



【学习任务】
力量、速度训练中肌肉产生较大的力,用于克服大阻力或者在较大阻力下快速收缩,运动时间较短,能量供应以无氧代谢供能为主,训练方法中应充分调动无氧供能系统产生能量,并在运动后刺激肌肉壮大,产生的适应特点主要是无氧供能系统供能能力提高,肌肉蛋白质合成增多。可以通过血乳酸测试反映运动中无氧供能能力。耐力训练中肌肉产生的力相对较小,但是运动时间较长,能量供应以有氧代谢供能为主,无氧阈运动强度下有氧代谢供能能力达到最高,产生的适应特点主要是心血管系统供应血液和氧的能力提高,肌肉线粒体数量和有氧代谢系统供能能力提高。运动疲劳的因素十分复杂,其生化变化机理还在不断深入研究之中,神经—肌肉疲劳链和神经—内分泌、免疫系统和代谢网络疲劳理论从整体上总结了目前关于运动疲劳环节的生化机理。机体对运动训练的恢复和适应是保持和调节人体全身机能稳态的系统性问题,其中人体物质和能量代谢能力的提高是基础之一,表现为蛋白质合成增加、能源物质超代偿恢复、能量生成的酶活性的提高、线粒体数量增加等。上述运动代谢是运动营养的生化原理和实践应用的基础。运动疲劳的规律和适应的规律是制订训练计划的重要基础。
【学习目标】
1.了解力量和速度训练中能量供应的特点。
2.掌握和应用无氧低乳酸训练中能量供应系统能力生化原理。
3.掌握和应用高乳酸重复训练中能量供应系统能力生化原理。
4.理解力量和速度训练适应的生化机理。
5.了解并应用力量和速度训练中营养的消耗特点和运动后营养补充的生化原理。
6.掌握耐力训练中能量供应系统的供能特点。
7.理解耐力训练适应的生化机理。
8.掌握并应用乳酸阈训练的生化原理。
9.掌握多种不同特点的耐力训练中能量的供应特点。
10.综合分析实际的运动训练实践中,同一运动项目多种训练方法中不同的能量代谢特点。
在运动中,骨骼肌通过分解能量物质产生能量,并用于骨骼肌的收缩,进而产生作用于骨骼的力,身体不同部位的骨骼肌作用于骨骼的力有多种形式和特点,使全身各环节在运动中保持协调和相对稳定。我们一般习惯性地按照肌肉用力的大小,将运动训练强度分为最高强度训练、高强度耐力训练和耐力训练。
从不同的角度来看,人体运动中力量的特点和性质多种多样,有多种描述表达方式,如爆发力、快速力量、速度力量、耐力力量、速度耐力力量等,可以参考其他学科如运动生理学、运动训练学,以及运动专项训练的专著。按照运动中肌肉内能量代谢的特点,可以直观简要地把肌力训练分为力量和速度训练及耐力训练。但是,力量和速度训练及耐力训练的含义并不仅限于肌力的训练,是全身各器官在中枢神经系统的支配和调节下,产生综合协调的效果。
力量和速度训练包括发挥最大肌力以对抗阻力,或者肌肉以尽可能快的速度收缩产生快速的环节运动,运动的特点是运动时间较短,肌肉产生的绝对力量较大,包括常见的克服大阻力的运动如举重等典型力量项目,以及克服较大阻力的快速运动如短跑等典型的速度项目,运动中肌肉的能量供应以无氧代谢供能系统为主。
耐力训练包括运动中肌肉产生的力量相对较小的运动,运动的特点是运动时间相对较长、运动环节的速度相对较慢,包括长跑、马拉松等典型的耐力项目,运动中肌肉的能量供应以有氧代谢供能系统为主。
当然,还有很多的运动项目中,运动中既有典型的力量和速度性的运动,也有典型的耐力性的运动,如很多非周期性的集体项目;也有一些周期性的运动项目如中距离跑(800,1 500 m)等,运动中肌肉兼有大力量、快速度和长时间耐力力量的特性,运动中能量供应来源中无氧供能系统和有氧供能系统都占有较大比重。同时,任何项目的运动员都会根据专项能力素质发展的需要,对不同的肌群进行力量训练、耐力训练以及中间类型的多样化训练。
由于实际运动实践中肌力特点的复杂性,为了便于学习运动训练中物质与能量代谢和产生的适应特点,我们将其简化为力量和速度训练、耐力训练,中间各种类型运动中物质与能量代谢和适应特点一般介于两者之间,我们可以在具体项目中再进一步深入了解。我们先简要回顾骨骼肌力量生成的过程,并在此基础上学习力量和速度、耐力训练中骨骼肌内的能量和物质代谢的特点与适应。
在第三章的学习中,了解了运动中的能量供应系统,在此基础上,这一节学习力量和速度、耐力训练中的物质代谢能量供应,可以简要地概括为运动时物质与能量代谢的两个过程和4个系统。具体情况主要取决于运动强度及运动强度的变化,如图4.1所示。
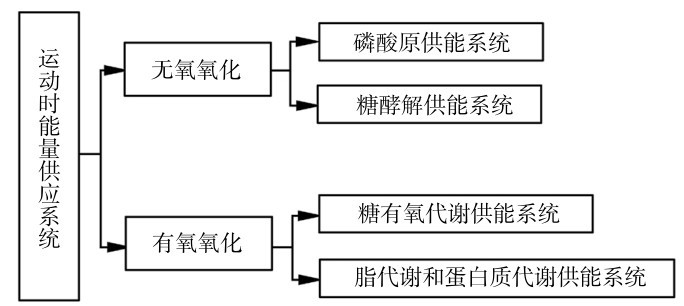
图4.1 运动时物质与能量代谢的两个过程和4个系统
一、运动中能量是连续释放的过程并与运动强度紧密相关
如图4.2所示,包括3个随运动强度而变化的转折区,分别称为糖阈、无氧阈或乳酸阈、磷酸肌酸阈。

图4.2 运动时腿部肌肉不同能源物质生成ATP速率与运动强度关系估算
1.糖阈
处于安静或轻度活动时,人体主要消耗脂肪供能,随运动时能量需求大于脂肪供能输出功率,也即运动负荷超过30%~50%VO2max时,糖氧化功能明显增加,这一转折点称为糖阈。当运动强度在糖阈之下,如慢跑、日常生活活动和其他低强度体力活动中,脂肪和糖都参与供能。
2.无氧阈或乳酸阈
当运动负荷达到55%~75%VO2max时,运动时能量需求大于脂肪供能和糖氧化供能产生的输出功率,则通过糖酵解途径供能迅速增加,这一转折点称为无氧阈或乳酸阈。如400~800 m跑等持续2~3 min的高强度运动。
3.磷酸肌酸阈
当进行85%~95%VO2max的最高强度运动时,一方面运动时能量需求大于糖酵解供能产生的输出功率;另一方面由于乳酸的大量生成使肌肉细胞内[H+]持续增加而抑制糖酵解,此时肌肉中则启动磷酸原供能系统,以维持和适应最高强度运动对能量和功率的需求。这一转折区称为磷酸肌酸阈。
根据运动时运动负荷和运动时间的特征,进一步把机体运动时的物质和能量代谢类型进行细分,并与运动项目相对应,形成不同运动项目训练中制订训练计划的基础依据,如图4.3所示。[想一想]
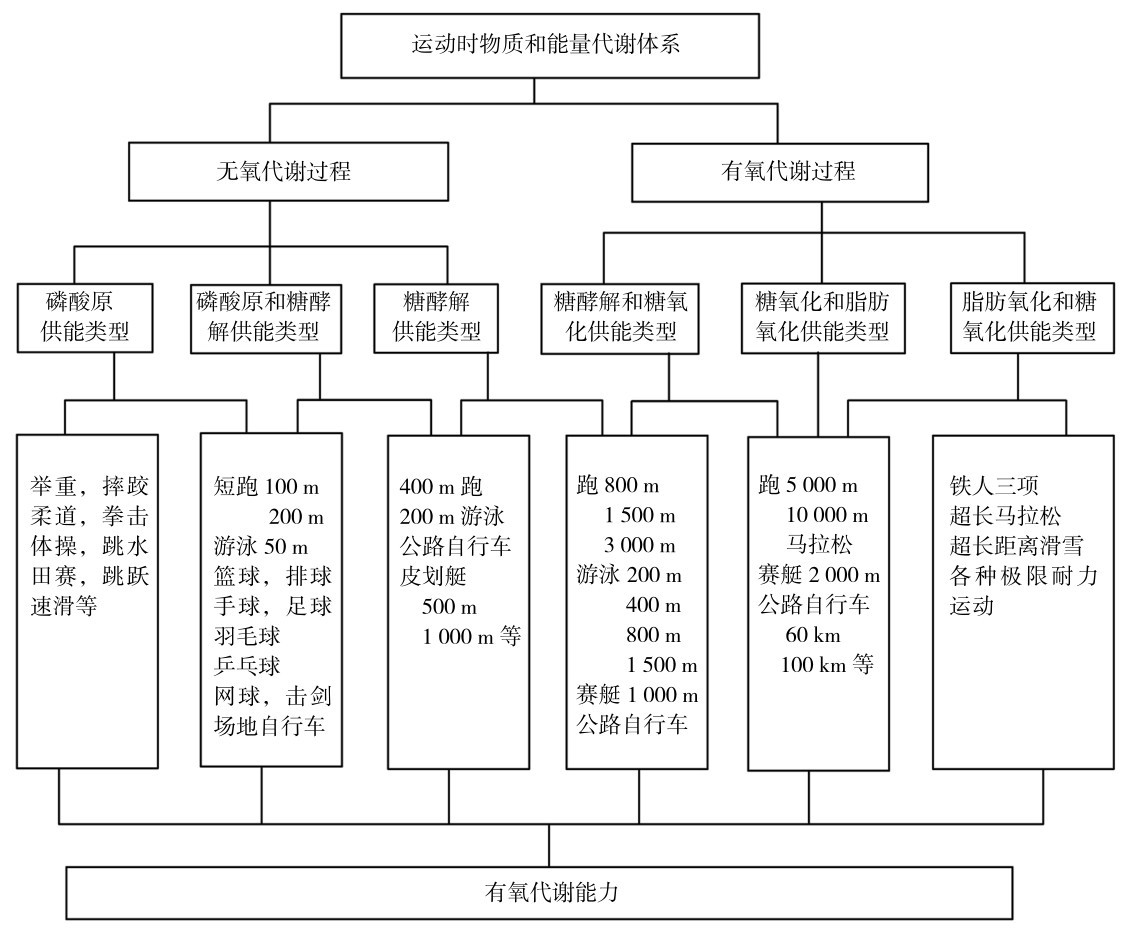
图4.3 运动时物质和能量代谢过程与各运动项目的分布
伟大的牙买加运动员博尔特,在100,200,400 m跑中,均有惊世骇俗的表现;我国著名女运动员邢慧娜在1 500,5 000,10 000 m跑中均有世界级的成绩。但是他们在不同项目中的运动速度节奏有很大差别,自然他们运动中的能量供应特点也有明显区别。请查阅资料,对他们的运动表现细节及与其他不同水平的运动员进行比较,你能发现什么吗?
