



第三章 竞技运动的能量和物质代谢基础
【学习任务】
人体在运动中,特别在竞技体育运动中,肌肉收缩需要的能量供应,需要分别在肌肉内通过生成能量的3个途径的生化过程来生成能量,分别为磷酸原系统、糖酵解系统和有氧氧化系统,分别与不同的运动强度直接相关,并在运动和长期训练中产生适应性调节了解运动训练实践中,肌肉能量供应方式具有连续性并与训练强度的关系;运动中伴随肌肉内的能量消耗,能量物质释放能量的一部分能量(约1/3)转化为肌肉的机械功,其余部分以热量形式释放,并随血液循环从运动肌肉到达身体其余部分,造成体温升高,运动中人体通过水盐代谢及其调节保持血液水分和体温相对稳定。
【学习目标】
1.掌握运动中物质代谢和能量代谢的基本概念。
2.理解磷酸原供能系统的基本过程。
3.理解糖酵解供能系统的基本过程。
4.理解糖、脂肪有氧供能系统的基本过程。
5.理解常见运动项目中能量供应的特点。
6.掌握运动中水盐代谢与体温调节的关系。
人体在运动中,特别是竞技体育运动中,肌肉大量消耗能量用于产生力量,进而带动人体各个环节产生运动。人体整体和各个环节运动中的速度不同,运动时间不同,对肌肉力量的要求差别很大,因而对能量的需求速率和需要量相差很大。肌肉直接利用的能量物质是三磷酸腺苷(ATP),由能量物质(主要是磷酸肌酸、糖、脂肪和氨基酸)通过代谢转化生成ATP(图3.1)。在人体肌肉内,存在3种不同类型的能量生成方式,分别利用不同的能量物质,用于分别满足相应的对肌肉力量的不同要求和总能量消耗需求,分别称为磷酸原(PCr)供能系统,糖酵解供能系统(前两者合称无氧代谢系统),糖、脂肪有氧代谢系统(图3.2)。
图3.1 不同强度负荷30 min运动时动用能量物质的分布
(引自谢敏豪等,《运动生物化学》,2008)

图3.2 不同速度跑主要的供能系统
(引自Don Maclaren,2012)
[想一想]
图3.1和图3.2分别表示了在不同运动强度下运动中能量的不同物质来源和不同能量的生成途径,说明了能量代谢是运动强度的决定因素,那么我们在学习中如何把具体的理论和运动相联系呢?
磷酸原是三磷酸腺苷(ATP)和磷酸肌酸(CP)的合称,是骨骼肌内重要的高能化合物。通过ATP和CP供应能量的代谢过程,称为磷酸原代谢供能系统,包括ATP,CP的分解与合成。ATP释放的能量可以直接被肌肉利用于肌肉收缩,也是所有生命活动中能量的直接来源,称为能量“通用货币”,CP和其他能量物质都要转化为ATP才能被肌肉利用。
一、ATP的作用和代谢
(一)ATP的化学结构
如图3.3所示为ATP的分子结构式,可以分解高能磷酸键释放能量,生成二磷酸腺苷(ADP)和一磷酸腺苷(AMP),同时,ADP可以接受其他能量物质(糖、脂肪、氨基酸等)分解产生的能量重新合成ATP,反应式为
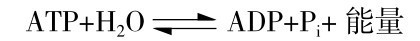
