



第二节 细菌遗传变异的物质基础
细菌遗传变异的物质基础是细菌染色体和质粒DNA、转位因子、毒力岛、噬菌体等。
一、细菌染色体
细菌染色体(chromosome)是由两条环状双螺旋DNA长链组成,含细菌的遗传基因,控制细菌的遗传与变异。染色体DNA在细菌细胞浆中呈螺旋形式缠绕成团,附着在横隔中介体上或细胞膜上,不含组蛋白,外无核膜包围,故称核质。细菌染色体DNA以半保留方式进行复制。如大肠埃希菌染色体DNA有5 000多个基因,足以满足控制大肠埃希菌全部遗传性状的需要。
二、质粒
质粒是细菌染色体外的遗传物质,存在于细菌胞质中,为环状闭合的双链DNA。质粒有两类,大质粒含有几百个基因,小质粒仅含20~30个基因。质粒基因可编码产生很多重要的生物学性状。质粒在细菌间的转移是细菌获得某些遗传基因的重要方式。
质粒的基本特征:①质粒具有自我复制的能力,并随细菌的分裂可传入子代细菌。②质粒基因编码的产物赋予细菌某些特殊性状,如致育性、耐药性、致病性等。③质粒并非细菌生命活动不可缺少的遗传物质,可以自行丢失或消除。细菌丢失质粒后照样生存,但由质粒决定的相应性状随之消失。④质粒具有转移性,可通过接合、转化或转导等方式在细菌间转移。⑤质粒可分为相容性和不相容性两种,几种不同质粒共存于一个细菌内称为相容。几种质粒不能共存于一个细菌内称为不相容性。
医学上重要的质粒:①致育质粒(F质粒)编码性菌毛。带有F质粒的细菌(F+菌)可产生性菌毛,称为雄性菌。无F质粒的细菌(F-菌)不产生性菌毛,称为雌性菌。F+菌能通过性菌毛把某些遗传物质(R质粒、F质粒)以接合方式传递给F-性菌,使其获得F+菌的某些遗传性状。②耐药质粒(R质粒)亦称R因子,决定细菌耐药性的产生。带有R质粒的细菌有大肠埃希菌、沙门菌、志贺菌、铜绿假单胞菌等革兰阴性菌。60%~90%革兰阴性菌的耐药性由R质粒转移获得。③细菌素质粒编码各种细菌产生的细菌素。如Col质粒(Col因子)编码大肠埃希菌的大肠菌素。细菌素对同品系或近缘细菌具有抑制作用。④毒力质粒(Vi质粒)编码与细菌致病性有关的毒力因子,如致病性大肠埃希菌肠毒素、破伤风梭菌痉挛毒素、炭疽毒素、金黄色葡萄球菌剥脱毒素均由相应的毒力质粒编码产生。
三、转位因子