二、利用运动时物质和能量代谢的规律,提高训练的针对性和训练效果
(一)在发展既定供能系统能力的训练中,严格控制运动强度和运动时间
参考表3.1和表4.1,不同时间最大强度运动中供能代谢的分布比例是不一样的。
表4.1 不同时间最大强度运动时供能代谢的分布

(二)发展运动中能量供应系统能力素质的训练方法
1.提高磷酸原供能系统供能能力素质的训练(对应于最大力量和最高速度能力)
提高磷酸原供能系统供能能力素质的训练,又称为无氧—低乳酸训练,其训练原则为:
①最大速度或最大力量的训练,练习时间以不超过10 s为宜。
②两次动作训练的间歇时间不短于30 s,60~90 s的间歇具有较好的训练效果。
③两组训练练习的间歇不短于3 min,3~4 min的组间休息间歇是磷酸原物质恢复的最短时间。
两次动作训练的间歇时间不短于30 s,是发展磷酸原供能系统训练的关键因素,如图4.4所示显示,休息时间过短,则糖酵解供能增加(血乳酸增加)。
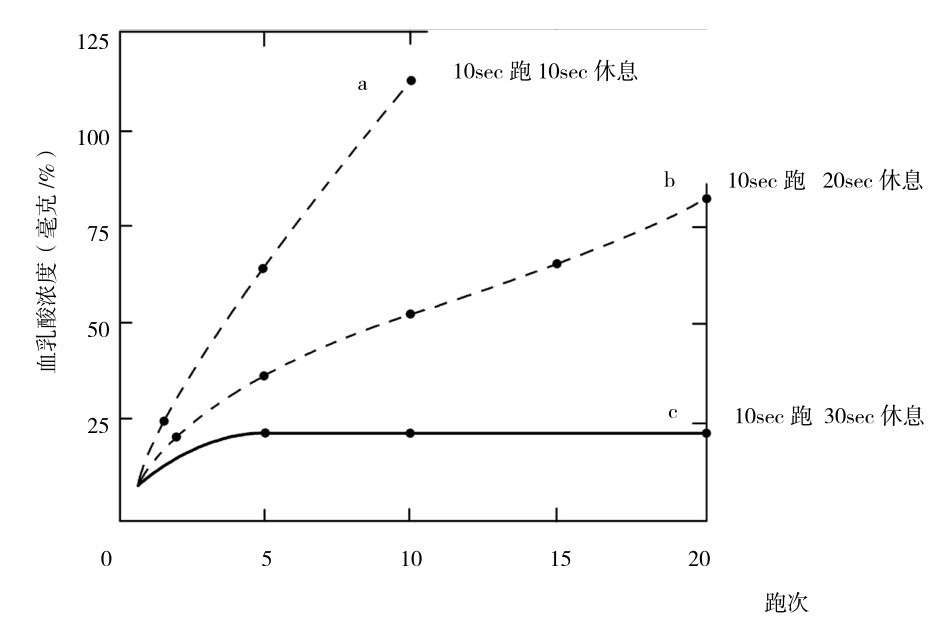
图4.4 20次10sec全力跑间歇休息不同时的血乳酸浓度
2.提高糖酵解供能系统供能能力素质的训练
①最大乳酸训练法。采用运动时间为1~2 min,两次运动之间间歇1~4 min(强度越大间歇时间越长)、多次数(10~20次以上)或每组4~6次运动、组间间歇2~5 min(强度越大间歇时间越长)的多组数间歇练习,使机体乳酸达到最大堆积,提高机体乳酸耐受能力,从而提高机体最大糖酵解供能能力的训练方法,称为最大乳酸训练法。如图4.5所示为最大乳酸训练法中血乳酸浓度的持续升高。
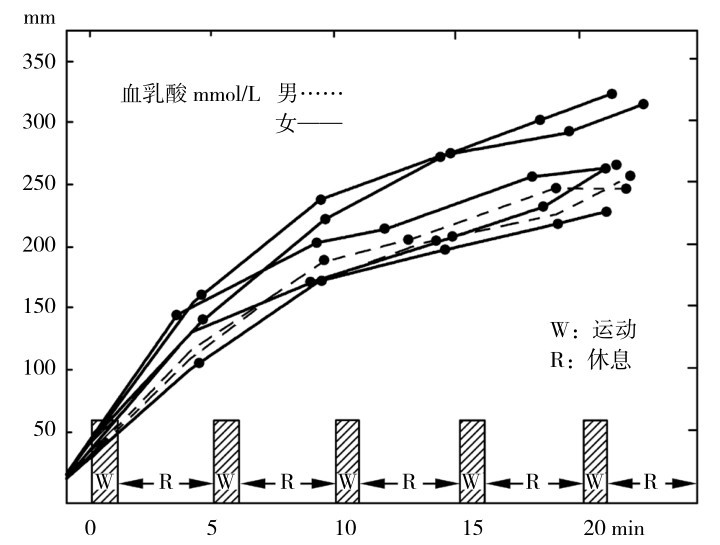
图4.5 5次1 min极量运动,间歇休息4 min后的血乳酸变化
需要注意的是,在间歇期和训练后要采用低强度训练积极恢复的方法加速血乳酸的消除。对于提高中距离项目和长距离项目中的速度能力,最大乳酸训练法是有效的训练方法。
②乳酸耐受力训练法。采用1~1.5 min超负荷的最大运动强度和4~5 min休息间歇的多次重复性训练,使每一次练习中血乳酸均达到最高值,通过休息间歇消除产生的乳酸,使之不会产生积累的训练方法,称为乳酸耐受力训练法。适用于中长距离项目提高速度耐力,训练中血乳酸保持在6~12 mmol/L,又称为强化间歇训练,如图4.6所示。其中图4.6下部为较低血乳酸浓度的乳酸耐受力训练,又称为乳酸阈上间歇训练,对应于一般速度耐力,相应地练习的重复次数较多,应根据实际项目的特点选择使用,如马拉松项目,或者较低训练水平的运动员使用。
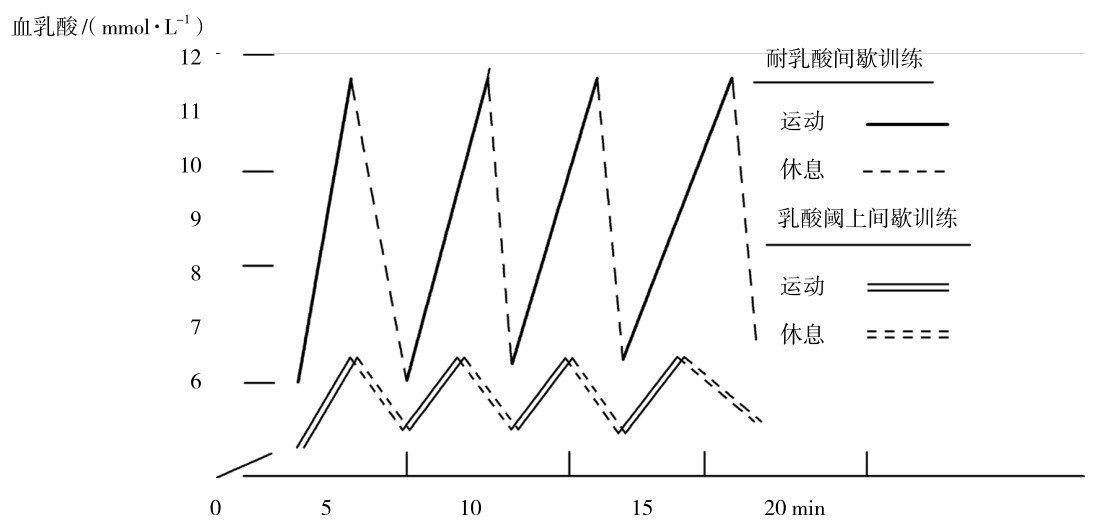
图4.6 中长距离运动项目耐乳酸和乳酸阈上间歇训练的乳酸浓度
延伸阅读
场地自行车运动员提高糖酵解供能能力的训练方法
在提高糖酵解供能系统供能能力素质的训练中,血乳酸是衡量无氧糖酵解能力的直接指标。测运动后的乳酸峰值时,选择合适的取血时间非常关键。负荷时间越短,乳酸峰值越延迟出现,否则,反之。如1~2 min的高强度运动,取血时间一般安排在运动后第3, 5,7 min。若运动时间在4 min以上,在运动后第1,3,5 min取血。表4.2是对无氧耐力训练中最高乳酸训练和乳酸耐受能力训练的监控。
表4.2 我国自行车成年运动员无氧训练强度特点与训练监控
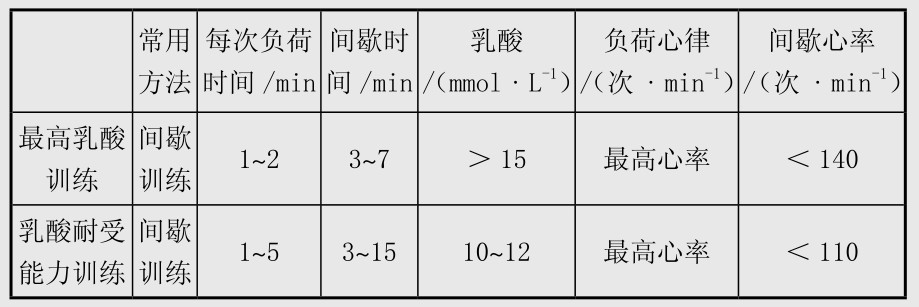
最高血乳酸训练要求每组以最大强度运动1~2 min,间歇时间是负荷时间的2~4倍,优秀短距离运动员动员较快,第二组负荷后就可达到本人的最高血乳酸浓度,一般水平的运动员在第四组负荷后可以达到乳酸峰值。
在以乳酸耐受能力为目的的间歇训练中,为防止乳酸值过分升高,疲劳过早出现,后面负荷的间歇时间往往较前面负荷的间歇时间长。另外,上述规律可能不适用于青少年运动员。
3.提高有氧供能系统供能能力素质的训练
乳酸阈(无氧阈)强度训练法如下:
在运动中,血液中的乳酸浓度是乳酸不断由肌肉进入血液和离开血液不断被其他组织吸收这一过程平衡的结果。在递增负荷运动中,随着运动强度的加大,当血乳酸浓度达到4 mmol/L时,所对应的运动强度(以对应的最大摄氧量百分比、运动速度、输出功率等来表示运动强度)称为乳酸阈强度。此时机体由以有氧代谢供能为主转向无氧代谢供能迅速增加的临界转折点。在训练中通过测试血乳酸浓度,找到与4 mmol/L血乳酸浓度对应的运动强度,并以此强度进行的训练称为乳酸阈(无氧阈)强度训练法,可以有效地提高机体的有氧代谢能力。如图4.7所示运动中血乳酸随运动强度和对应的乳酸阈运动强度变化。
随着运动员机体对训练强度的适应和运动能力的提高,运动员个体乳酸阈强度会发生变化,需要定期(4~6个月)测试确定。大量研究证明在乳酸4 mmol/L时运动员的跑速与耐力高度相关,故乳酸阈强度训练就成为提高耐力素质的重要训练方法。
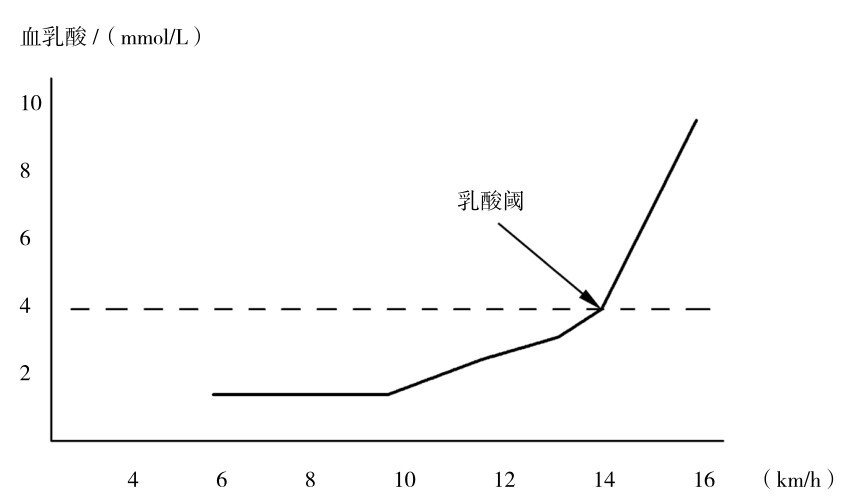
图4.7 递增负荷跑台跑乳酸阈曲线
知识拓展
个体乳酸阈
4 mmol/L的乳酸阈并未考虑运动时乳酸动力学的个体特点,忽略了机体的个体差异性,应根据运动时和运动后血乳酸动力学的特点求出每个运动员的乳酸阈值,称为个体无氧阈。个体无氧阈值范围为2.1~4.6 mmol/L,更能准确反映机体有氧代谢能力。个体无氧阈强度训练,可以因人而异地发展机体的有氧代谢能力。
一般认为,血乳酸浓度的大小是反映活动肌肉内乳酸生成、血液间乳酸弥散率和消除率相差的结果,当血乳酸的消除率等于生成和弥散率时,这时的运动强度可视为个体无氧阈强度值(t Em)。当强度增加时,血乳酸浓度增加,在运动结束时达到最大(A),但在运动后恢复期中血乳酸消除率增加而使血乳酸下降,在下降中存在一点与A平行相交的血乳酸值(B);在A与B间相连成平行线,从乳酸动力学变化可推算出个体无氧阈值,在B和血乳酸曲线切点处(C)(图4.8)。计算方法为:
dn(t)/dt=Mc·ΔC=Em
dn(t)/dt:弥散率
Mc:弥散常数
ΔC:运动肌与血液乳酸浓度的梯度
Em:弥散率
当最大弥散率=消除率时,即为个体无氧阈值,在运动后,弥散率函数逐渐降低,当消除量Em(t A-t B)时,等于弥散量。则
dn(t)/dt = Em+Mc·Em(t B-t A)
Mc为弥散常数,用1 mmol/L乳酸梯度弥散量的增量,即分钟-1(1/dt)代入上式:
dn(t)/dt=Em(t B-t Em)
Em(t B-t Em)是从B到乳酸弥散曲线上的切点,从而求出个体乳酸阈值,这个值不受测定负荷强度的影响。