在肌肉收缩(ATP分解)和松弛(ADP再磷酸化合成ATP)循环中,维持肌肉持续收缩需要的能量。
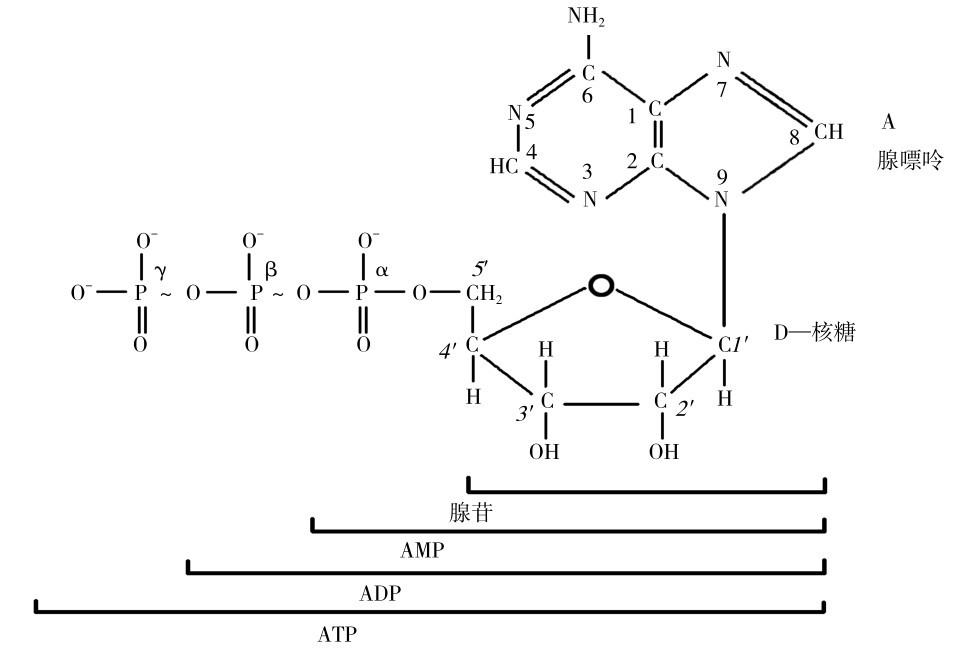
图3.3 ATP的分子结构
(二)ATP的作用和代谢
①ATP是肌肉收缩中直接的供能物质。ATP在肌肉中的储备量极少,是运动中的直接供能物质,但不是运动中的主要供能储备物质,其他供能储备物质转化为ATP才能被肌肉利用,有关知识在后面讲述的内容中。
②ADP再转化为ATP的速率很快。ATP分解后随即可以重新合成,运动中肌肉内ATP含量基本稳定。
③骨骼肌细胞内p H值下降(酸性增加)降低肌肉对ATP的利用率。
二、CP的作用和代谢
(一)CP的化学结构
如图3.4所示是磷酸肌酸(缩写为CP或PCr)的化学结构式,包括两个组成部分:一部分称为肌酸;另一部分称为磷酸。两部分之间通过高能的氮磷键连接。CP分解时这一高能化学键中的能量释放出来,用于ATP的合成。
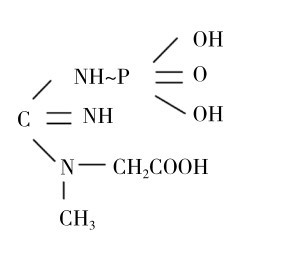
图3.4 CP的分子结构式
(二)CP的作用和代谢
1.分解释放能量,用于快速再合成ATP
这一反应由肌细胞浆内活性很高的肌酸激酶催化完成,因此可以在运动中快速供能,是肌肉大力量高强度运动的主要能量来源。

2.肌肉细胞内的能量储备物质
①短时间大强度运动中主要的供能物质。在肌肉最大强度运动中,磷酸肌酸供能可以维持5~7 s,特点是快速将能量转移生成ATP,使肌肉输出功率大,但维持时间短。因此,磷酸原供能系统与速度、力量素质关系密切,影响短时间极量运动的运动能力。例如举重中拉起和举起杠铃,拳击中的快速出拳,短跑起跑后的快速加速跑等。
②运动训练影响肌肉内磷酸肌酸的储备量。CP是肌肉收缩的主要能量来源之一,肌肉内CP含量下降与运动强度成正比,在极量强度运动至力竭时,CP储备量几乎耗尽,为安静状态下3%以下。例如短跑的100 m跑和更短距离的连续最大速度冲刺跑。运动后肌肉内的CP含量可以迅速恢复,约30 s可以恢复至运动前含量的一半,2 min左右可以恢复至运动前含量水平。
③口服肌酸结合训练,可以增加肌肉细胞内磷酸肌酸和肌酸的储备量。一周左右口服肌酸并与训练结合,在最大强度的训练中CP大量消耗,在运动后的恢复中,CP含量可以增加20%~30%,同时肌肉内肌酸的含量也可以增加15%~25%。通过这种方式在肌肉中增加的肌酸和磷酸肌酸含量,可以维持约3周。
3.肌酸和磷酸肌酸在细胞内起能量转运作用
存在于肌肉细胞浆内的ATP和CP可以快速为肌纤维收缩和各种耗能的生命活动提供能量,但是人体最大量的能量来源于线粒体内糖、脂肪酸和氨基酸氧化分解生成的ATP,线粒体是细胞内的能量“工厂”,线粒体内生成的ATP不能直接透出线粒体,需要通过肌酸转换为磷酸肌酸转运出来,再合成ATP。因此,在运动中,线粒体内生成的ATP用于肌肉收缩,需要通过肌酸和磷酸肌酸的中转转换。