转位因子是存在于细菌染色体或质粒DNA分子上的一段特异性核苷酸序列片段,它能在DNA分子中移动,不断改变它们在基因组的位置,能从一个基因组转移到另一基因组中。转位因子通过位移改变了遗传物质的核苷酸序列,或影响插入点附近基因的表达,或转位因子本身携带一定的基因序列。但是否能引起细菌的变异要根据染色体或质粒受转位因子作用后的整体功能状况。转位因子主要有三类:①插入序列(insertion se-quence,IS)是最小的转位因子,长度不超过2kb,不携带任何已知与插入功能无关的基因区域,往往是插入后与插入点附近的序列共同起作用,可能是原细胞正常代谢的调节开关之一。②转座子(transposon,Tn)长度一般超过2kb,除携带与转位有关的基因外,还携带耐药性基因、抗金属基因、毒素基因及其他结构基因等。因此当Tn插入某一基因时,一方面可引起插入基因失活产生基因突变,另一方面可因带入耐药性基因而使细菌获得耐药性。转座子可能与细菌的多重耐药性有关。③转座噬菌体或前噬菌体(proph-age)是一些具有转座功能的溶原性噬菌体,当整合到细菌染色体上,能改变溶原性细菌的某些生物学性状,如白喉棒状杆菌、肉毒梭菌等的外毒素就是由转座噬菌体的有关基因所编码的。
四、毒力岛
毒力岛(pathogenicity island,PAI)是20世纪90年代提出的一个新概念。PAI指病原菌的某个或某些毒力基因群,分子结构与功能有别于细菌染色体,但位于细菌染色体之内,因此称之为“岛”。PAI虽然是染色体的DNA片段,但两端往往具有重复序列与插入元件,其鸟嘌呤(G)和胞嘧啶(C)的百分含量及密码使用与细菌染色体有明显差异,分子量多为30~40kb,也有达100kb者。
五、噬菌体
噬菌体(bacteriophage)是能感染细菌、真菌、放线菌、螺旋体等微生物的病毒。因能裂解细菌故名。噬菌体与细菌的变异密切相关。噬菌体广泛分布于自然界,个体微小,需用电子显微镜观察。在电子显微镜下可见噬菌体的基本形态有蝌蚪形、微球形和纤线形。以蝌蚪形居多。蝌蚪形噬菌体有头部和尾部,并由尾须、尾领连接。头部为双辐射状的六棱柱体,尾部呈管状,尾部中心是尾髓,外包尾鞘,终止于尾板。尾板连接的尾刺和尾丝是噬菌体与敏感微生物接触、吸附的部位(图18-1)。噬菌体的化学成分是核酸和蛋白质。