在测定个体无氧阈时,必须采用逐级递增负荷,用不同功率计或在运动场上的取血时间见表4.3。
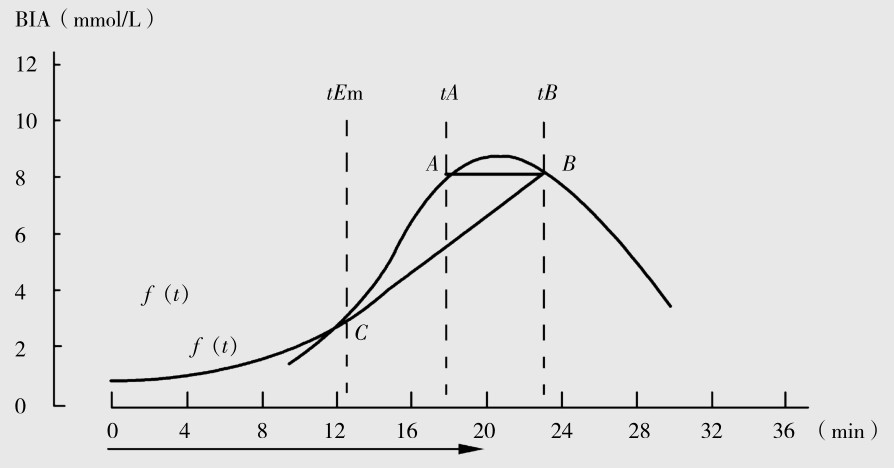
图4.8 递增负荷运动时血乳酸动力学
(引自Ste g mann, 1981)
表4.3 递增负荷测定个体无氧阈的方法
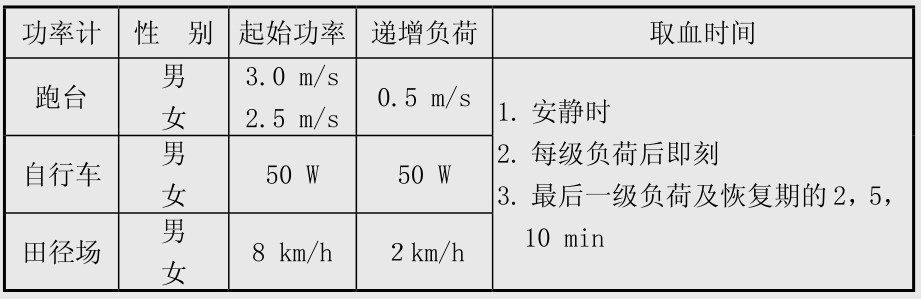
将负荷数值和时间作横坐标值,以各血乳酸值为纵坐标值,用曲线板平滑坐标上各血乳酸值的点,绘出血乳酸动力学变化曲线,从A, B值求出C点值,即为个体无氧阈。
知识链接
乳酸阈在耐力训练中的应用
1.长跑和超长距离跑耐力训练
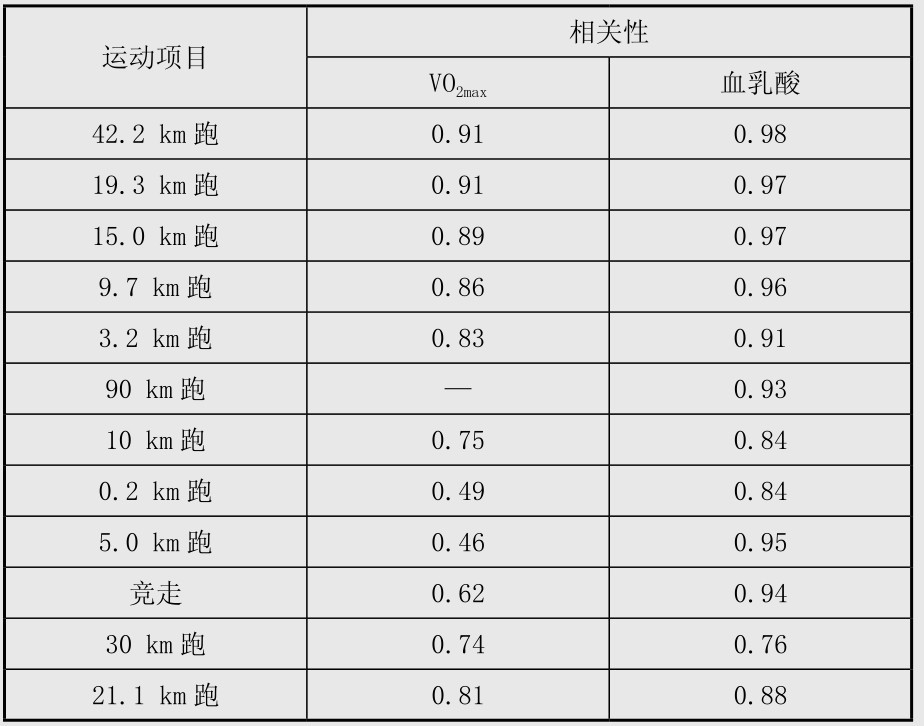
在耐力训练中,血乳酸是评定耐力的敏感指标,在训练开始时,提高耐力和VO2max、心率的相关性较高,但随着训练水平提高,用血乳酸来评定时灵敏度就更高了。一般人在耐力训练4~16周后,VO2max提高7%,血乳酸变化为16%。在优秀运动员中,训练14周后VO2max没有变化,但血乳酸变化为5%,其他受试者的变化也一致。可见,血乳酸指标是比较敏感的。血乳酸和耐力的相关性比最大摄氧量高(表4.4),而且血乳酸指标还有它的优越性,采血后可现场报告结果,利于教练员掌握训练过程。故当前血乳酸在评价耐力训练负荷强度和专项训练要求中是最常用的指标。
表4.4 各项耐力性运动时血乳酸和最大摄氧量(VO2max)和耐力的相关性
(引自Gledhill,1992)
2. 乳酸阈与其他指标的结合应用
血乳酸积累起点(OBLA)和体力感知度(ratio of perceived exertion,RPE)相结合:在进行有氧代谢训练时,在运动场上可将血乳酸测定和主观感觉结合起来掌握运动强度,当血乳酸超过4 mmol/L时,受试者的主观感觉又达到体力感知度值6时,说明运动强度过大(图4.9、图4.10)。
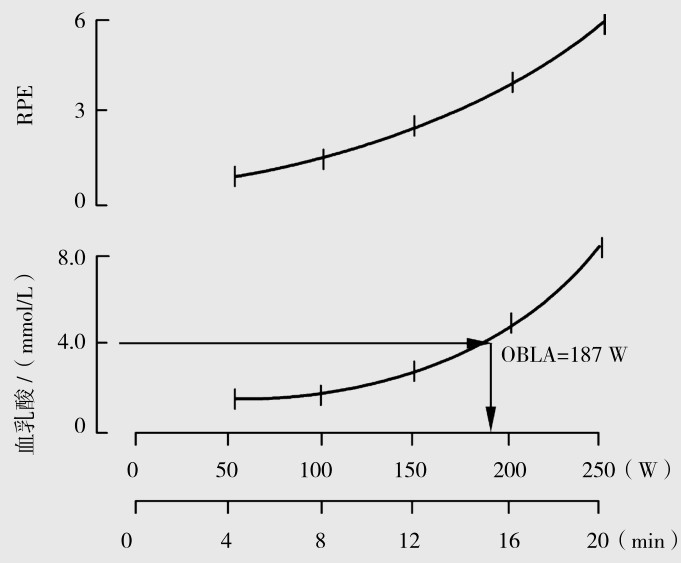
图4.9 递增强度负荷时OBLA和RPE的关系

图4.10 体力感知度(RPE)示意图
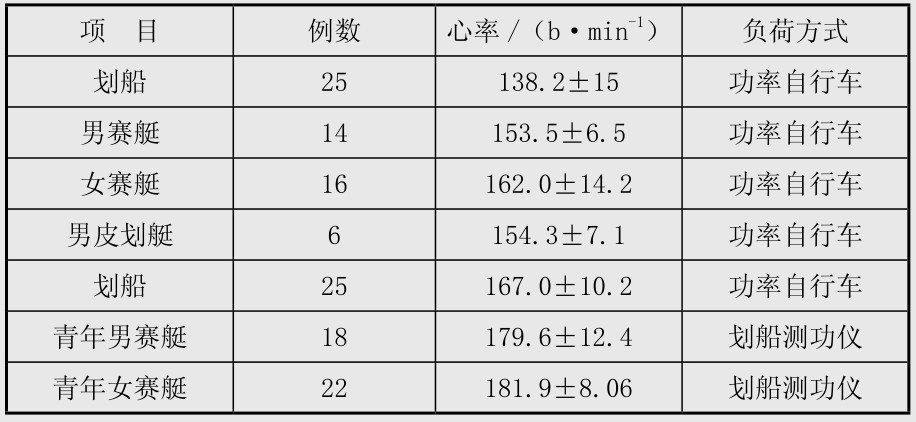
血乳酸和心率相结合:血乳酸在4 mmol/L时的心率简称小时4,小时4训练是指运动强度达血乳酸4 mmol/L时的心率。因此,在测定乳酸阈的同时,要测定心率,以找出小时4的值。在训练课中以心率来掌握血乳酸4 mmol/L时的训练强度,就不用在每次训练课中都测血乳酸。但经过一个训练阶段后便要重新测定,以了解训练对乳酸阈的影响来评定训练效果。如运动员在训练季度开始时的小时可能是160次/min,经过一个阶段训练后,可改变为170次/min。表4.5是我国划船运动员的小时4值。3.游泳的耐力训练
表4.5 划船运动员LAT(血乳酸达4 mmol/L)时的心率
在耐力训练中监测有氧代谢强度时,血乳酸非常重要,因为游的速度太快时,体内无氧代谢所占比例加大。血乳酸增多,对有氧代谢能力训练效果就差;但游速太慢,也收不到预期的训练效果。不同运动员的身体代谢能力也不同,不能在训练中用同一强度进行训练。如图4.11所示,A,B两名运动员的有氧代谢能力不同。A和B的V4速度(V4pace)(即血乳酸为4 mmol/L时的速度)不同,A要赶上B和V4速度,血乳酸值则需要7 mmol/L。如按这强度训练,其强度会过大而不能坚持训练。因此,应当测定个体无氧阈,但精确测定个体无氧阈有一定困难,Mader等提出了一个粗略划定个体无氧阈的办法,就是根据一般测定V4速度的方法,因为有氧代谢能力越好,V4速度越快,这时血乳酸值就越低。
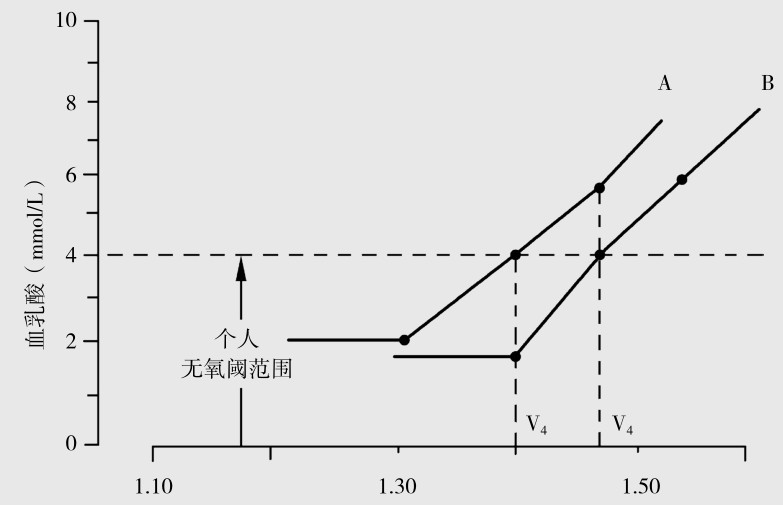
图4.11 A,B两运动员血乳酸曲线比较
在100,200,400 m游泳训练中,如间歇时间为10 s,30 s时,连续游百米的平均时间,根据血乳酸数值可分4种强度安排(表4.6)。
表4.6 游泳耐力训练的强度与成绩的换算
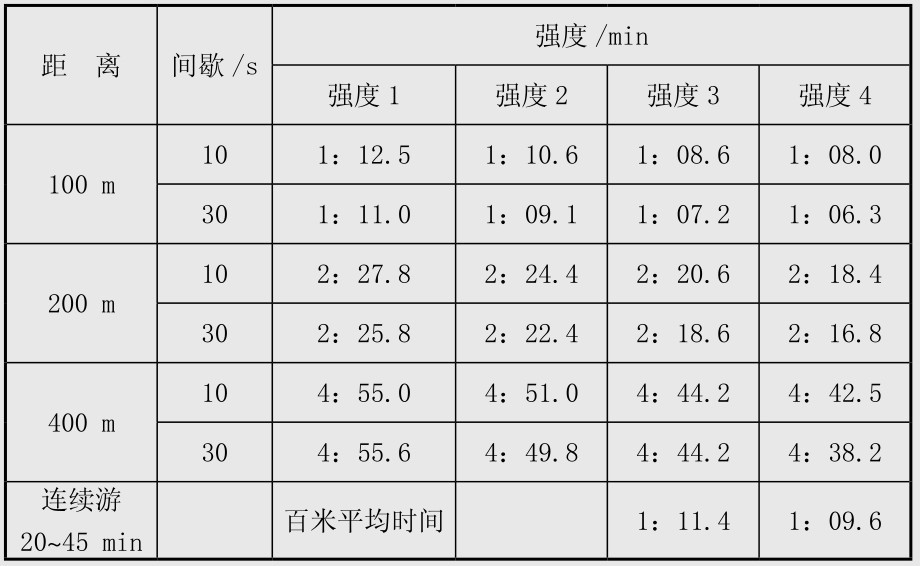
强度1:低于中等强度,能量供应主要来自脂肪酸,因此血乳酸值为1~2.5 mmol/L,主要用于乳酸无氧阈强度训练后的恢复。
强度2:中等速度,能量供应主要依靠脂肪酸和糖原,训练时血乳酸值为1.5~2.5 mmol/L。这种强度主要用于3 000~5 000 m游泳或长距离连续游泳。
强度3:乳酸阈速度,能量供应主要是糖原,训练时血乳酸为2~5 mol/L。这是提高有氧耐力最有效的强度。
强度4:强度较大,能量完全依靠糖原酵解供应。血乳酸值要求比乳酸阈高1~1.5 mmol/L,这种强度用于负荷总量较小(2 000~2 500 m)、休息较长(30~45 s)的练习,这种强度对有多年训练和水平较高的运动员较适宜。
4.自行车运动员的耐力训练
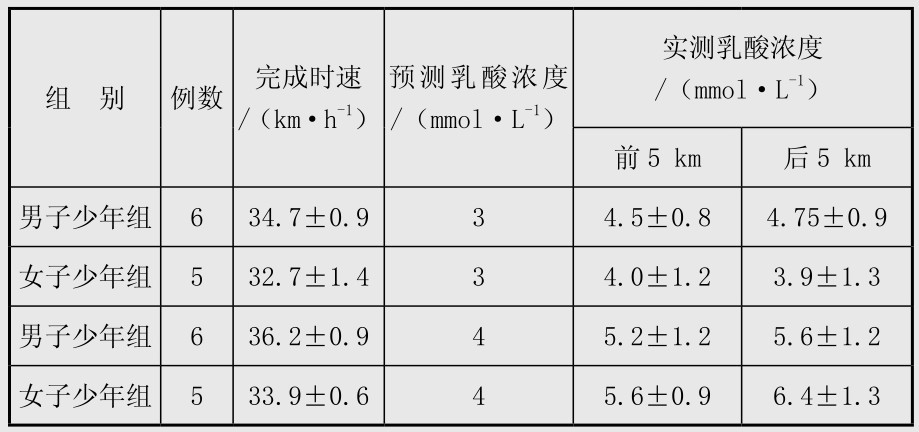
在自行车耐力训练中,先求出4 mmol/L乳酸阈的速度,然后以相当于3 mmol/L和4 mmol/L预测速度骑行10 km,总时间为17~18.5 min,两次骑行间歇1 h,每次试验前,后5 km末停30~60 s,采耳血测血乳酸。教练员按预计时速给运动员报时,实际完成时速与预计时速相差少于±0.7%,运动后血乳酸值见表4.7,说明以血乳酸3 mmol/L时的速度匀速骑行后,男女运动员的实际血乳酸范围为3.9~4.75 mmol/L,前后5 km相差不超过0.25 mmol/L。但以4 mmol/L乳酸阈速度运动时,前后5 km的血乳酸相差0.8 mmol/L。因此,建议以相当于3 mmol/L的乳酸速度作为场地自行车运动一段耐力训练的有效强度,以4 mmol/L或大于4 mmol/L的速度作为有氧无氧混合训练的强度。
表4.7 低于和相当于4 mmol/L乳酸阈速度的测验
(三)了解运动项目中不同训练手段与能量供应系统的关系是制订训练方法、选择训练手段的基础依据之一
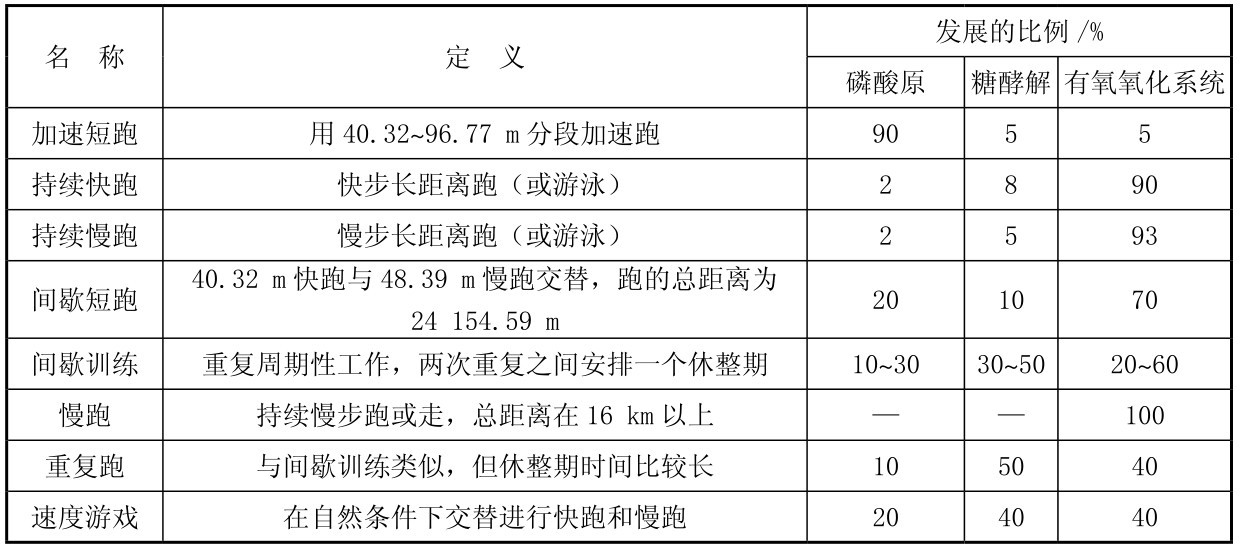
通过符合项目特点的训练手段,可以达到发展专项素质能力的供能系统的目的。表4.8中各种跑的训练手段与发展供能系统的比例,再进一步结合项目和运动员的个体特点,通过总结可以作为训练计划中选择训练手段的重要依据。
表4.8 各种跑的训练方法和发展供能系统的比例
知识拓展
自行车项目的专项供能
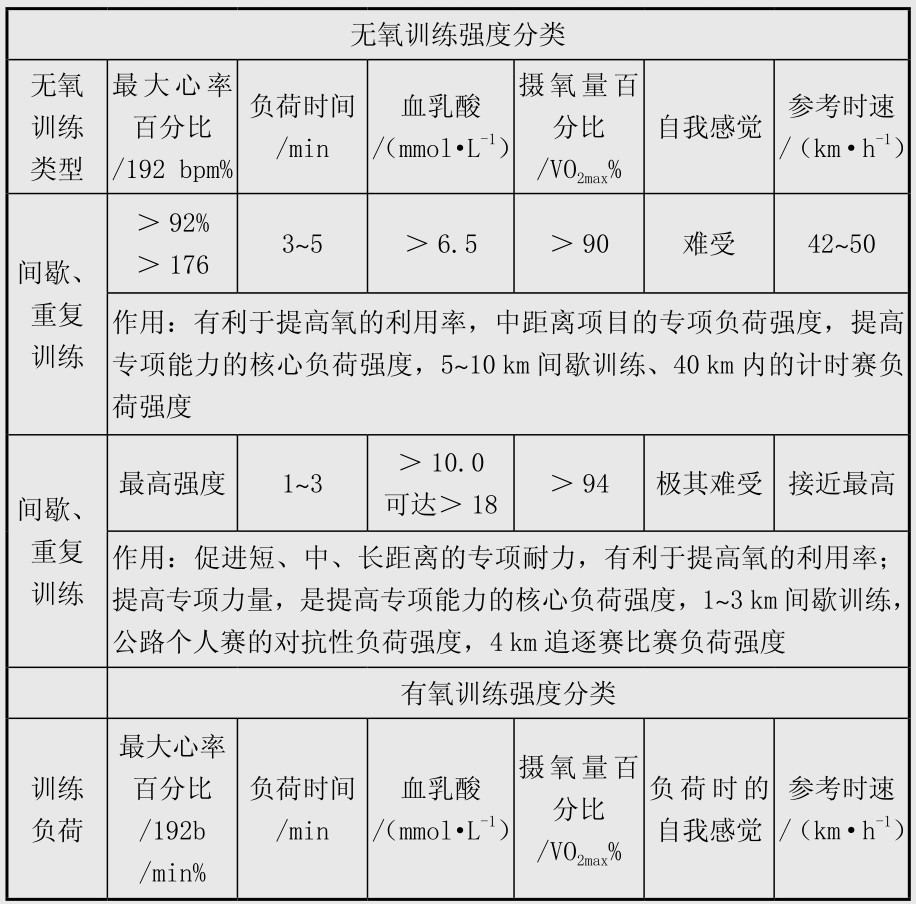
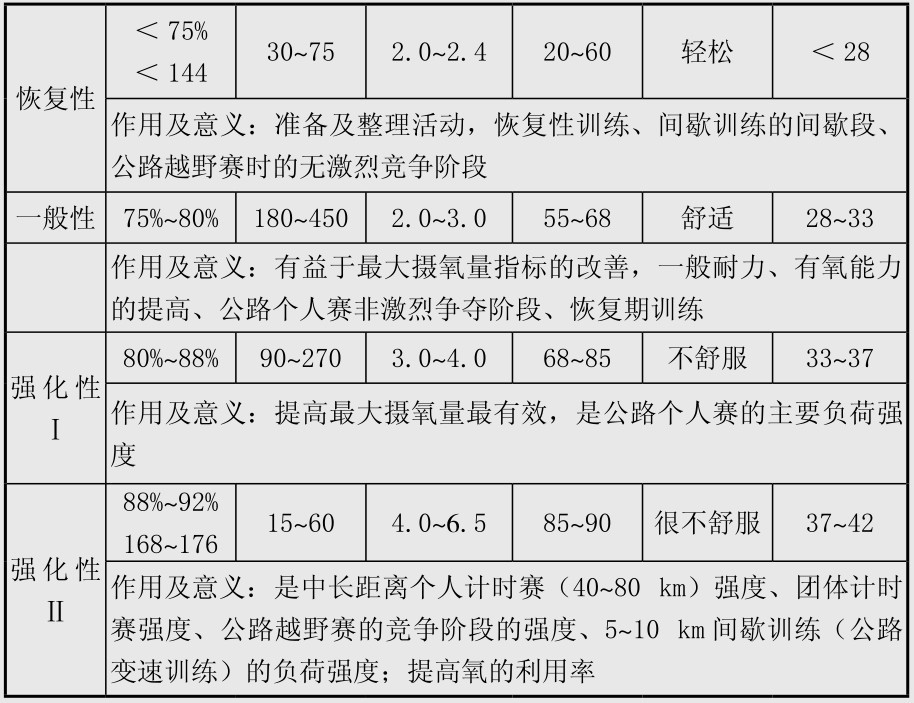
