



第一节 生理因素与学习
长期以来心理学家根据遗传、环境和教育对心理发展的关系的不同认识,发展出了三种不同的观点:遗传决定论、环境决定论和相互作用论。遗传决定论以优生学创始人高尔顿为代表,认为:儿童的心理发展由先天的遗传因素决定,其发展只是内在因素的自然展开,不是后天环境和教育作用的结果。相反,以行为主义者华生为代表的环境决定论者认为:儿童的心理发展主要是后天环境和教育的结果,他们否认了遗传的作用。以皮亚杰为代表的相互作用论是一种折中的观点,这种观点认为:儿童心理的发展是先天的遗传因素和后天的环境与教育相互作用的结果。人类的生长发育是先天遗传和后天环境的因素共同作用的结果。现在很多研究者和教育者都认为:先天遗传是人类生长发育的基础,它决定了后天生长发育的可能性,而后天环境因素又决定了后天生长发育的现实性。
一、遗传因素对学习的影响
遗传是指子代和亲代之间在形态结构及生理功能上的相似,它是人后天的各种能力发展的生理前提和物质基础。一个受精卵包含父母亲所给予的23对染色体,在每一对染色体中又包含了2万个基因。基因是生物遗传物质的最小功能单位,它们是决定和影响个体特征的载体。大多数的基因又是由脱氧核糖核酸(DNA)组成的。对遗传因素影响人类生长发育产生的研究一般有以下三个途径:一是由遗传学提供材料;二是家谱调查;三是双生子的对比研究。其中对双生子的遗传度进行比较研究,考察在相同的生活环境下血缘关系对个体差异的影响是一种非常重要的方法。遗传度是指在遗传和环境因素对个体产生的影响中,遗传因素所占的比例。而双生子又分为同卵双生子(MZ)和异卵双生子(DZ)两种。同卵双生子由同一个受精卵分裂成两个一模一样的子细胞,两者之间的遗传相似度为100%,具有完全相同的遗传基因。异卵双生子分别由两个受精卵发育而成,二者之间的遗传相似度只有50%,它们之间的性状差异来源于遗传和环境两个方面。将同卵双生子和异卵双生子进行比较,就能区分出遗传和环境因素相对作用的大小。国内外的研究发现遗传因素确实影响着以下的这些方面。
(一)遗传因素对智力的影响
智力是人的各种能力的总和。智力受遗传因素的影响的最好证据就是在相同的生活环境、文化教育环境中的儿童的智力水平仍然存在着很大的差异。如莫扎特两岁时就会演奏钢琴,高斯三岁时就能纠正父亲算账时的错误。美国《时代》周刊报道:普通人用右脑来辨别音符,而天才音乐家是使用左脑进行分辨的。普通人通常只能分辨出两个音乐频率,而音乐家一般能分辨出16个。托尔曼曾将白鼠放进迷宫测试,分别选出的最快和最慢的雌、雄鼠作为“聪明鼠”和“笨鼠”作为实验组和对照组进行比较研究。他分别让两组的雌、雄鼠交配。让这两组鼠分别这样繁殖下去。这两组鼠分别繁殖到七八代时,他发现“聪明鼠”的子代更聪明,而“笨鼠”的子代更笨。
美国加利福尼亚大学的詹森在其研究中发现不管是在相同的环境还是在不同的环境中抚养长大的同卵双生子,其智商上的相似程度都是很高的,这说明遗传性越相近,学习能力和智力的潜力越接近,良好的遗传素质是智力发展的重要条件。
