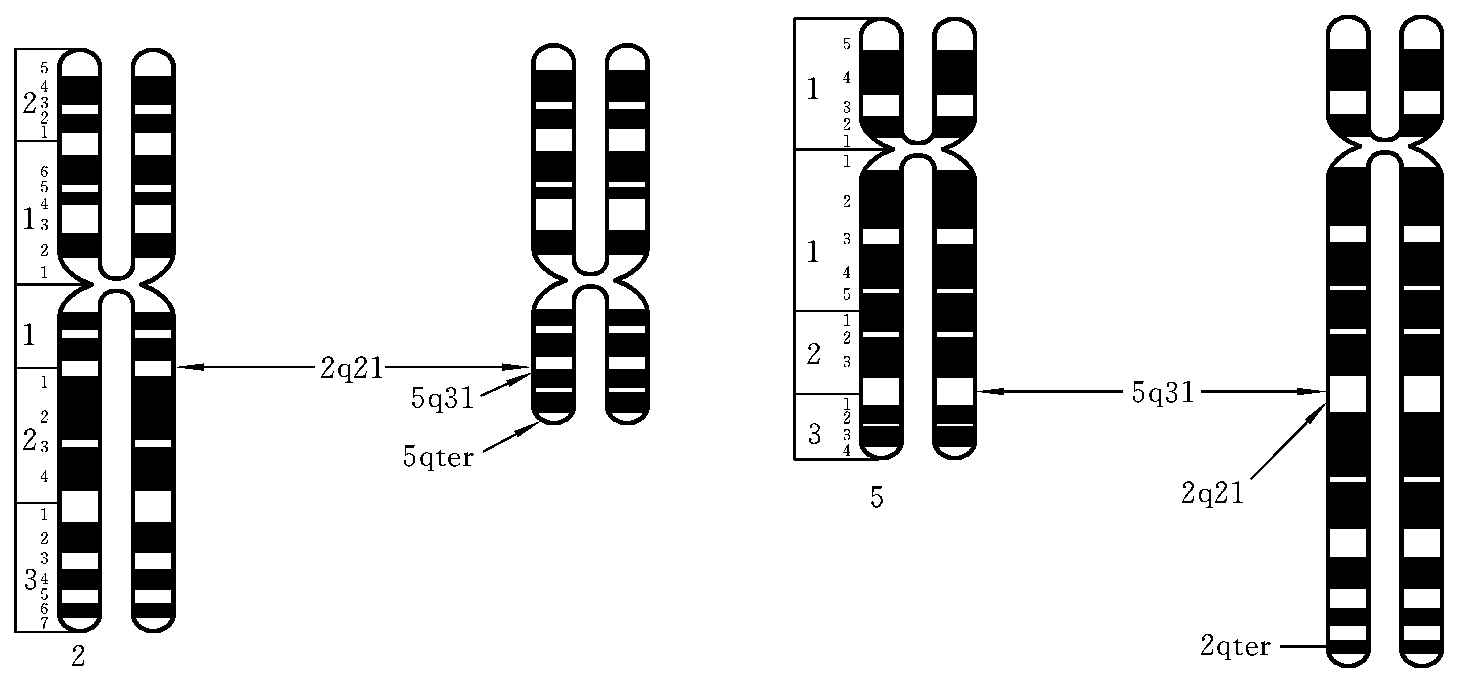
图7-10 相互易位
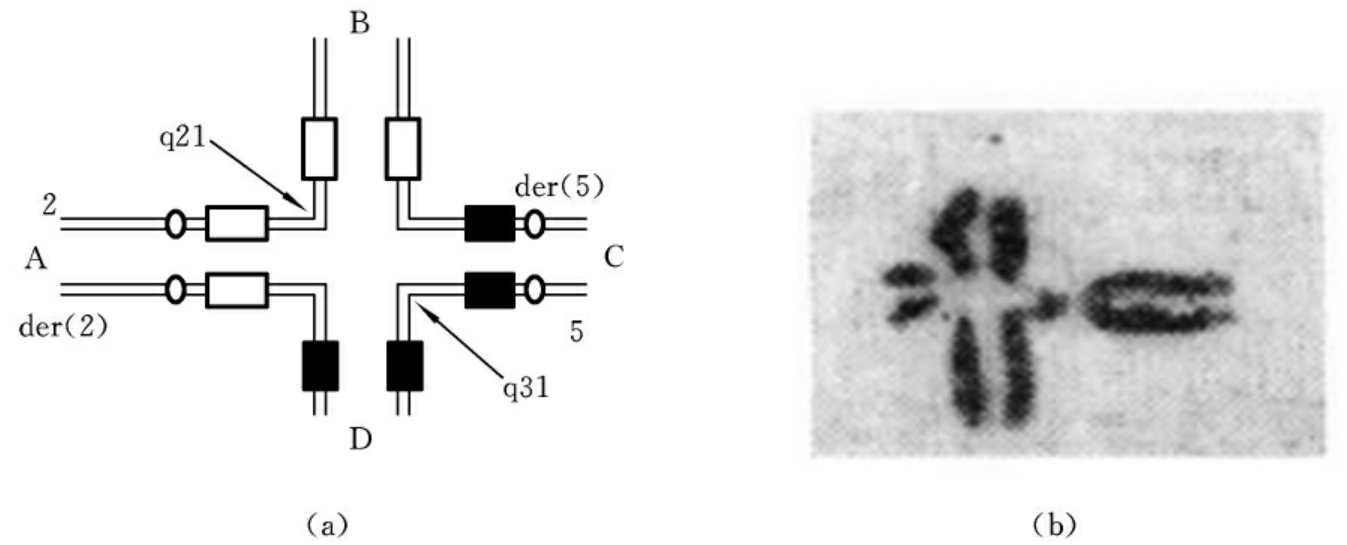
图7-11 四射体
(a)四射体图解 (b)四射体照片
(2)罗伯逊易位(Robertsonian translocation)。又称着丝粒融合(centric fusion),是发生于近端着丝粒染色体的一种易位形式。当两个近端着丝粒染色体在着丝粒部位或着丝粒附近部位发生断裂后,二者的长臂在着丝粒处接合在一起,形成一条由长臂构成的衍生染色体;两个短臂则构成一个小染色体,小染色体往往在第二次分裂时丢失,这可能是由于其缺乏着丝粒或者是由于其完全由异染色质构成所致。由于丢失的小染色体几乎全是异染色质,而由两条长臂构成的染色体上则几乎包含了两条染色体的全部基因,因此,罗伯逊易位携带者虽然只有45条染色体,但表型一般正常,只在形成配子的时候会出现异常配子,异常配子与另一正常配子受精形成的胚胎,可形成死胎而流产或出生的患儿伴有先天畸形。