场地自行车项目比赛中能量系统的专项供能特点见表4.9。表中项目中不同能量系统的供能比例数据是基于运动员在比赛中动用其最大的有氧和无氧能力,这些数据在发表的文献中出入比较大,在男子4 km个人追逐赛也有学者认为无氧供能20%~30%,有氧供能占70%~80%,女子3 km个人追逐赛中无氧供能25%,有氧供能75%;在小于4 km的项目中出入更大,如有学者认为1 km项目中无氧与有氧供能分别是70%和30%,也有学者认为分别是95%和5%,最新的研究则认为分别是50%与50%;作者倾向于认为男子1 km中有氧与无氧供能各占一半左右,这些有待于今后进一步研究。表4.10是Burke发表的研究结果,与表4.9有出入,在这里也列出,供比较参考,表4.11是自行车无氧训练强度分类和有氧训练强度分类,可以作为实际训练中选择训练方法的参考。场地计分赛中比赛后男女运动员的血乳酸值一般均大于10 mmol/L,最高可达到15~16 mmol/L,实际上在世界高水平的场地计分赛中运动员一直保持较高的骑行速度,在冲刺圈前1~2圈即开始高速骑行,更重要的,运动员进行的战术脱圈以求得得分的绝对优势更需要运动员有非常强的无氧糖酵解供能能力,因此糖酵解无氧供能在场地计分赛中取得胜利非常重要。男子奥林匹克竞速赛中,3名运动员的能量供应特点完全不同,这对于比赛中合理安排运动员的次序非常重要。训练与比赛中磷酸原供能和糖酵解供能与运动员神经活动特点和肌纤维类型密切相关,一定程度上与遗传有关。
表4.9 场地自行车主要项目比赛中能量系统分配比例
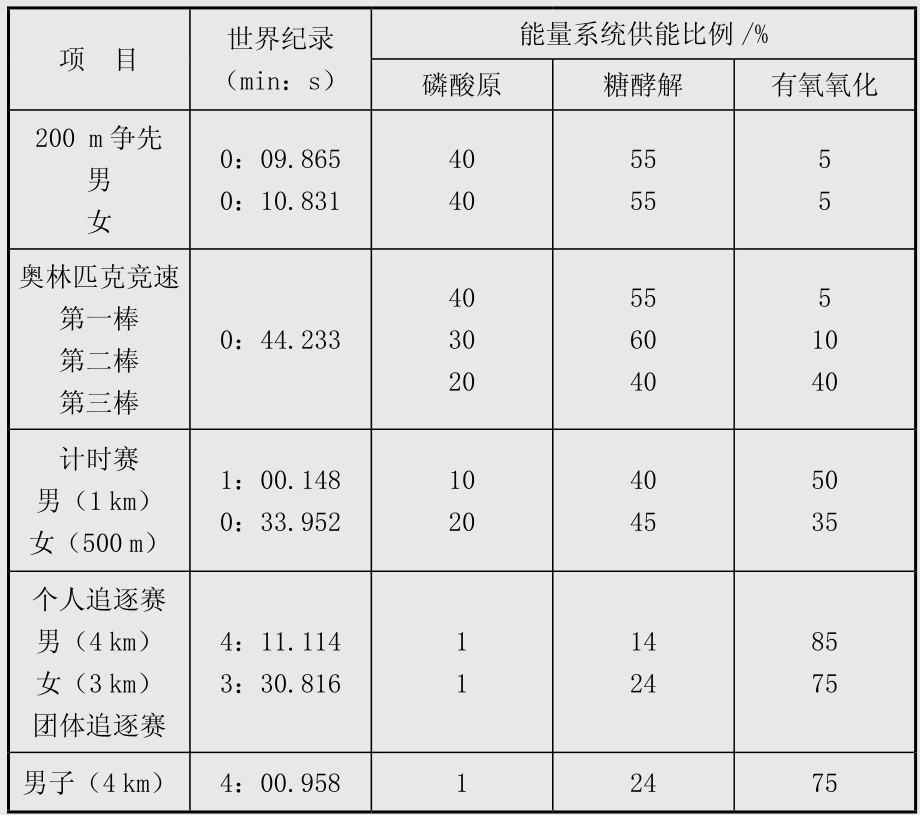
(引自Crai g等,sports medicine,2001:31(7):458)
表4.10 不同距离自行车比赛的主要能量系统供能比例
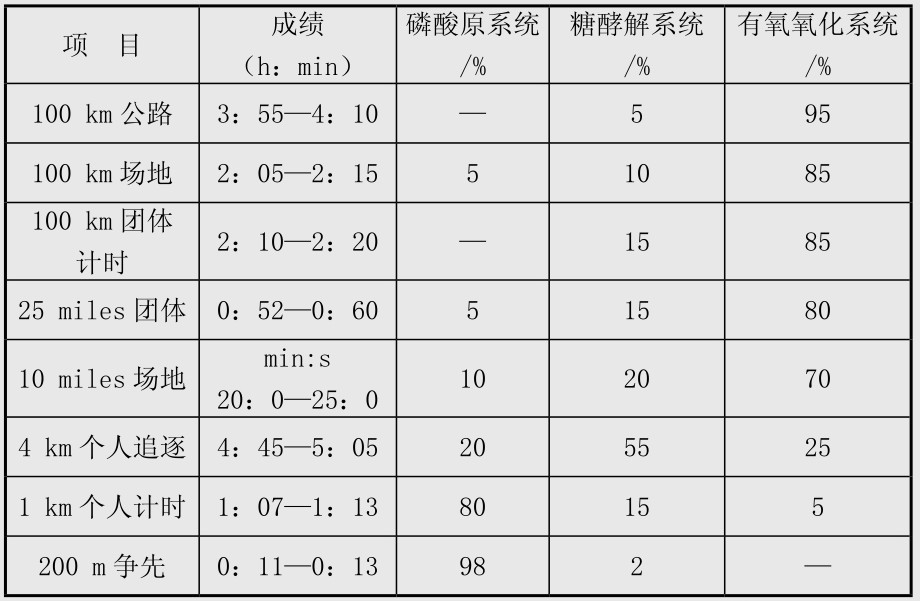
(引自Burke,1986)
表4.11 自行车无氧训练强度分类和有氧训练强度分类
续表
一、运动疲劳的概念
1982年第五届国际运动生化会议上将运动疲劳定义为:“身体机能的生理过程不能持续在特定水平和/或身体不能维持预定的运动强度。”
这个定义反映了运动的特点,主要表现在:
①在研究运动疲劳时,要将身体各组织器官的机能水平和运动能力结合起来分析疲劳发生和发展的规律。
②评定运动疲劳要将生理生化指标——心率、最大摄氧量、血乳酸、血尿素、血红蛋白等和运动能力例如在功率计上的输出功率、在运动场上的成绩结合起来。
③运动疲劳应注意其专项特点,如进行100 m跑和马拉松跑都存在不能维持预定运动强度(疲劳)的问题,体内生理生化变化不同,和劳动疲劳有根本的区别。
2001年Gibson A.S.C等总结了20世纪90年代以来运动疲劳的神经机理,提出了中枢疲劳和外周疲劳的区别,认为外周疲劳可定义为“在神经驱动没有变化或增强时骨骼动作电位减弱,或肌纤维蛋白在肌肉收缩时横桥循环障碍或兴奋收缩耦联减弱使肌肉力量下降”;中枢疲劳定义为“神经驱动或神经调控减弱导致力量生成或肌张力下降”。
二、运动疲劳时身体的生物化学变化
1.运动疲劳时能量物质储备的变化
①如短时间(短于20 s)运动疲劳的能源物质消耗以磷酸肌酸为主,下降可达95%以上。
②在长时间运动(1~2 h)时,以肌糖原消耗为主,消耗可达储量的90%以上。
③脂肪在运动时消耗增多,但体脂的储量减少,不是疲劳的因素(表4.12)。
表4.12 运动时人体能源储备变化与导致疲劳的主要因素
注:↓:下降 ≈:接近
(引自冯炜权,1999)
2.运动疲劳时出现能量代谢过程紊乱,加重运动疲劳(表4.13)
①长时间运动疲劳时能量代谢产物氨明显增加,并进入血液,导致物质和能量代谢调节紊乱。血液和组织中的氨生成影响肌肉工作能力;在脑组织中氨生成增多会出现氨中毒症状,表现为运动平衡失调,严重时引起肌肉痉挛;血氨升高也加重肝、肾的负担。
②短时间高强度运动中肌肉内乳酸的大量生成使肌肉中p H值下降,抑制糖代谢中的磷酸果糖激酶活性,使糖酵解供能受阻。
表4.13 运动疲劳时能量代谢过程紊乱

注:BCAA:支链氨基酸,AMP:一磷酸腺苷。
(引自冯炜权,1999)
3.运动疲劳时中枢产生的生化变化导致兴奋抑制失调,并导致脑异常症候群
在运动疲劳时,中枢出现ATP减少,克-氨基丁酸、儿茶酚胺增加,血糖下降等,都会引起兴奋抑制失调。
在运动疲劳时,色氨酸进入脑中过多,生成5-羟色胺,造成困倦、嗜睡、食欲减退;肌肉ATP大量消耗,导致氨生成,氨会引起脑中毒症状,如运动平衡失调等。
4.运动疲劳时出现内分泌调节紊乱
运动疲劳时,内分泌调节受到影响,并会短时间发生紊乱。目前研究较多的是下丘脑—垂体—肾上腺轴和下丘脑—垂体—性腺轴激素在运动疲劳时发生的紊乱,如图4.12所示。
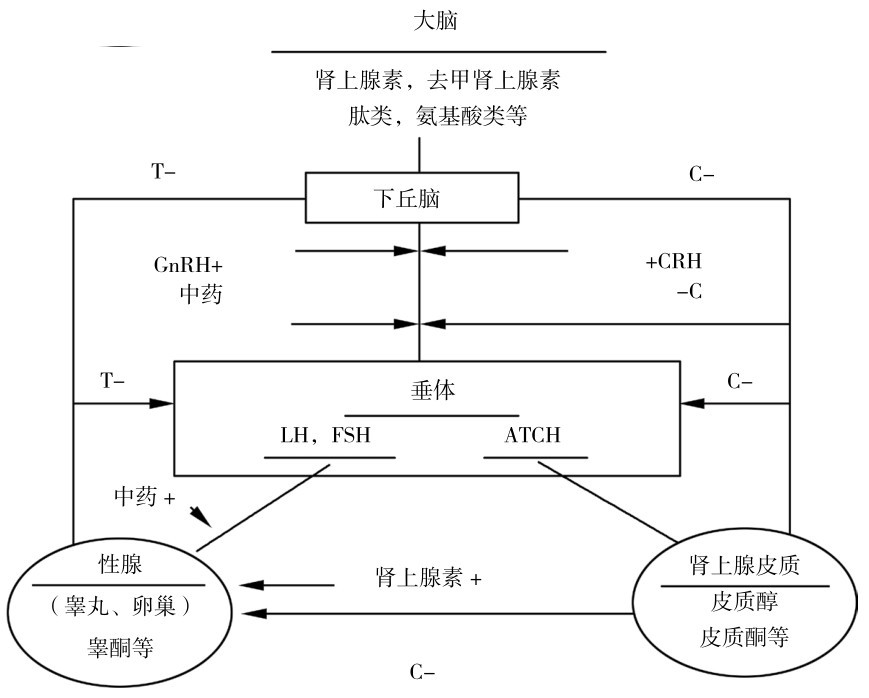
图4.12 运动疲劳与下丘脑—垂体—性腺/下丘脑—垂体—肾上腺皮质轴调节关系
(引自冯炜权,2000)
运动应激的一般规律是首先引起下丘脑—垂体—肾上腺轴活动加强,人体血中皮质醇明显上升,加速分解代谢,以适应运动的代谢需求,同时性腺分泌雄激素减少,合成代谢减弱。运动后,皮质醇分泌减少,雄性激素分泌增多,提高合成代谢,加速身体恢复,从而提高运动能力。
短时间运动可引起血清睾酮、皮质醇、催乳素、生长激素和肾上腺素明显升高,血睾酮升高,主要原因在于运动引起肾上腺素增加使睾酮分泌增加。
在长时间运动中,运动负荷强度和量度过大时,皮质醇分泌持续增加,对下丘脑—垂体—性腺轴有广泛的抑制作用,对免疫系统也有抑制效应。处于疲劳状态时,睾酮分泌减少。长期大强度训练和一次长时间大强度训练或比赛(如马拉松跑)是造成下丘脑—垂体—性腺轴不同环节不同程度的功能抑制的原因,与运动训练的强度和时间有关,长期大强度训练使血皮质醇持续处于超过正常的高水平,对合成睾酮有抑制作用。
三、运动疲劳机理的生物化学基础
(一)产生运动疲劳的环节因素整体概况:神经-肌肉疲劳链
运动疲劳的因素十分复杂,其生化变化机理还在不断深入研究之中。在一次运动中和长期训练中,由于运动的强度与持续时间不同,有多种不同的生理过程甚至非生理的意识过程参与了疲劳的发生与调节,因此,运动性疲劳是运动特点决定的。总体来说可以归纳为:
①神经中枢对运动中人体主要器官发生的生理变化的控制与调节。
②运动中肌肉等外周器官的物质和能量代谢,以及机能的调节与适应性变化。
③肌肉等外周器官在运动中的机能变化产生的向中枢的反馈,中枢进行相应的反馈调节。