研究智力的遗传因素的方法除了双生子智力对比研究外,还有养子与养父母之间、子女与生父母之间智力的相关性比较,以及一直在孤儿院中生活的儿童的智力的差异性比较。采用这些方法的研究发现:子女的智力与其生父母的智力之间呈正相关,在与生父母家庭环境相同的环境中成长的早年分开的养子的智力水平与其生父母的智力水平也呈正相关。
智力低下的儿童也通常被作为研究影响智力的遗传因素的研究对象。有研究发现那些有生理缺陷(有脑损伤,有明确的病因)的智力低下儿童的智力水平是先天因素缺陷造成的结果。而那些没有生理缺陷(无脑损伤,无明确的病因)的智力低下儿童的智力水平又与其家庭的智力水平呈正相关[1]。国内外的许多研究证实了染色体异常是智力低下的重要原因。
另外,生化机制的研究也成为了智力受遗传因素影响的有力证据。Eysenck H J.在1994年对219名在WAIS量表中IQ平均得分为108分的学龄儿童进行脑电波的复杂性和变化性测试后得出,脑电波的复杂性和变化性与IQ的相关分别是0.72和-0.72[2]。还有研究发现,特定的视觉平均诱发电位(AEPs)的潜伏期与IQ呈负相关。影响智力的遗传因素。有研究发现学前组的儿童的IQ遗传度高于学龄组的。有研究表明,IQ在140分以上的儿童长大后其子女的IQ平均值为128分。
众多的研究都证实遗传因素对于智力的形成确实起着非常大的作用。智力作为学习活动中很重要的条件之一,直接影响着学习者的学习效果。正确的认识遗传因素与智力之间的关系,通过科学合理地利用遗传因素来提高智力水平是教育者越来越重视的方向。
(二)遗传因素对语言能力发展的影响
国内研究发现,在相同环境中长大的异卵双生子在以下几方面表现出了很大的差异:语言出现的早晚时间、语声的高低粗细(尖)、说话的多少(好说或不爱说)、掌握的各类语言形式(口头语言、书面语言)、语言机能以及掌握的词汇量的多少等方面,而在相同环境中长大的同卵双生子的差异不大。这充分说明了,遗传因素对于个体语言能力的发展起着举足轻重的作用。
(三)遗传因素对思维的影响
1.对思维类型的影响
巴甫洛夫把思维的高级神经系统分为三种类型:偏抽象型、偏形象型和中间型。其中偏抽象型擅长以抽象思维为主的理科有利于发展数、理能力,而偏形象性擅长以形象思维为主的文科,有利于发展文艺创作能力。这种高级神经类型特点在一定程度上是来自遗传的。在同龄儿童中,兴奋型、易扰型、迟缓型者的脑力工作能力较低,而灵活型者的脑力工作能力较强。
2.对思维的品质和运算能力的影响
林崇德在其1981年的研究中,对在相似或相同环境中长大的同卵双生子和异卵双生子(其中同性异卵和异性异卵双生的各为一半)的几项思维能力指标进行比较后发现:遗传对运算能力和思维的智力品质的影响显著。有研究者通过对相同环境条件下成长的同卵双生子、异卵双生子进行分析后发现,同卵双生子之间的运算能力或学习成绩的相关性最高,同性异卵双生子次之,异性异卵双生子最低。这说明遗传性越相近,学习能力和智力的潜力越接近,良好的遗传素质是智力发展的重要条件。