知识拓展
口服肌酸对人体运动能力的影响
一、口服肌酸对运动性疲劳的影响
由于口服肌酸能提高体内肌酸和磷酸肌酸水平,使运动时能有更多的能量合成ATP,延缓了运动性疲劳的发生。补充肌酸能从以下几个方面延缓肌肉疲劳:
①增加合成CP的能力,肌细胞内高浓度的CP有助于肌肉收缩持续进行。
②增加CP的再合成,恢复过程中肌细胞内高水平游离肌酸,使更多的CP得以再合成。
③减少肌肉内的酸度。磷酸肌酸在肌肉中充当代谢缓冲剂,由ADP合成ATP时要消耗H+,故能够提高肌肉的缓冲水平,增加大强度练习的运动时间。
④增加恢复期有氧代谢的能力。在锻炼过程中,肌酸能增加肌肉内蛋白质合成能力,耐力运动员在训练期间服用适量的肌酸,会更有利于身体机能的恢复。
⑤提高训练水平,增加肌酸或磷酸肌酸能使运动员适应更大的运动负荷,提高间歇性冲刺能力,这对于要求较高绝对力量的运动项目如摔跤、拳击等以及反复最大速度的运动项目,如手球、足球、橄榄球等是有利的。
二、口服肌酸的副作用和减轻口服肌酸副作用的几种方法
合理的口服肌酸可以明显提高骨骼肌的自由肌酸的浓度,从而增加CP的浓度,可提高肌肉短时间激烈运动的力量或速度。但如果口服不合理,则会发生下列的副作用:
①口服肌酸可以抑制体内肌酸的合成。因此,口服肌酸要注意:a.服用的量和时间,不能整个训练期中都用;b.服用肌酸的计划要和力量或速度等训练内容及增加有关营养相配合。
②增加体重。当每天口服肌酸20 g,5 d后可使体重增加1.3 kg。这对于控体重或降体重的运动员应注意。
③肌肉酸胀感。肌肉吸收肌酸的同时还要吸收水分,口服肌酸者前5 d,尿量在前3 d左右明显减少,这是水在骨骼肌中潴留的结果。这将使肌肉体积增大,而造成酸胀感,也使体重增加。减轻上述谈及的副作用,目前认为可采用的方法有:
a.与含糖饮料同时服用:饮用含糖饮料后可以刺激胰岛素的分泌,增加血胰岛素浓度,胰岛素可以促进肌肉从血液中吸收肌酸。
b.增加维生素E:维生素E缺乏时,骨骼肌中吸收肌酸减少,在口服肌酸时应增加维生素E。
c.增加优质蛋白质和氨基酸等。在口服肌酸期间要注意增加蛋白质的补充,如补充卵清蛋白或氨基酸等。因肌酸在肌肉细胞内增加时,会加速胞浆和线粒体间肌酸和磷酸之间能量穿梭转移,有助于肌肉蛋白质合成,使口服肌酸加快瘦体重的增加,同时有助于力量和速度运动能力的提高。
小结
磷酸原系统供能特点:快速供能,使肌肉达最大输出功率,但由于储存量较少,连续供能持续时间较短(4~5 s),补充肌酸可以增加磷酸原供能系统的能量储备,有利于力量和速度项目运动能力的提高。
探索与思考
1.磷酸原供能系统在所有强度的肌肉运动中均起重要的提供能量的作用,为什么?
2.通过进一步查阅资料,请您设计一个训练中补充肌酸的方案。如果不需要补充肌酸,为什么?