目前的研究,在运动疲劳的整体环节因素中提出了神经-肌肉疲劳链等学说,并对外周因素进行了较为深入的研究。如图4.13所示,在疲劳链中,一个或几个因素的发生和发展都可以影响肌肉功能而产生疲劳,如运动时肌肉乳酸、H+堆积,血糖下降、肌糖原几乎耗竭等都是运动疲劳链的重要因素。
(二)运动疲劳的机理
运动时肌肉疲劳的发生和发展明显依赖于运动时间、强度、运动性质、肌纤维组成、运动员体质水平和内环境等因素,故直接从运动类型特点分析其机理更符合实际。
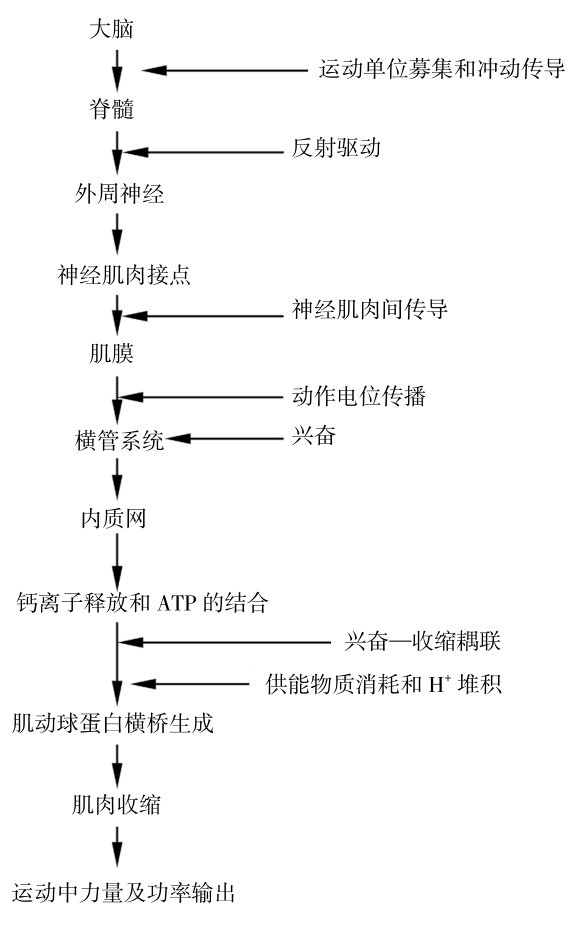
图4.13 肌肉收缩链的调控和肌肉疲劳的可能机理
(引自Maclaren和Edwards等,1989)
1.短时间高强度运动疲劳
主要从兴奋—收缩耦联、电位活动期和不应期、收缩期、疲劳动因的代谢能力、疲劳和电刺激频率等方面阐述肌肉在短时间高强度运动时的疲劳特点。现简要介绍如下:
(1)兴奋—收缩耦联引起疲劳
如图4.14所示是肌细胞兴奋—收缩耦联的主要部位,可能是短时间高强度运动疲劳的部位。
疲劳时会因细胞外钾堆积和/或钠下降影响膜电位,动作电位下降。肌肉疲劳出现的位点包括肌纤维膜和t管膜的去极化,兴奋—收缩耦联的一般过程受损。短时间肌肉疲劳是最高速率的肌紧张下降,并引起细胞外Ca2+增加,从而降低肌浆网和Ca2+结合蛋白中Ca2+的重吸收,可导致肌紧张发生速率下降,这都影响兴奋—收缩耦联,从而影响肌肉力量。
(2)物质代谢能力引起疲劳
高强度运动时,能量需求大于个体的最大有氧代谢功率和需要高水平的无氧代谢,高强度运动疲劳的动因是ATP和CP下降,同时Pi(磷酸)、ADP、乳酸和[H+]增加。其间的关系是:①ATP和CP:ATP是肌球蛋白横桥连接产生力的能源,又是肌膜和t管Na/K泵维持正常机能的必需。加之,ATP是肌浆网ATP酶的基质,由此需求肌浆网中Ca2+的重吸收,这个过程被扰乱就可导致疲劳。
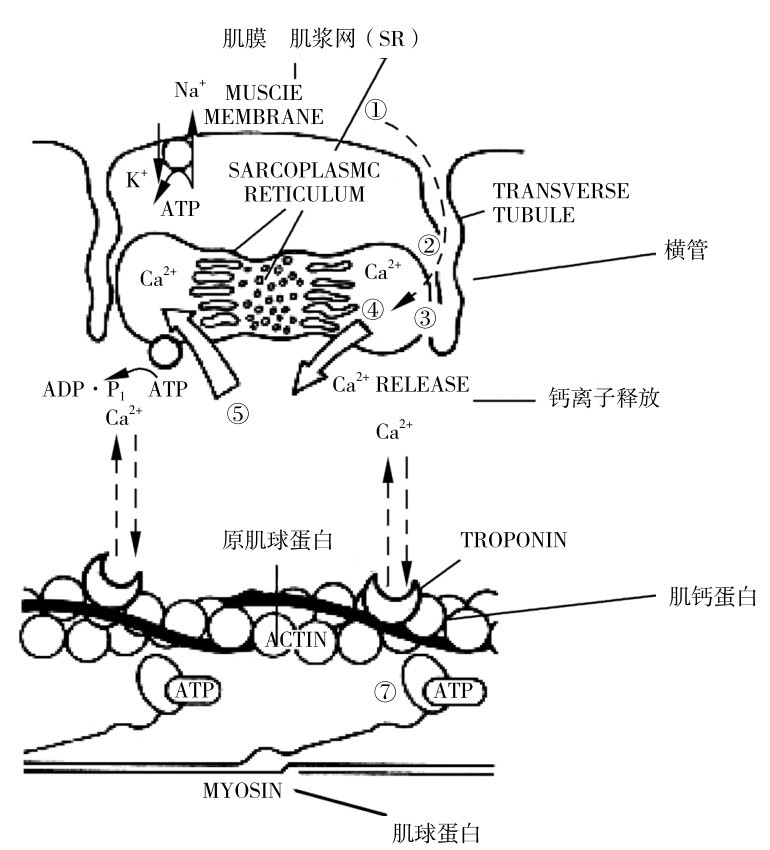
图4.14 肌肉细胞中兴奋—收缩耦联主要成分图示(Fitts,Metzger,1988)
注:图中数字可能是激烈运动的疲劳部位:①膜表面;②t管电荷运动;③t管电荷运动同SRCa2+耦联尚不明的机理;
④SRCa2+释放;⑤SRCa2+重吸收;⑥Ca2+结合肌钙蛋白;⑦ATP水解肌动球蛋白生成和横桥力量发生及循环速率。
在高强度运动时,ATP浓度下降不是直接引起疲劳的因素,因为在最大强度运动至疲劳时, ATP下降最多为原水平的40%,ATP不可能过多被消耗。CP被大量消耗(达储量95%),则在肌肉中浓度降低,可导致疲劳。线粒体中的ADP生成和ATP供给能量不断消耗,如ADP再磷酸化生成ATP速率减慢,也可导致疲劳。
②[H+]是运动疲劳的重要动因:在运动疲劳时,肌肉p H可降至6.33,这时导致疲劳的主要因素有:a.抑制磷酸果糖激酶活性而降低糖分解速率;b.竞争性抑制Ca2+结合肌钙蛋白C降低横桥活动性;c.抑制肌浆网ATP酶降低Ca2+重吸收和随后的Ca2+释放。
Fitts和Metzger总结运动类型疲劳时(1988)认为:短时间高强度运动时的主要机理可能有两个方面:a.快速肌肉收缩放松平衡中的兴奋—收缩耦联和Ca2+调节及非H+(或Pi)的中间干扰因素;b.较慢的变化包括肌肉收缩中一系列部位和H+或(Pi)代谢物及其产生的影响。
研究与应用
氢离子疲劳学说与碱性缓冲物质的补充
一、氢离子疲劳学说
运动性疲劳的原因中,氢离子学说是被普遍接受的学说之一。该学说认为,机体在高强度运动中由于氧供应不足,骨骼肌细胞需要通过糖酵解供能进行收缩运动,但在ATP产生的同时有大量乳酸在短时间内产生,而机体自身的缓冲系统难以完全缓冲急剧产生的乳酸,致使乳酸堆积,[H+]浓度增高,p H值下降。过多的H+可直接作用于肌原纤维,抑制肌动蛋白和肌球蛋白的结合,横桥不能形成,致使收缩能力下降。高浓度H+还可抑制钙离子从肌浆网释放,间接作用于兴奋—收缩耦联过程,影响收缩能力,导致疲劳的产生。高浓度H+还可抑制磷酸果糖激酶和磷酸化酶的活性,减慢糖酵解过程,使ATP和磷酸肌酸的合成减少,也会导致肌肉收缩能力下降。H+增多还可损害神经冲动的传导,从而影响骨骼肌的收缩能力。因此,大量乳酸引起的H+增多是肌肉收缩能力下降和疲劳产生的重要原因,如图4.15所示。
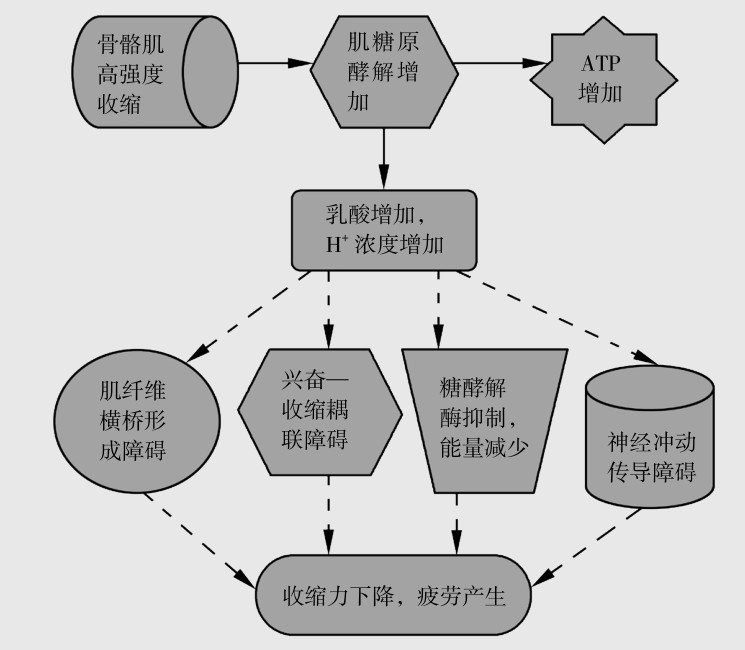
图4.15 氢离子疲劳学说示意图
高强度运动依赖于中枢神经系统对于Ⅱ型肌纤维的募集能力,乳酸堆积会使Ⅱ型肌纤维在过酸环境下受到影响,并在酸性持续下降的情况下疲劳,长时间或者重复组数的高强度训练会使Ⅱ型肌纤维疲劳,而一旦中枢神经系统不能募集足够的肌纤维去维持功率输出,就会表现出运动能力降低。
基于氢离子疲劳学说,多数资料表明,对于高强度、时间为45 s~7 min或更长的运动,不管是持续性运动还是间歇性运动,补充酸性缓冲剂可起到提高运动成绩的作用。目前最常用的缓冲剂是碳酸氢钠。补充碳酸氢钠可使血浆或细胞外液Na HCO3/H2CO3缓冲对比值升高,碱储备增加,清除酸性物质的能力增强,体液酸碱平衡和电解质平衡趋于稳定,进而促进能量的生成,增强肌肉收缩,延缓疲劳发生,提高运动成绩。有研究报告,运动前1 h服用0.4 g/kg体重的碳酸氢钠,可以显著提高受试者股四头肌的收缩能力,并减轻肌肉的疲劳程度,加快运动后恢复。
二、补充碳酸氢钠常规服用方法
1.服用剂量
补充碳酸氢钠的剂量范围为0.2~0.5 g/kg体重,但一般认为最低有效剂量为0.3 g/kg,最佳有效剂量为0.3~0.4 g/kg。有人总结,在0.1~0.5 g/kg剂量范围内,服用剂量越大,运动成绩提高的幅度就越大。如果碳酸氢钠剂量小于0.1 g/kg,则效果不明显。
2.服用形式
有胶囊、片剂、液体等。如果使用胶囊或片剂,应同时服用0.5~1 L水或饮料。也可与柠檬酸钠、磷酸钠等酸性缓冲剂联合使用,但剂量上要作调整。
3.服用时间
一般要求训练或赛前1~2 h内服用。曾有人让受试者连续5 d服用碳酸氢钠(0.5 g/kg体重)后,血浆p H值和碳酸钠浓度升高,60 s最大强度蹬车做功量和最大功率值明显增加。该研究表明在运动前5 d连续服用碳酸氢钠也有效果。
4.服用次数
一般建议一次性服用,或运动前数小时内多次、少量服用。多次、少量服用有可能减轻或避免胃肠不适等不良反应。
剂量越大,越有可能引起或加重不良反应。剂量过大,还可导致碱中毒,甚至有过量服用引起胃破裂的个案报告。长时期大剂量服用有可能引起心率失常、肌肉痉挛、情绪变化,如冷漠和易冲动等。因此,碳酸氢钠的服用剂量必须适合。
延伸阅读
短时间运动疲劳的决定性因素是什么?