还有研究者认为遗传对儿童和青少年思维发展的影响是有着年龄特征的。总的趋势是,遗传因素对思维的影响随着年龄的增长而减弱,即随着年龄的增长,遗传因素的作用不如环境与教育的影响那么明显和直接。
虽然每个人之间的遗传因素和生理因素的差异都是明显的,但是这些因素都只是人类各种能力发展的重要条件,而不是决定条件。而且随着年龄的增长,这些遗传和生理因素的作用逐渐表现得不明显,后天的环境和教育因素的作用就相对明显起来。
二、神经系统与学习
(一)神经系统与学习
神经系统分为中枢和周缘两部分。其中中枢部分包括脑和脊髓,周缘部分包括脑神经、脊神经以及周缘的交感、副交感神经结和神经。
1.脑的机能
脑可分为延髓、脑桥、中脑、间脑和大脑两半球五部分(如图13-1所示),它们在结构和机能上是不可分割的整体,但各个部分又有特定的机能。
(1)脑内关于学习的分区。
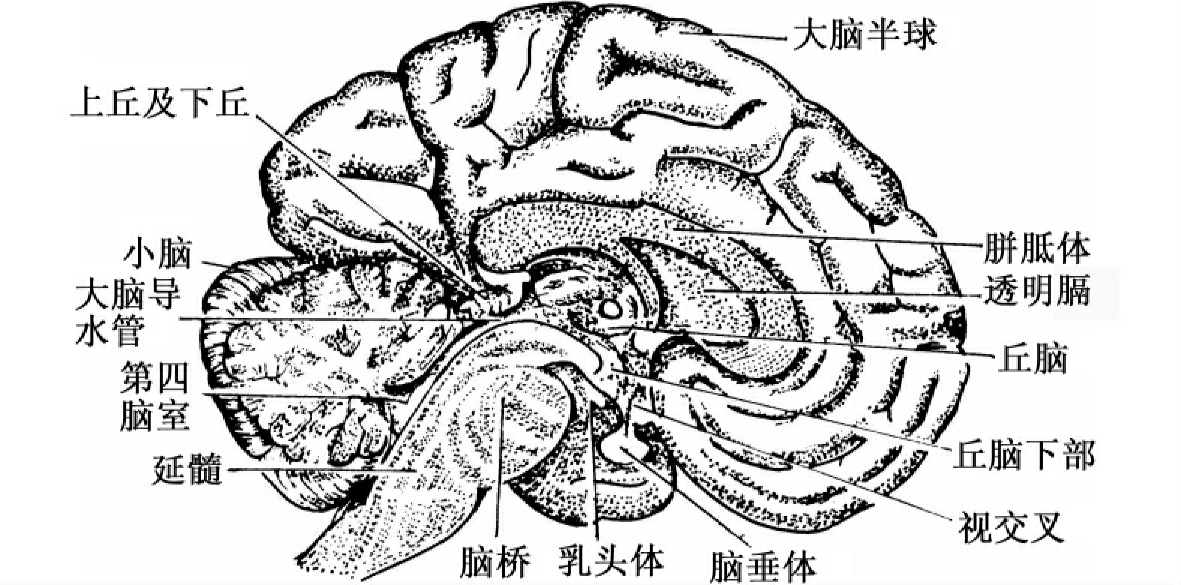
图13-1 脑的正中矢状切面
①大脑皮层与学习。
大脑皮层是人的高级中枢,大脑皮层的发育与学习记忆密切相关(如图13-2所示)。在动物实验中发现,生活在复杂环境中的大鼠的大脑皮层厚,突触联系多;而生活在简单环境中的大鼠的大脑皮层薄,突触联系少。这说明,学习记忆活动多的大鼠的大脑皮层更发达。
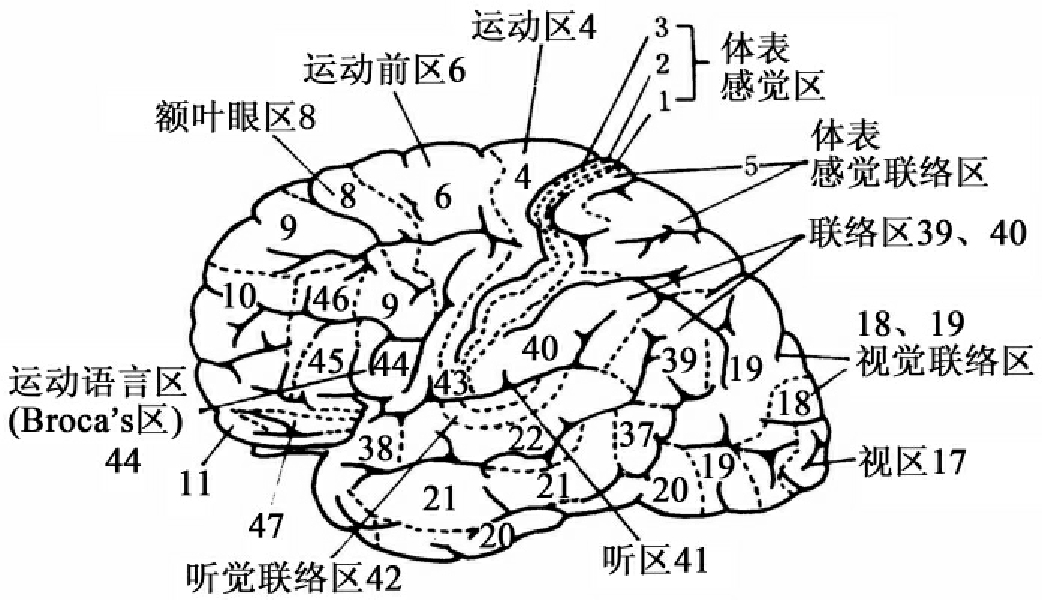
图13-2 大脑皮质细胞结构分区图
具体来说,前额叶皮层与运动学习行为、复杂的时间、空间定向的学习有关。前额皮质的活动与工作记忆有关。其中前额侧面参与简单的短时记忆操作,前额的后面参与执行高水平的操作。右侧前额参与持续性注意左前额皮质比右前额皮质更多地参与语义记忆的提取,而右前额皮质则更多地参与情景记忆的提取。有研究表明,在对记忆材料进行编码、使之进入情景记忆时,如果所处理的是语言材料,则前额的激活主要是左侧的,而对非言语材料进行编码时,会出现左侧或右侧的激活[3]。在语义操作中,左前额的损伤会十分明显地导致词汇流畅性方面的障碍。
颞、顶、枕联合区皮层与感觉和空间关系的学习有关。左侧颞叶与编码相关进行工作记忆时,顶叶也会出现激活[4]。顶叶参与持续性注意,顶叶下部控制着选择性注意。枕叶在词汇的视觉处理方面起着主要的作用,在视觉词汇识别的过程中,枕叶的侧面和视觉皮层都会受到激活。
视觉代表区:在枕叶距状裂两侧(Brodmann第17区)。刺激该区,可以使患者产生简单的主观光感觉,但不能引起完善的视觉形象。右侧枕叶主要与两眼的左侧视野有关,而左侧枕叶主要与两眼的右侧视野有关。一侧枕叶皮质受损造成对侧偏盲,两侧皮质受损则造成全盲。
听觉代表区:在颞叶的颞横回(Brodmann第41区)。电刺激该区可以使患者产生铃声样或风声样的主观感觉。一侧内叶皮质受损并不会影响听觉,只有左右两侧听觉代表区同时受损害才产生完全的耳聋。
②边缘系统与学习。