核酸存在于头部,大部分噬菌体的核酸是双链DNA;蛋白质组成头部的外壳和尾部。噬菌体具有严格的宿主特异性,即某一种噬菌体只能感染某一种微生物,甚至只能感染某一种中的某一型。因此,可以利用噬菌体对细菌等进行鉴定与分型。噬菌体对理化因素的抵抗力比一般细菌繁殖体强。一般在70℃30min仍不失去活性,在低温条件下能长期存活。
根据噬菌体与宿主菌的相互关系,可将噬菌体分为两种类型:①毒性噬菌体(virulent phage):能在敏感细菌中增殖并引起细菌裂解的噬菌体称为毒性噬菌体。毒性噬菌体通过尾刺或尾丝特异地吸附在敏感细菌表面相应受体上,尾鞘收缩将头部中核酸经尾
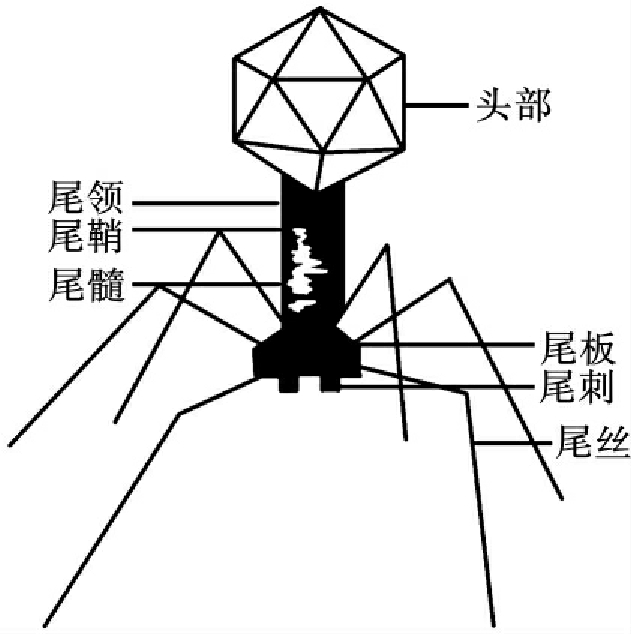
图18-1 蝌蚪型噬菌体结构模式图
髓小孔注入菌细胞内,蛋白质外壳留在菌体外。噬菌体DNA进入菌细胞后,开始生物合成。以复制的方式进行增殖,即以噬菌体DNA为模板,复制子代核酸,合成子代蛋白质,子代DNA与子代外壳蛋白在细菌胞质中装配成完整成熟的子代噬菌体。当子代噬菌体达到一定数目时,菌细胞裂解,释放出噬菌体,此过程称为溶菌周期或复制周期(需15~25min)。②温和噬菌体(temperate phage):感染敏感细菌后不增殖,不引起宿主菌裂解,而是噬菌体的基因整合于细菌染色体中,这样的噬菌体称为温和噬菌体。此过程称为溶原周期。整合在细菌染色体中的噬菌体基因称为前噬菌体(prophage)。带有前噬菌体的细菌称为溶原性细菌(lgsogenic bacterium)。某些前噬菌体可导致细菌基因型和性状发生改变,称为溶原性转换(lgsogenic conrersion)。例如白喉棒状杆菌的白喉外毒素、肉毒芽胞梭菌的肉毒毒素、溶血性链球菌的红疹毒素,它们的产生都与溶原性转换有关。整合的前噬菌体可在某些理化和生物因素的诱导下或偶尔自发地脱离宿主染色体进入溶菌周期,导致细菌裂解(图18-2)。温和噬菌体既有溶原周期又有裂解周期,而毒性噬菌体只有裂解周期。
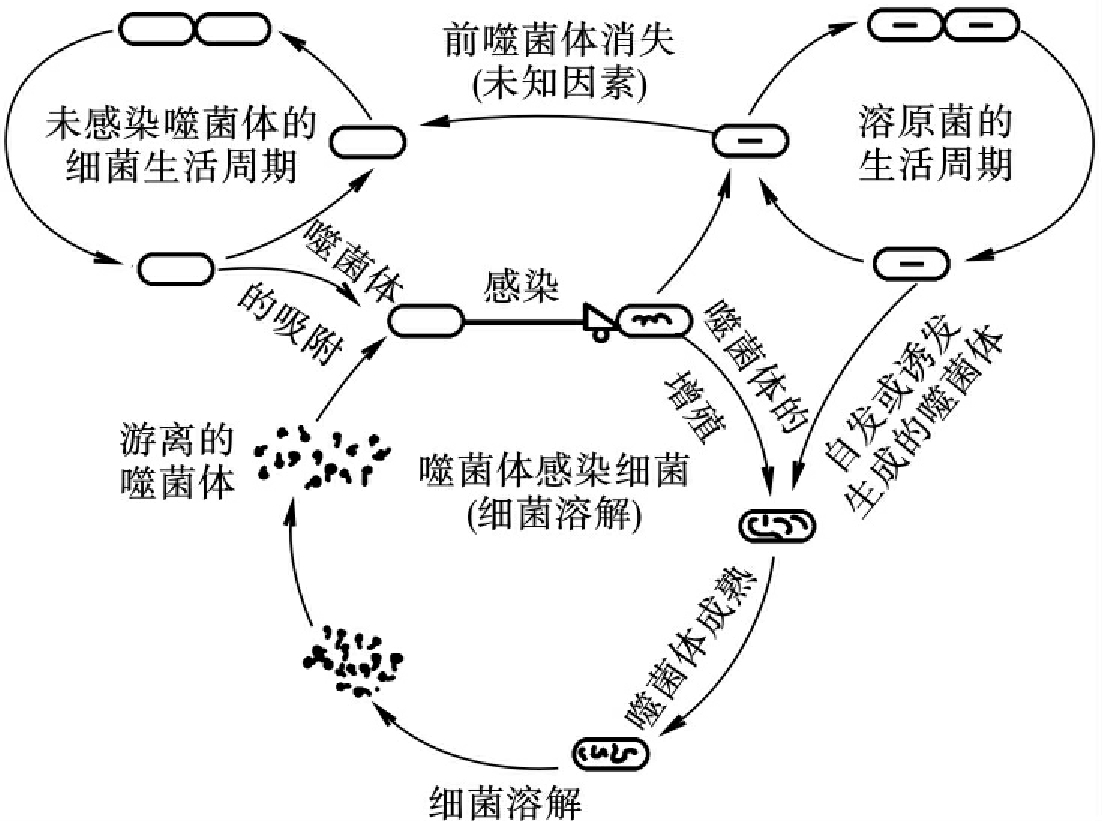
图18-2 噬菌体与细菌相互作用示意图
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