Gibson和Noakes等(2001)总结了他们实验室和其他研究者的研究结果,在人体运动中,代谢性疲劳因素包括乳酸增加、肌细胞内p H下降、磷酸肌酸大大减少、肌纤维兴奋—收缩耦联减弱等,尽管这些因素可能通过作用于感觉神经传入对运动神经元和运动中枢产生影响,但都不是直接导致运动性疲劳发生的直接原因(Fitts,1994;Noakes,1998);在一次运动中,由于运动的强度与持续时间不同,有多种不同的生理过程甚至非生理的意识过程参与了疲劳的发生与调节,因此,运动性疲劳是运动特点决定的。
不论是一次的最大速度跑或多次间歇性最大速度跑训练中,力量下降与疲劳发生与能量物质的消耗和代谢物质堆积并没有紧密的相关关系,中枢神经控制机制是疲劳发生的主要决定因素,可能是外周肌肉感觉神经传入使运动的中枢策略发生适应性变化,是避免肌肉损伤的保护性反应。
2.长时间耐力运动疲劳
耐力运动疲劳的主要因素包括肌肉和肝脏糖原的消耗、血糖下降、脱水和体温上升。这些疲劳的因素在不同运动性质和不同环境都会有差异,如肌糖原的动用在25%VO2max强度运动时为0.3葡萄糖单位千克-1min-1,当负荷强度为100% VO2max时升至3.4葡萄糖单位千克-1min-1。当以70%VO2max强度运动至力竭时,肌糖原接近耗竭(消耗90%左右),可维持运动约90 min。因此肌糖原的储量和代谢能力是决定耐力的重要因素。脂肪酸虽是耐力运动的能源,但储量很大,不是疲劳的能量因素。在长时间游泳至力竭时,慢肌纤维中ATP酶活性变化不显著,收缩力下降不是由酶活性引起的。在长时间耐力运动时,体内糖储备和肌糖原大大减少是疲劳的原因。
(三)神经—内分泌(激素)—免疫—代谢物调节的运动疲劳网络
在运动疲劳研究中,也发现了在运动应激细胞的各种代谢变化与神经—内分泌—免疫调节网络存在明显的相互作用。根据这些成果,冯炜权在《运动性疲劳和恢复过程与运动能力的研究新进展》一文中,总结当时研究成果,初步提出了神经—内分泌(激素)—免疫—代谢物—调节疲劳网络(图4.16),从人整体的水平上用分子网络分析运动疲劳,更加整体地阐明了运动疲劳各种因素和环节的相互关系。
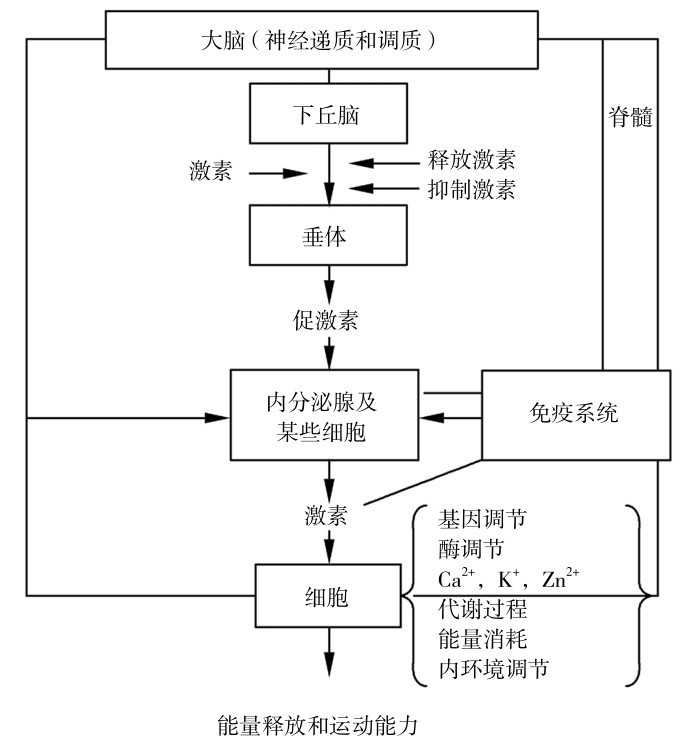
图4.16 神经—内分泌、免疫系统和代谢调节的运动疲劳网络
(引自冯炜权,1993)
热点讨论
运动中疲劳的中枢调节
近年来,大量的研究中根据对不同条件下的耐力运动训练中不同疲劳环节的分析,比较一致地认为中枢是耐力训练中最重要的疲劳调节环节。
①中枢调节运动单位的募集。在高强度耐力训练中,过去一般认为肌肉可利用的能量物质肌糖原、血糖、脂肪酸是疲劳的主要限制因素,但近年的研究发现,肌糖原消耗或糖类的氧化并不是疲劳的绝对因素,不论是一次的最大速度跑或多次间歇性最大速度跑训练中,力量下降与疲劳发生与能量物质的消耗和代谢物质堆积并没有紧密的相关关系。同时中枢也通过降低对外周肌肉的运动驱动下调肌肉的功率输出,运动单位募集减少。可能是外周肌肉感觉神经传入使运动的中枢策略发生适应性变化,是中枢神经系统传出的保护性反应,以预防肌肉损伤。
②中枢接受外周反馈,下调对肌肉的传出行兴奋。在长时间大强度耐力训练中,最大有氧能力并不是大强度耐力运动疲劳的主要决定因素(Noakes,1998),肌肉内的能量代谢不是一次最大强度的耐力训练中疲劳的第一限制因素。在长时间大强度耐力运动疲劳时,骨骼肌和心脏发出感觉传出信号到大脑中枢,下调运动皮层对运动肌肉的传出兴奋性,骨骼肌的工作能力降低,称为中枢控制疲劳理论。
一、力量和速度、耐力训练与骨骼肌供能物质代谢能力的适应过程
(一)耐力训练与骨骼肌供能物质代谢能力的适应过程
1.耐力训练动物模型对骨骼肌供能物质代谢能力适应过程的一般特点
在运动过程中,肌肉供能过程的主要途径是糖有氧代谢或糖酵解,脂肪氧化。在供能代谢中,耐力训练的影响和适应在肌肉细胞的基本特点是什么?
在耐力训练动物模型中,以慢频率电刺激兔肌肉的胫骨前肌,每秒刺激10次,每天2~4 h,在3天~10周内用活检法测定肌肉中酶活性的变化特点(图4.17、图4.18)。
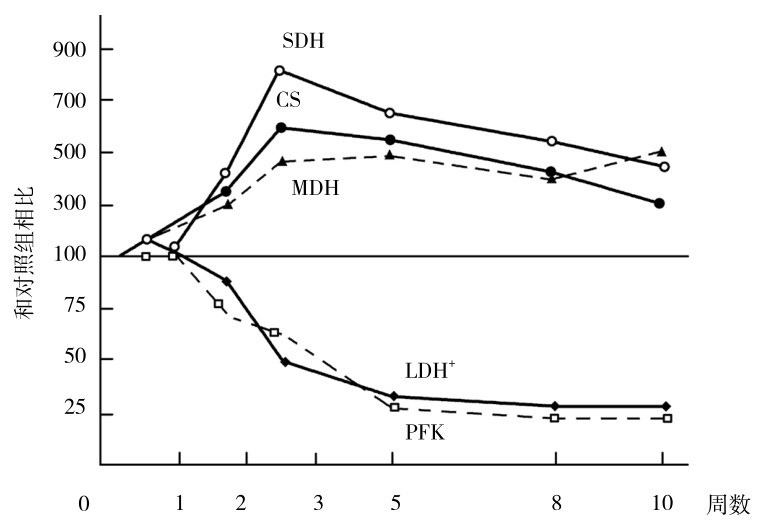
图4.17 慢频率电刺激兔胫骨前肌过程中肌肉中酶活性的变化(Henriksson等,1986)
注:SDH—琥珀酸脱氢酶 CS—柠檬酸合成酶 MDH—苹果酸脱氢酶 LDH—乳酸脱氢酶 PFK—磷酸果糖激酶
从图4.17和图4.18中可见,用慢频率长期电刺激模仿耐力运动时肌肉中酶适应过程中的有氧代谢酶活性,在第2~3周时酶活性提高最大,如琥珀酸脱氢酶、柠檬酸合成酶、苹果酸脱氢酶和己糖激酶等糖有氧代谢酶,以及脂肪有氧代谢酶3-羟乙酰辅酶A脱氢酶。其后逐渐下降,至第10周时虽仍有提高,但接近原水平。从图中可见,无氧代谢的糖代谢酶逐渐下降,乳酸脱氢酶、磷酸果糖激酶、磷酸化酶、磷酸葡萄糖变位酶等逐渐下降至原水平的25%。
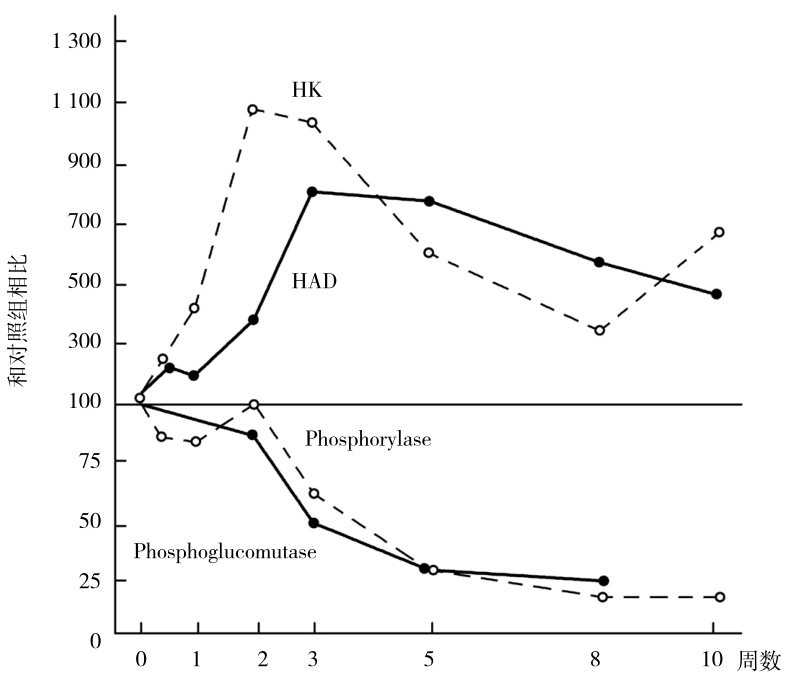
图4.18 慢频率电刺激兔胫骨前肌过程中肌肉中酶活性的变化
(Henriksson等,1986)
注:HK—己糖激酶 HAD—3-羟乙酰辅酶A脱氢酶 Phosphorylase—磷酸化酶 Phosphog lucomutase—磷酸葡萄糖变位酶
当大鼠在活动跑台上,以递增跑速10~60 m/min的强度,每周跑5 d,训练两个月后,在不同类型的肌纤维中,快肌氧化糖酵解纤维(typeⅡ A)、慢肌纤维(typeⅠ)和快肌糖酵解纤维(typeⅡ B)中有氧代谢能力变化的特点也不同(图4.19)。
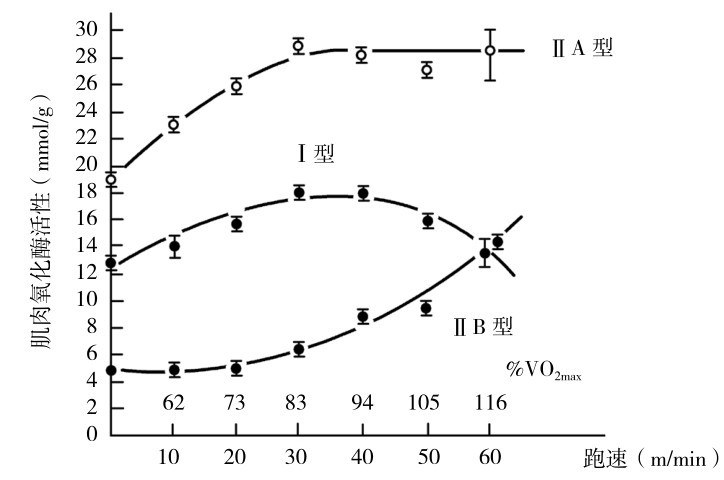
图4.19 大鼠在不同跑速运动时不同肌纤维氧化酶活性的变化
(Dudley,1982)
由上可知,肌肉耐力活动中供能物质代谢的适应过程特点是:
①与肌肉活动强度(收缩频率)相关。
②和固定的强度(频率)活动的总时间有关。从图4.19中可知,强度30 m/min(83% VO2max)时是肌肉氧化酶活性提高的最适强度,负荷强度加大时,肌肉氧化酶活性反而下降。在专项训练时,应当注意根据适应过程的特点,适时提高负荷强度或量度,依靠科学监控适时调整。