边缘系统包括扣带回、海马、杏仁核、膈、前丘脑和乳头体。边缘系统实现伴有情绪体验或情绪反应的学习,其中海马是与记忆活动关系最为密切的脑区。海马在辨别空间信息、新异刺激性调节和短时记忆向长时记忆的过渡中起着重要的作用。临床上由于治疗的需要而切除双侧颞中叶的病人,如损伤了海马及有关结构,则引起近期记忆能力的丧失,丧失的程度常取决于损伤部位的大小。损伤的病人只能回忆和提取信息,而不能形成新的长时记忆。海马的不同区域对学习和注意的学习与记忆的作用是不同的,对海马腹侧的破坏比背侧的破坏更能引起严重的学习和记忆障碍。
③小脑与学习。
小脑在运动技能的学习中起着重要的作用。小脑与快速运动反应性学习密切相关,是简单运动条件反射形成中最基本和最必要的脑结构。一些复杂的运动技巧如走钢丝等的学习也都受小脑的控制。
(2)大脑两半球机能的分工和协作。
大脑由对称的左、右两个半球组成。分割左、右两半球的深沟称为纵裂。纵裂底部由胼胝体相连。1973年Luria提出了“功能系统”的概念。他认为功能系统是脑的许多皮层和皮下区域神经纤维通路联系而协调地工作。脑的功能是把各种不同的脑联系联合起来,形成不同技能系统的协调活动。左半球在语言与和数字等有关的记忆任务中的活动表现得明显,而在使用非言语材料时,激活则是双侧的[5]。
2.神经系统的信息加工网络
神经系统由神经细胞(神经元)和神经胶质组成。神经元(如图13-3所示)是神经系统结构和功能的基本单位。一般的神经元由胞体、树突、轴突、髓鞘和突触五部分组成。神经元之间通过突触、反射弧以及神经网络进行联系。
(二)生理发育与智力和能力发展的关系
1.脑重的增加
婴儿在出生时的脑重为:350~400g,大约是成人脑重的25%。出生后6个月,脑重达700~800g,约为成人脑重的50%。12个月时达到800~900g,24个月时达1050~1150g,36个月时的脑重已接近成人水平。之后发育速度慢慢变缓,在15岁时达到成人水平。
2.大脑皮质的发育
胎儿在六七个月时,脑的基本结构就已具备。出生时脑细胞已分化,细胞构筑区和层次分化已基本完成,大多数沟回都已出现,脑岛已被邻近脑叶掩盖,脑内基本感觉运动通路已髓鞘化(白质除外)。之后,婴儿皮质细胞迅速发展,层次扩展,神经元密度下降且相互分化,突触装置日益复杂化。到了2岁时,脑及其各部分的相对大小和比例,已基本类似成人大脑。白质已基本髓鞘化,与灰质明显分开。其中,大脑的髓鞘化程度是婴儿脑细胞成熟状态的一个重要指标。整个皮质广度的变化与髓鞘化程度密切相关。
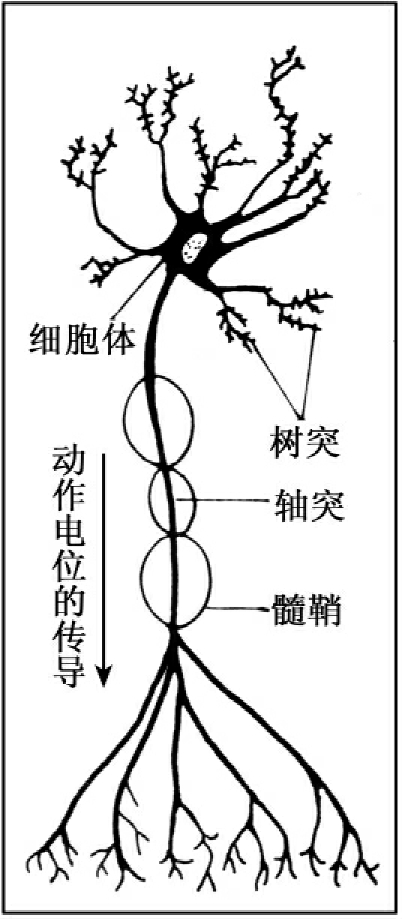
图13-3 神经元
3.脑电波的发展
在大脑皮质表面记录到的“自发”节律性电位活动成为皮质电图(ELOG),在头皮表面记录到的称为脑电图(EEG)。根据频率范围把脑电图划分为α、β、θ、δ四种基本波形。其中:
α波是每秒8~13次范围内的电位活动,振幅范围为50~100微伏。α节律在大脑的各个区都有,但是在顶枕部最为明显。睁眼时发生α阻断现象(消失),闭眼后又复出现。在轻睡时α节律逐渐消失,唤醒时又恢复原来清醒时的α节律。情绪紧张时α节律也可能消失,或波幅降低,但是过度换气后α节律又可恢复。α节律可以作为意识水平的指标,表示安静、不瞌睡、不做定向思考时的精神状态。