一、运动中肌肉内糖无氧代谢的基本过程
葡萄糖和糖原在缺氧的条件下,分解生成乳酸,同时生成ATP的生化过程,称为糖无氧分解代谢,又称为糖酵解。
(一)糖酵解的途径
如图3.5所示,骨骼肌细胞浆内,存在大量的肌糖原,在进行糖酵解时,首先分解为葡萄糖;血液中的葡萄糖进入骨骼肌细胞后,也可以通过糖酵解途径被分解产生能量,同时生成乳酸。
图3.5 糖酵解过程示意简图
(二)乳酸的代谢
骨骼肌内通过糖酵解生成的ATP可以被利用为肌纤维收缩等活动,而大量生成的乳酸也需要排出细胞,因为乳酸在肌细胞内堆积会严重抑制糖酵解的进行,称为终产物浓度抑制。
1.运动中乳酸的生成
在20~60~90 s短时间最大强度运动中,大量募集肌纤维收缩,糖酵解速率达到最大,乳酸迅速大量生成,直到运动结束,如200 m跑和400 m跑;在较长时间的大强度运动中,加速阶段和冲刺阶段也会大量募集快肌纤维收缩,生成大量乳酸,跑后血乳酸可达15 mmol/L左右,又如5 000 m跑,有氧代谢不能满足肌肉对能量消耗的需求,特别在比赛时,最后冲刺也会使糖酵解供能增加,也生成大量乳酸,如800 m、1 500 m跑也一样。运动中乳酸生成越多,说明通过糖酵解供能越多。
2.乳酸的消除
首先,从发生糖酵解的骨骼肌细胞转运出细胞。然后,从骨骼肌细胞转运出来的乳酸进入血液循环。最后,在运动中和运动结束后:①运动中通过血液循环进入心肌、骨骼肌内被完全分解为二氧化碳和水,是乳酸消除的主要方式。因此,大强度运动后及时进行低强度的恢复性运动,既增加肌肉内的血液循环,也是快速减少体内乳酸的最好方式,如快步频慢跑、自行车小传动比慢骑、放松性游泳等。②运动后在肝脏中重新合成糖原,占总量约20%以内。③转化为其他物质分子,占10%~15%。④通过汗液和尿液排出,约占5%。
3.乳酸消除的意义
①维持糖酵解系统持续供应能量。
②防止骨骼肌细胞内因p H值过度降低而降低ATP的利用效率。
③转移乳酸使其再利用。转移至心肌和其他骨骼肌纤维中分解提供能量。
多数的运动项目中,长期高水平训练的运动员,消除运动中产生的乳酸的能力很强,特别是心肌能更多地完全分解乳酸生成能量。
二、运动中肌肉内糖无氧代谢的调节
运动中的无氧代谢的调节包括3个方面:①能源物质的储量,即肌糖原、肝糖原的储备量,特别是肌糖原的储备量。②运动后肌糖原恢复的调节。③肌糖原糖酵解过程的调节,包括催化糖原分解的酶活性在训练影响下的改变,神经、激素等对糖酵解的调节,内环境变化时酸碱平衡的协调等。这3个方面的调节因素,受运动强度和运动时间的综合影响。我们重点了解糖原储备和恢复的调节。
(一)肌糖原、肝糖原储备量的调节
1.饮食
膳食中长期糖类(营养学中称为碳水化合物)(主食)摄入不足,肌糖原和肝糖原含量较低,运动中维持糖酵解供能的时间较短,肌肉力量不能维持易疲劳,不能维持大运动量训练。如体操等项目运动员减体重、健美运动员控制主食摄入等,容易发生糖原储备较少。
2.训练中糖原储备大量消耗
如图3.6所示,大强度长时间训练后,肌糖原和肝糖原均被大量消耗,训练后糖原储备处于很低的水平;在连续的大运动量训练中,训练前一餐碳水化合物摄入不足,会发生低血糖,说明训练前糖元储备量不足。

图3.6 不同运动强度和运动时间对肌糖原影响示意图
3.超代偿性恢复
训练结合碳水化合物摄入,可以使糖原储备量发生超代偿性恢复。大运动量的糖酵解供能训练和大量消耗糖原的长时间耐力训练,训练中肌糖原和肝糖原被大量消耗,训练中和训练后及时补充糖类,可以使糖原储存量超出运动前,称为糖原的训练后超代偿性恢复。