2.耐力训练对人体骨骼肌能物质代谢能力适应过程影响的一般特点
8名正常人以80%VO2max运动每天1 h,每周5 d,训练7周后,用活检法在股外肌取样,测定丙氨酸氨基转移酶(AAT)和柠檬酸合成酶(CS)的活性。如图4.20所示,7周自行车耐力性训练后AAT和CS活性显著增加,同时测定其和酶促代谢的相关产物浓度,从表4.14可知,耐力训练7周后,运动后肌肉中通过AAT酶促的有关代谢物(乳酸、丙酮酸)浓度相对减少,与CS相关的柠檬酸明显上升。
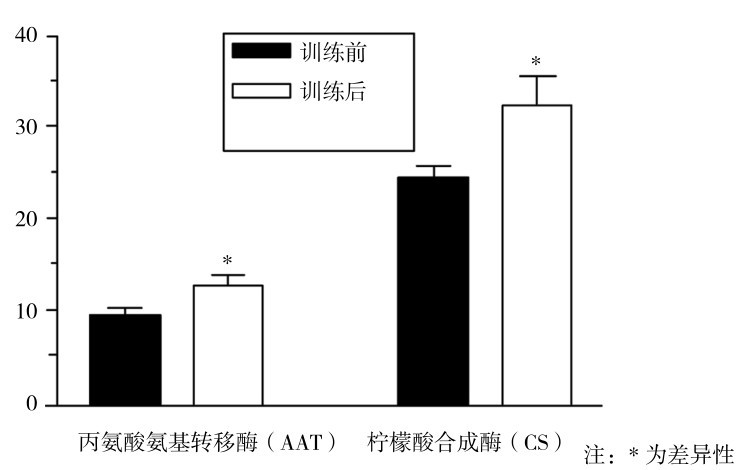
图4.20 自行车耐力训练7周前后肌肉AAT,CS活性明显升高
(Krista等,2004)
表4.14 自行车耐力训练7周前后肌肉中的代谢物(µmol/kg干肌)
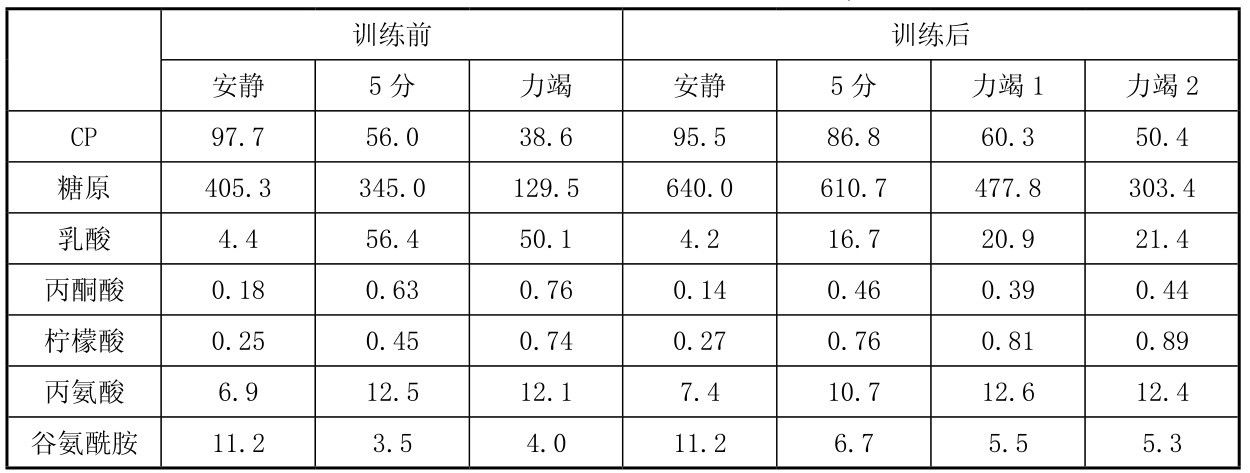
(摘自Krista等,2004)
3.停训对人体骨骼肌能物质代谢能力适应过程的影响
未受训练者在自行车功率计上以80%VO2max强度每天运动40 min,每周4 d,训练8~10周。在训练过程中和停训后2,4,6周,分析其有氧代谢酶,包括琥珀酸脱氢酶、细胞色素C氧化酶和最大摄氧量的变化(图4.21)。有氧代谢酶活性变化幅度大于最大摄氧量(VO2max)。而在停训后酶活性下降较快,而VO2max下降速度平缓。
动物和人体的耐力性训练能提高机体的有氧代谢能力,而无氧代谢能力反而降低,对训练的适应存在专项性。当停止训练时,训练的生化效果便很快消退。在相关的供能物质代谢中,肌肉对耐力训练适应的指标归纳见表4.15,供进行训练监控时应用。
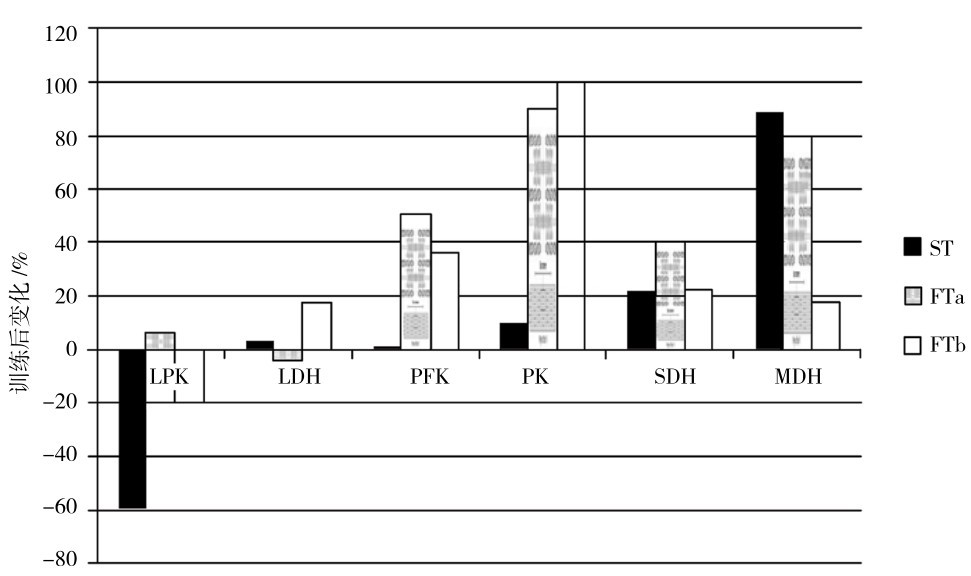
图4.21 速度训练后肌肉酶活性的变化(Takckura & Yoshioka 1990)
表4.15 耐力训练时肌肉的适应性变化
(引自Maughan R.等,1997)
(二)力量和速度训练对骨骼肌供能物质代谢能力的适应过程
1.无氧代谢酶活性升高,有氧代谢酶活性无明显变化
力量和速度训练时,不同肌纤维中酶活性的适应也不同,在FTa和FTb中的变化与ST中的变化不同(图4.22)。速度训练时,在FTb中,LDH(乳酸脱氢酶)、PFK(磷酸果糖激酶)、PK(丙酮酸激酶)明显上升,而有氧代谢酶SDH和MDH改善比ST少;在ST和FTb中,CK(CPK)反而下降。FTa中各种酶的变化特点居ST和FTb之间。
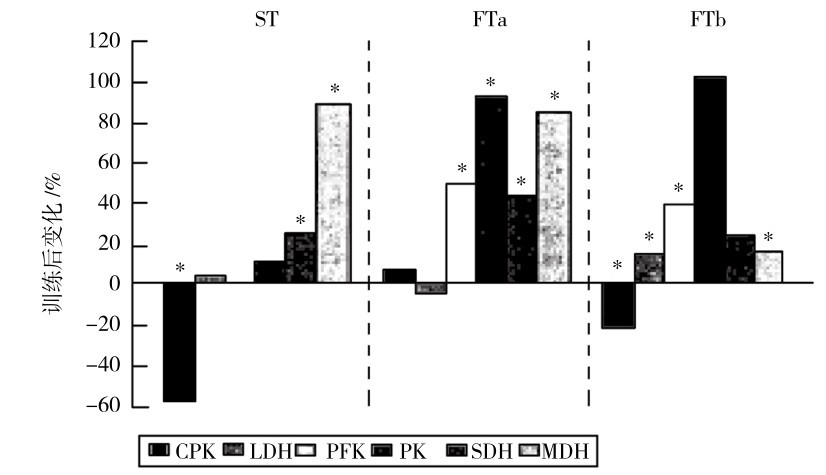
图4.22 速度训练后肌肉酶活性的变化
(Takckura & Yoshioka,1990)
2.CP和肌糖原是力量和速度练习时的主要供能物质
9名健身运动员在力量训练时,完成前后蹲起和座位蹬腿及负重坐凳起20组:每组重复6~12次,共30 min,测定股四头肌和血中代谢物的变化。从表4.16中可知,运动后肌肉和血中乳酸上升,肌肉中糖原明显下降(由690 mmol/kg干肌降至495 mmol/kg干肌),肌肉葡萄糖升高,血乳酸升高至11.7 mmol/L;而血中游离脂肪酸没有变化,但肌肉甘油三酯减少,3-磷酸甘油也增多,同时,肌肉中ATP,CP也下降,这都说明运动时ATP,CP,肌糖原和肌肉中的脂肪都参与运动时供能;但CP和肌糖原下降最多,是力量练习时的主要供能物质。
表4.16 健身运动员力量训练时肌肉和血液中代谢物的变化
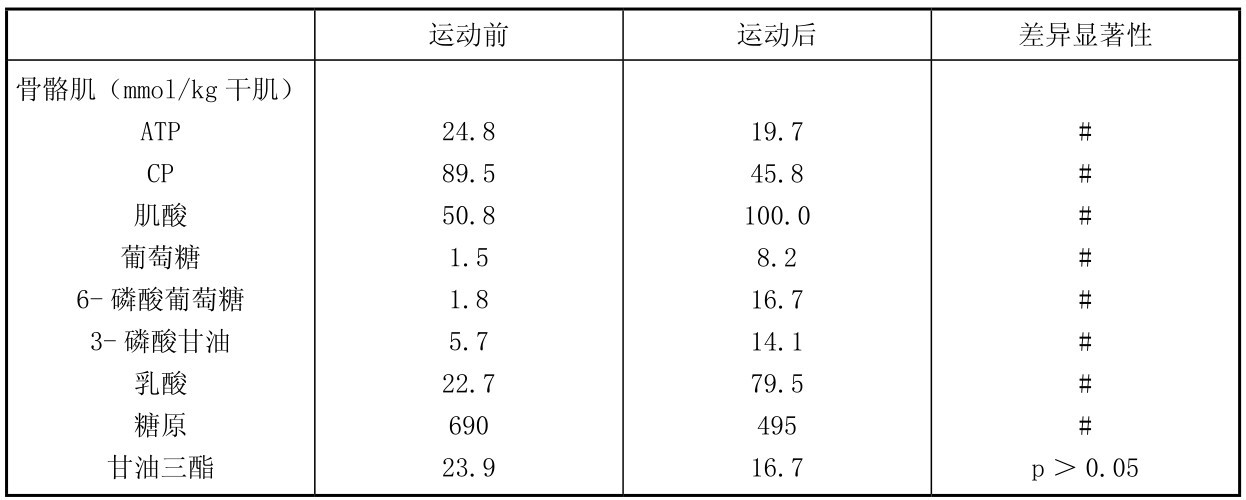
续表
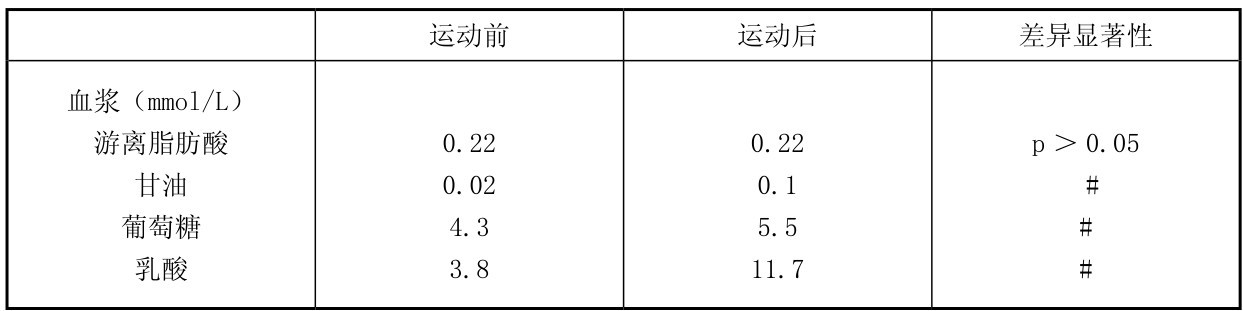
(引自Tesch等1986;Essen-Custavsson & Tesch,1990)
3.肌肉糖原大量消耗,快肌中糖原消耗大于慢肌
在运动员以70%~90%最大负荷完成大负荷抗阻力运动(20组,重复6~12次)后,在四头肌中可见,运动后15%的FT中糖原处于大量消耗;77%处于中等消耗水平;8%大量消耗;在ST中95%处于中等消耗水平,5%大量消耗(图4.23)。
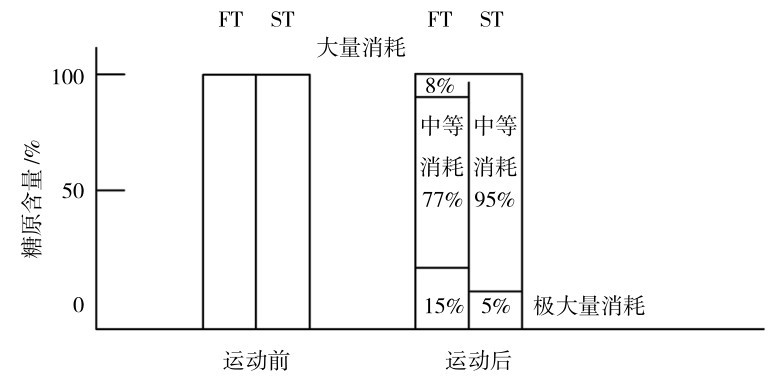
图4.23 肌肉中糖原在大强度力量训练时的变化
(引自Komi,1991)
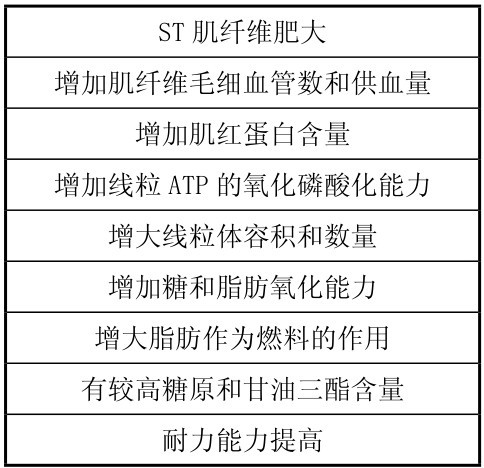
总的来说,力量和速度训练后,肌肉适应的主要表现见表4.17所示。
表4.17 力量和速度训练时肌肉的适应性变化
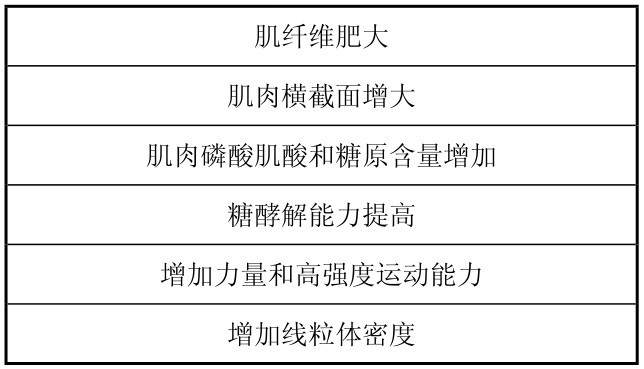
(引自Maughan R.等,1997)
二、运动后恢复过程的生物化学基础
(一)骨骼肌细胞的恢复与适应
运动训练对身体的适应过程应包括各器官、系统和其调节机理,但细胞适应是器官水平的基础。骨骼肌细胞是完成运动的基本单位,故不同运动都可引起肌细胞产生适应性变化。
①力量和速度训练使肌肉体积增大,力量训练的类型和方法不同,引起的适应也有不同特点。
抗阻力训练可优先使快肌纤维增大,在肌纤维类型组成不变下可使快肌纤维(Ⅱ型)在正常范围内增大90%。速度或力量训练可选择性地使快速糖分解纤维(typeⅡb)或快速有氧糖分解纤维(typeⅡa)变得肥大,如图4.24所示。
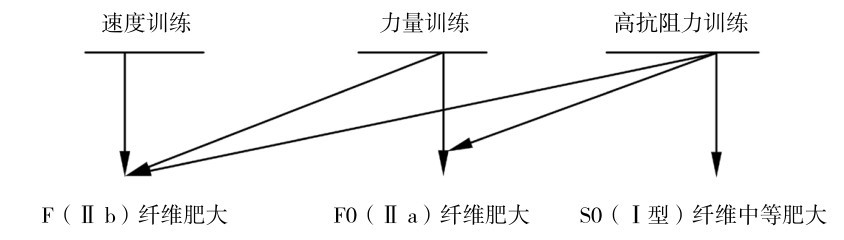
图4.24 不同训练类型对肌纤维肥大的效果
(引自Viru A.,Viru M.,2001)
②耐力训练时,主要增加慢肌纤维(Ⅰ型)的线粒体数量,有氧代谢的酶活性增加,有助于提高有氧代谢生成ATP的能力,从而提高耐力。
如图4.25所示是耐力训练的适应。
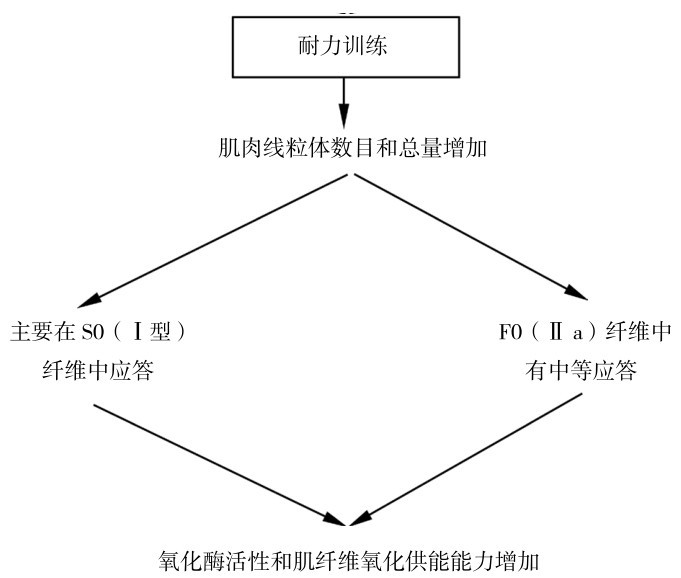
图4.25 耐力训练对肌纤维线粒体的适应特点
(引自Viru A.,Viru M.,2001)
(二)蛋白质合成的恢复与适应
1.蛋白质合成适应的一般规律
蛋白质是生命活动中完成各种机能活动的结构和活性物质。因此,运动后对蛋白质合成的适应主要为:
①运动中消耗的蛋白质修复和运动机能提高需求的蛋白质增加,从而使肌肉细胞结构蛋白增加。