β波是每秒13~30次范围内的电位活动、振幅为20~50微伏。β波以额叶区中央最明显。当被试睁眼视物,或听到突然响声,或思考时α波阻断,出现β波。情绪激动或焦虑状态可使β节律增多。β节律一般代表大脑皮质的兴奋性。
θ波频率为每秒4~7次,波幅为20~40微伏,是儿童的正常脑电活动,成人在困倦时一般也可见到。轻睡时α波逐渐消失,θ波首先出现于前额区。θ节律的出现可视为中枢神经系统抑制的表现。
δ波的频率为每秒0.5~3次电位活动,波幅为10~20微伏,为儿童的(正常)主要脑电活动,任何年龄的人睡眠时都有。成人清醒时出现δ波,表明皮质和皮质下可能病变或智力障碍。
在头皮上记录到的与刺激核反应出现有一定关系的电位变化,成为事件相关电位(event-related potential,简称“ERP”)。其中P300被认为与预期或注意一类心理现象相关。当意外的某种声音出现引起人的注意时就能记录到P300的波[6]。
三、学习时伴随的生理变化
神经科学家近十余年的研究发现,只要给予大脑适当的刺激(学习和工作用脑),则可显著地增加神经细胞的体积和某些细胞的数量。换言之,只要给予大脑适当的刺激,脑细胞便不会随着年龄的增长而衰减,甚至大脑的许多部位还可以继续增长,不断提高大脑功能和智力。
(一)学习时伴随的脑电生理变化
近年来,除了一直采用的脑损伤法外,研究者们开始借助于诸如正电子断层扫描技术(PET)、神经磁成像(MEG)和核磁共振(MRJ)等的无创技术来研究学习时的脑电变化。
1.平均诱发电位(AEP)的研究
Hendrickson(1982年)对219名青年被试(121名男性、98名女性)进行研究:对被试进行WAIS量表(韦克斯勒成人智力量表)测试并计分和利用EEG(脑电图)测量各被试的AEP(听觉诱发电位)数值。两种生理指标的测分与WAIS中11个分量表之间的相关程度值得人们注意。为了验证这一结论,Stough、Nettel-beck和Coppper(1990年)用修订的WAIS也发现了在被试的AEP测分中和他们的言语智商(VIQ)、操作智商(PIQ)、全量表智商(FSIQ)之间存在较高相关。
2.神经传导速度(NCV)的研究
Reed和Jensen(1992)召集了147名男大学生分别进行了如下测量:测量被试在APM量表(高级推理测验)上的得分;测量他们的视觉诱发电位(VEP);用各被试脑的长度和VEP出现之前的潜伏期之间的比值求出各被试的视神经传导速度。经统计表明,被试的NCV值(视神经传导速度)和APM分数之间呈正相关(37),在147个NCV数值中,从最低的1.75米/秒到最高的2.22米/秒不等。这说明随着IQ的提高,视神经传导速度也相应地加快。
(二)学习时的生物化学变化
1.人脑内葡萄糖代谢速度的研究