如14号染色体长臂的着丝粒(14q10)和21号染色体的长臂的着丝粒(21q10)同时发生了断裂,两条染色体带有长臂的断片相互连接,即在着丝粒部位融合,形成的衍生染色体包含了21号染色体的21q10→qter节段和14号染色体14q10→qter节段,其余的部分均丢失(图7-12)。
(3)插入易位(insertional translocation)。插入易位是指两条非同源染色体同时发生断裂,其中一条染色体的片段插入到另一条染色体的非末端部位。只有发生了三次断裂时,才可能发生插入易位。
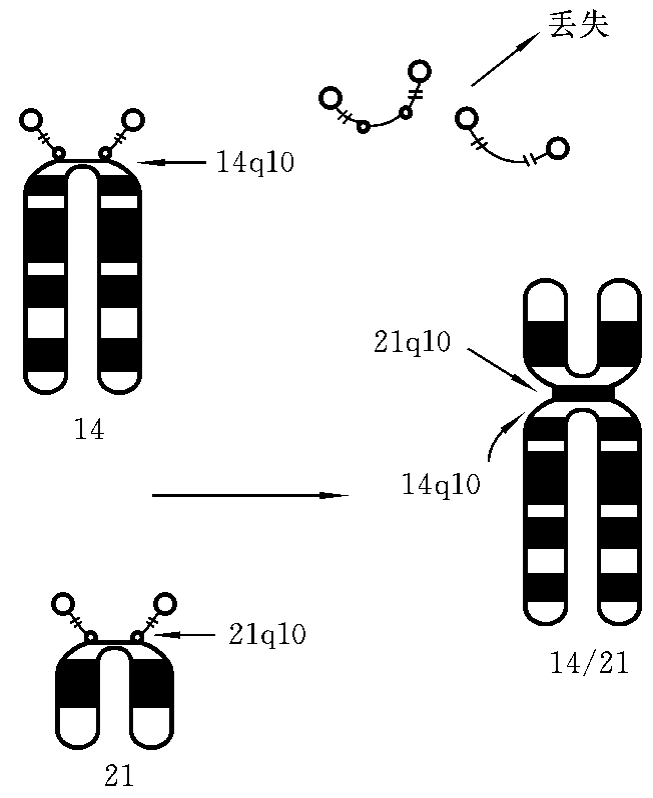
图7-12 罗伯逊易位
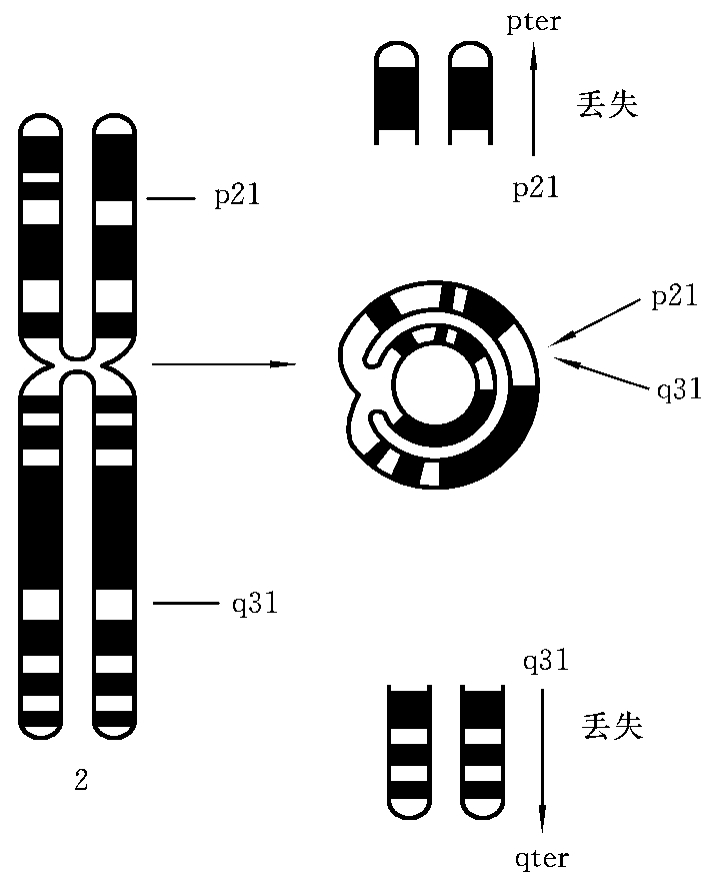
图7-13 环状染色体
5.环状染色体
一条染色体的长、短臂同时发生了断裂,含有着丝粒的片段两断端发生重接,即形成环状染色体。如2号染色体的p21和q31分别发生了断裂,断裂点以远的片段丢失,含有着丝粒的中间片段两断端p21与q31相接形成环状染色体(ring chromosome)(图7-13)。