②酶蛋白适应性增加,从而使某些酶数量增多而活性增加,有助于相应的代谢过程顺利或加速进行。训练课中激素的变化对代谢可起到放大和诱导的作用,因此,适应作用应达到结构变化增大和酶分子数量增加,其关系如图4.26所示。
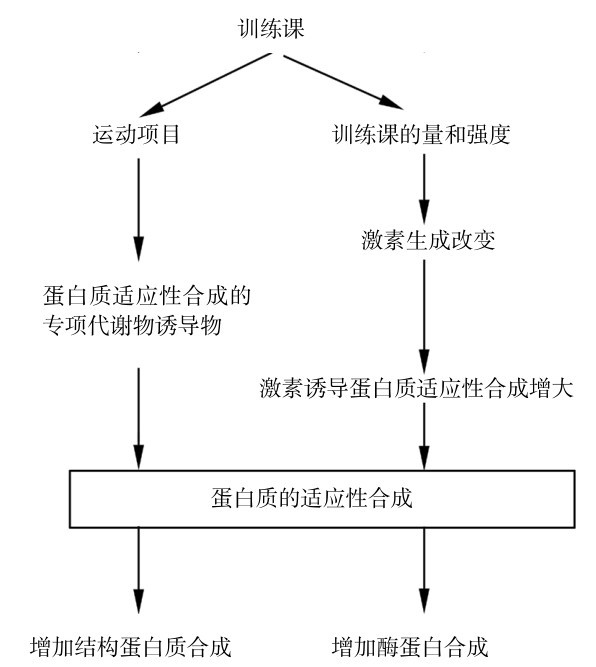
图4.26 运动训练和蛋白质合成的适应
(引自Viru A, Viru M.,2001)
2.酶蛋白合成的适应特点
酶蛋白合成的适应特点,从蛋白质合成的规律来看是一致的,但从运动的影响来看,则明显存在运动负荷性质的影响。
耐力训练与耐力相关的有氧代谢酶主要位于线粒体,提高线粒体酶活性的适应性可分为3个阶段,即:
①线粒体酶非补偿性适应阶段:耐力训练开始时,在线粒体有氧代谢中的酶活性升高的同时,细胞质中的酶活性也升高,表明在耐力训练初期线粒体中酶活性的提高不足以满足运动时的能量要求,而要动员糖酵解的无氧代谢参与供能。
②线粒体酶补偿的适应阶段:随着以同样强度继续训练,在线粒体三羧酸循环中的有氧代谢酶活性不断明显增加,而与糖酵解有关的无氧代谢酶活性下降,下降至接近训练前的水平。说明在这个阶段中,线粒体酶活性适应性升高达到能量需求,不需细胞质中无氧代谢酶升高来补偿。
③线粒体酶超补偿适应阶段:随着以同样负荷强度训练继续进行,线粒体酶活性处于稳定状态。
力量和速度训练中与力量相关的酶主要存在于细胞质中,酶活性的适应特点见第三章相关内容。
(三)能源物质的恢复与适应
详见第三章相关内容。
三、过度训练的生物化学基础
1.过度训练时糖代谢的变化特点
大强度耐力训练可导致骨骼肌和肝脏糖储备大量消耗,但在训练课之间合理安排休息和补充糖,可使糖原再合成至原水平。糖原储备极大量地慢性消耗不是导致过度训练本身的原因,但在运动时,糖过量消耗会引发肌肉疲劳。过度训练时糖储备大量消耗,而在训练课间休息时又没有足够的代偿。
2.过度训练时脂类代谢的变化特点
过度训练时,脂类代谢改变的特点为:①运动量增加提高氧化应激使胆固醇过氧化,导致较多的酯化胆固醇运转,在运动后,可见载脂蛋白A1(Apo A1)、载脂蛋白B(Apo B)增加,使酯化胆固醇和游离胆固醇比值增大。②进一步训练时,氧化应激导致甘油三酯/自由脂肪酸比值增加,血液中多不饱和脂肪酸增加,使载脂蛋白C3(Apo C3)浓度下降。③有助于脂肪氧化的瘦素(leptin)减少而减弱脂肪氧化等。这些变化可作为了解过度训练期的状况和研究参考。
3.过度训练时蛋白质代谢变化特点
在糖类和脂类出现上述变化后,继续进行高强度训练时,在过度训练者血浆中出现较高的氨基酸,而蛋血质降低,进一步可见氨基酸积累和3种蛋白质——α1-酸糖蛋白、Apo C3、免疫球蛋白G3(Ig3)下降,这说明蛋白质分解代谢加强,氨基酸从蛋白质中分解出来用于三羧酸循环能量生成的代谢中,在过度训练最后阶段时,糖—脂类代谢调节受阻导致更多地利用氨基酸,增加蛋白质分解以满足肌肉的能量需求,并由此导致大脑中色氨酸增加,5-羟色胺生成增多,造成中枢疲劳,从而改变免疫和/或内分泌调节。
过度训练过程中的代谢变化规律,如图4.27所示。
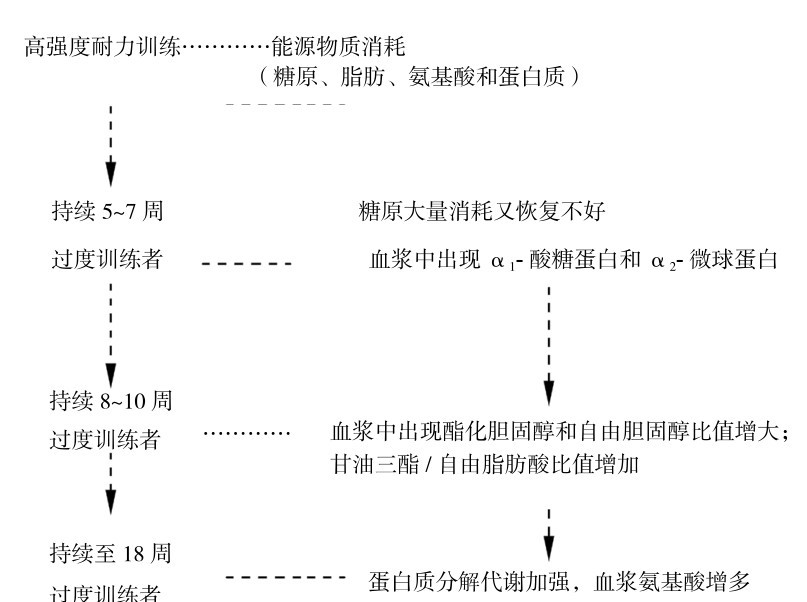
图4.27 耐力训练过度训练者的代谢过程变化规律
本章小结
首先界定了本章中力量和速度训练、耐力训练的含义,即力量训练包括发挥最大肌力以对抗阻力,或者肌肉以尽可能快的速度收缩产生快速的环节运动,运动的特点是运动时间较短,肌肉产生的绝对力量较大,包括常见的克服大阻力的运动如举重等典型力量项目,以及克服较大阻力的快速运动如短跑等典型的速度项目,运动中肌肉的能量供应以无氧代谢供能系统为主;耐力训练包括运动中肌肉产生的力量相对较小的运动,运动的特点是运动时间相对较长、环节运动的速度相对较慢,包括长跑、马拉松等典型的耐力项目,运动中肌肉的能量供应以有氧代谢供能系统为主。力量和速度、耐力训练中的物质代谢能量供应,可以简要地概括为运动时物质与能量代谢的两个过程和4个系统。具体情况主要取决于运动强度及运动强度的变化。
利用运动时物质和能量代谢的规律,提高训练的针对性和训练效果,无氧—低乳酸训练可以提高磷酸原供能系统供能能力素质的训练,最大乳酸训练法提高糖酵解供能系统供能能力素质的训练,常用于力量和速度项目运动员的专项训练。乳酸耐受力训练法也提高糖酵解供能系统供能能力素质的训练,乳酸阈(无氧阈)强度训练法提高有氧供能系统供能能力素质的训练,常用于耐力项目运动员的专项训练。
运动疲劳时身体的生物化学变化,主要包括运动疲劳时能量物质储备的变化,运动疲劳时出现能量代谢过程紊乱,运动疲劳时中枢产生的生化变化导致兴奋抑制失调,并导致脑异常症候群,运动疲劳时出现内分泌调节紊乱,特别是下丘脑—垂体—肾上腺轴和下丘脑—垂体—性腺轴激素在运动疲劳时发生的紊乱对疲劳的发生和发展产生较大影响。
力量和速度、耐力训练恢复和适应的生物化学基础,主要包括运动中骨骼肌供能物质代谢能力的适应,运动后骨骼肌细胞的恢复与适应、蛋白质合成的恢复与适应。不合理的适应则会发展为过度训练。训练中人体疲劳的特点、恢复和适应的规律,是制订训练方法和训练周期的基础。
练习题
一、名词解释
1.糖阈
2.乳酸阈
3.磷酸肌酸阈
4.无氧—低乳酸训练
5.无氧阈训练
6.最大乳酸训练
7.乳酸耐受力训练
8.运动疲劳
二、问答题
1.简述力量和速度、耐力训练中的能量供应体系。
2.以跑步为例,简述运动中肌肉内不同能源物质生成ATP速率与运动强度的关系。
3.如何利用运动时物质和能量代谢的规律,提高训练的针对性和训练效果?
4.举例说明如何进行乳酸耐受力训练。
5.举例说明如何进行最大乳酸训练。
6.简述无氧阈训练的原理。
7.举例说明如何进行无氧阈训练。
8.简述运动疲劳时身体的生物化学变化。
9.运动疲劳的特点是什么?
10.论述不同强度运动中能源物质的供能特点。
11.论述不同强度和时间运动后,能源物质恢复的规律。
12.力量和速度训练产生的肌肉的适应性变化有哪些?
13.耐力训练产生的肌肉的适应性变化有哪些?
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