Haier等人(1988)提出一个假设,认为人脑内葡萄糖代谢速度和IQ得分之间存在显著负相关。他进一步指出,个体在APM量表上得到高分者在对量表各条目做出反应时花费了较少的脑能量。为了验证这一假设,他(1992)选取了8个男被试分两次测量了他们脑内的葡萄糖代谢速度,两次测量分别安排在:被试开始学习一个复杂的电脑游戏软件的初始阶段;经过4~8周每天练习后的后期阶段。结果表明,练习后被试的操作获得明显改进,他们脑内葡萄糖的代谢速度也明显降低了,而且学得最好的被试其代谢速度也下降得最快。统计结果证实了最初的假设。这一研究结果与其他人的研究相一致,例如Parks等人(1988)选取16名被试,通过用PET测量他们脑内葡萄糖的代谢速度及另一项反映言语流畅性的神经心理测试,也发现了两者之间的负相关。
2.脑内核糖核酸的含量和成分的变化
核糖核酸(RNA)曾被认为是记忆分子。它在神经细胞中大量存在,在其结构中,拥有一个潜在的大仓库可容纳已编码的信息,而且它还决定并控制着在细胞内合成的蛋白质的特殊形式。当神经细胞受到反复的刺激时,RNA在神经细胞中的浓度会增加。对参与学习某种技能的主要细胞施以刺激时,这一细胞中的RNA不仅有所增加,而且在学习后的分析中还可发现RNA的特性有了改变,表现为四种碱基量比例(“基础比”)有变化。海登(1964)训练一组大鼠偏废惯用于取食的爪,而强迫用另一只爪取食。训练完之后,测定控制它们前爪的感觉运动皮层内第五层锥体细胞的RNA,以另一半球对称部位的锥体细胞为对照比较材料。结果表明,学习后相应一侧皮层内RNA含量较对照皮层有显著增加,RNA的内部结构也有变化。
3.脑内蛋白质含量的变化
海登(1968)在进行上述的换肢取食训练之后发现,这种学习过程伴随着脑内酸性蛋白质(S100)的含量的增加,这种蛋白质仅仅发现在脑内存在。然后他用抗S100蛋白血清注入其他大鼠,结果,这些大鼠的学习成绩显著落后于未接受处理的大鼠。因而认为,学习训练后脑内酸性蛋白质S100的增加,的确与学习有关。其他报道也证实了海登所发现的事实。
4.脑内pH值的变化
Carluin和他的同事(1995)对42个年龄为6~13岁的男孩大脑中的酸度测定后发现,人的智力可能与pH值有关。IQ较高很可能与大脑的碱性较高或较多有关。他们用WAIS对上述被试进行了智商测试,发现pH值超过7.0的测试对象,其智商水平比一般人高出近一倍。
5.其他生物化学变化
一些研究表明,那些脑内乙酰胆碱(ACh)、乙酰胆碱酯酶(AChE)含量比较高的老鼠的学习能力更强,并且合成的乙酰胆碱类似物(AF102B)可以显著地减轻大鼠记忆力下降的现象。美国华裔科学家钱卓博士等认为NR2B是有利于学习和记忆的基因,它能够控制和产生在记忆中其作用于名为NMDA的蛋白质。他们给实验鼠的受精卵多添加一份NR2B,然后将其放入试验母鼠的子宫中孕育。产下的实验鼠与普通鼠相比,学习能力更强,记忆的持久性也更长。海登对托尔曼实验中的老鼠的脑细胞进行化学分析后发现,“聪明鼠”脑中的DNA比“笨鼠”的高10%。并且如果在“笨鼠”的体内注入这种神经多肽物质,“笨鼠”也变得聪明起来了。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