这种结构畸变的简式描述为46,XX(或XY),r(2)(p21q31);详式描述为46,XX(或XY),r(2)(p21→q31)。
6.双着丝粒染色体
两条染色体同时发生一次断裂后,两个具有着丝粒的片段的断端相连接,形成了一条双着丝粒染色体(dicentric chromosome)。在细胞分裂中,如果这条染色体的两个着丝粒分别被纺锤丝向相反的二极拉动,则会形成染色体桥(chromosome bridge)而容易发生断裂,故多数为不稳定的结构改变。如双着丝粒间较为靠近,则可稳定存在和传递。如5号染色体的q31和9号染色体的q21分别发生了断裂,两个具有着丝粒的染色体片段断端相互连接,形成了一条双着丝粒的衍生染色体(图7-14)。这种结构畸变的简式描述为45,XX(或XY),dic(5;9)(q31;q21);详式描述为45,XX(或XY),dic(5;9)(5pter→5q31∷9q21→9pter)。
7.等臂染色体
一条染色体的两个臂在形态、遗传结构上完全相同,称为等臂染色体(isochromosome)。等臂染色体一般是由于着丝粒分裂异常造成的。在正常的细胞分裂中,着丝粒纵裂,姐妹染色单体分离,形成两条具有长、短臂的染色体。如果着丝粒横裂,长臂、短臂各自形成一条染色体,即形成了一条具有两个长臂和一条具有两个短臂的等臂染色体。以X染色体为例:①具有两个长臂的等臂染色体的简式描述为46,X,i(X)(q10);详式描述为46,X,i(X)(qter→q10::q10-→qter)。②具有两个短臂的等臂染色体的简式描述为46,X,i(X)(p10);详式描述为46,X,i(X)(pter→p10::p10→pter)(图7-15)。
8.插入
插入(insertion)是一条染色体的片段插入到另一条染色体中的现象。它实际上也是一种易位。只有在发生了一共三次断裂时,“插入”才有可能发生。插入可以是正向的,也可以是倒转了180°的,即为反方向插入。插入如发生在同源染色体间,就会在一条染色体上发生重复,而另一条同源染色体则缺失了同一节段的染色体。
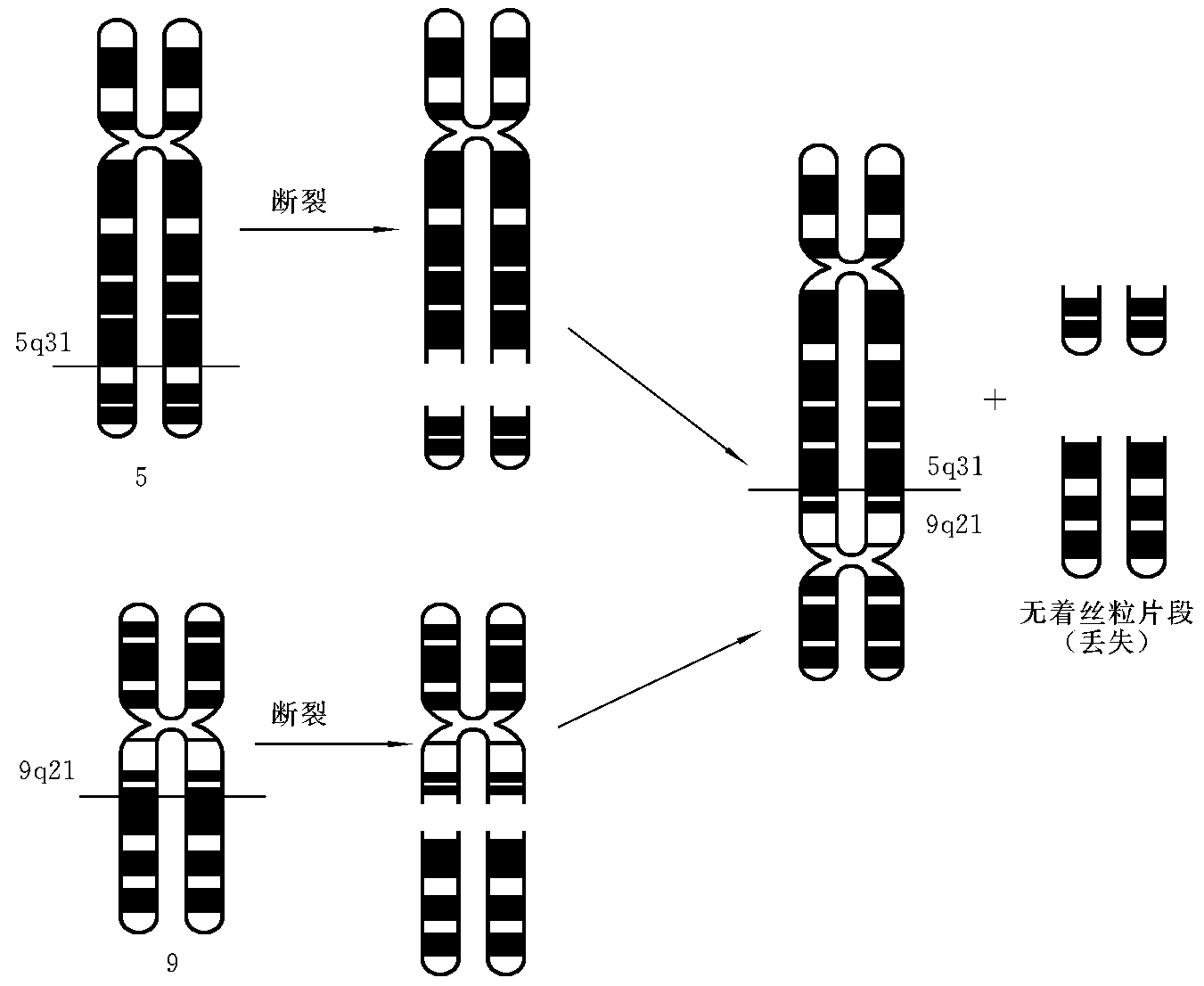
图7-14 双着丝粒染色体
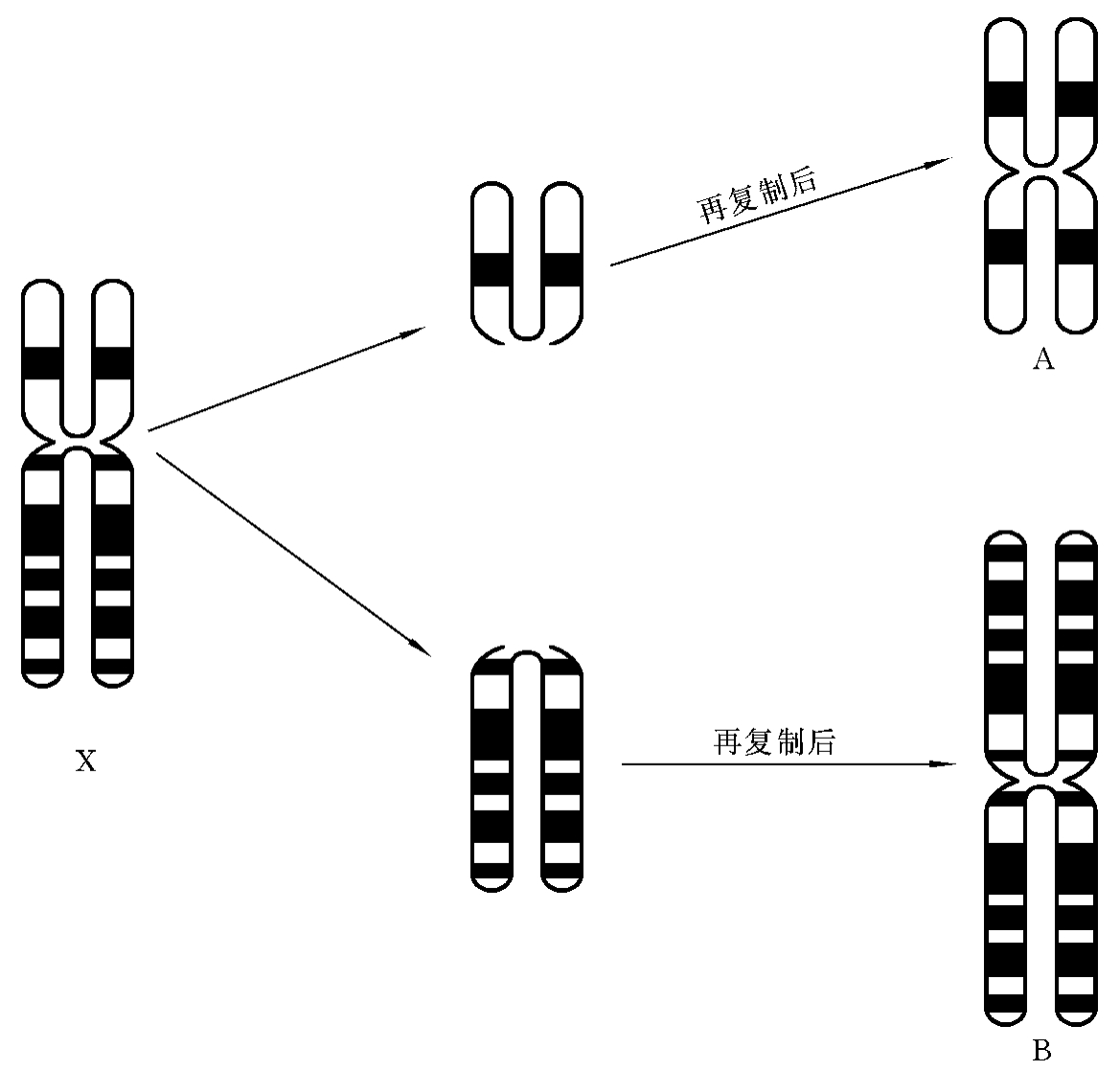
图7-15 等臂染色体
四、嵌合体
嵌合体可以是染色体数目异常之间、结构异常之间以及数目和结构异常之间的嵌合。由三倍体和二倍体两个细胞系组成的嵌合体的发生可能源自紧密结合的一个卵子和一个极体的各自受精,即一个细胞同两个精子受精发育成三倍体细胞系,另一个细胞则与另一个精子受精发育成二倍体细胞系,由于这是由两个起源不同的细胞系紧密结合组成的一个胚胎发育而成的机体,故称为异源嵌合体(chimera)。而由同一个合子发育成具有不同核型的细胞系所组成的个体,称为同源嵌合体(mosaic,简称嵌合体),这类嵌合体的发生是由于卵裂中染色体的不分离。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