采用高糖膳食并在训练后及时补充糖类饮料的方法(糖原填充法),训练中糖原消耗越多,超代偿性恢复越明显,最多可以在48 h后超出运动前2~4倍。两周左右的糖原填充法训练,可以明显提高糖酵解供能能力和肌糖原、肝糖原储备量。
(二)运动后肌糖原恢复的调节
①运动后的高糖膳食(糖类占总能量摄入70%以上)可以提高运动后肌糖原含量。
②运动后及时补充糖类更明显增加肌糖原超代偿性恢复(图3.7)。运动后即刻至运动后2 h,骨骼肌细胞对胰岛素敏感性最高,同时糖原合成酶的活性也最高,及时补充糖类,肌细胞在胰岛素帮助下从血液中快速吸收更多的葡萄糖,用于更多地合成肌糖原。
③运动后及时补充糖类和优质蛋白质混合物,肌糖原超代偿性恢复优于单独补充糖类(图3.8)。

图3.7 不同时间的糖补充对肌糖原含量的影响
(引自冯炜权等,《运动生物化学研究进展》,2006)
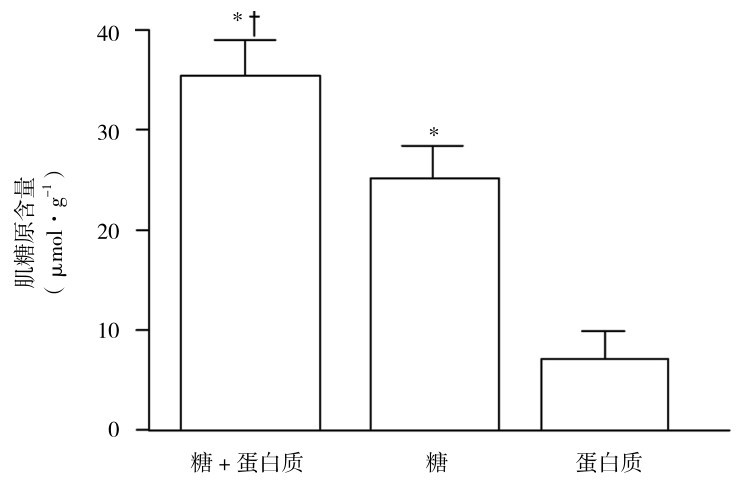
图3.8 运动后糖补充对肌糖原合成的促进作用及蛋白质添加的增强作用
(引自冯炜权等,《运动生物化学研究进展》,2006)
知识拓展
肌糖原糖酵解过程的调节
一、糖酵解酶活性的调节
1.糖原磷酸化酶活性调节
糖原磷酸化酶所催化的反应式将糖原分解为葡萄糖,是糖原分解代谢的限速步骤。磷酸化酶有a型和b型两种存在形式。安静时,此酶大多以低活性的b型存在,a型的比例是10%或更低;运动开始后, b型磷酸化酶迅速大量转化为高活性的a型磷酸化酶,使糖酵解过程迅速进行,生成能量供应肌肉收缩的需要。糖原磷酸化酶活性还受到肌细胞p H值的影响,p H值降低抑制了磷酸化酶b激酶,导致b型向a型的转化减少,糖原磷酸化酶a在高强度重复训练肌肉p H值达到6.4时被抑制,是肌肉中酸性物质的增加所致,结果是抑制了糖酵解过程,减少能量生成,降低肌肉力量。
2.二磷酸果糖激酶活性调节
通过二磷酸果糖激酶的催化,一个葡萄糖分子分解为两部分,是糖酵解生化反应过程中的限速酶。当肌细胞内ATP浓度降低时,如ATP-CP大量消耗后,二磷酸果糖激酶活性提高,增加糖酵解过程的反应速度,加快ATP的生成。
二、神经调节
大强度运动中,通过神经调节动员大量肌纤维特别是快肌纤维参与肌肉收缩,进而启动肌细胞内的糖酵解供能系统,分解肌糖原产生能量。
三、激素调节
大强度运动中,血液肾上腺素浓度迅速升高,直接刺激糖原磷酸化酶活性增加,加快糖酵解系统供能。
探索与思考
血乳酸测试,反映运动中糖酵解供能。运动后血液乳酸浓度越高,说明运动中糖酵解供能程度越高。如图3.9所示说明连续大强度运动中,休息间歇越短,CP不能及时恢复,动用糖酵解供能越多。如图3.10所示说明随着运动强度逐渐提高,到达运动强度的某一点时,开始大量动用糖酵解系统供能,血乳酸浓度开始急剧增加,这一点称为“乳酸阈”运动强度。

图3.9 10 s跑间歇训练休息时间与血乳酸浓度及跑次的关系

图3.10 递增负荷跑台跑血乳酸浓度变化曲线
小结
糖酵解供能系统的特点是供能速度较快,供能速率是磷酸原系统的约1/2,但却是有氧供能系统的约两倍。糖酵解供能系统是在高强度运动中,骨骼肌内CP消耗后主要的供能途径,磷酸原供能系统和糖酵解供能系统合称为无氧供能系统;在亚极量较长时间(1.5~5 min,CP大量消耗后)大强度运动中,利用氧分解葡萄糖和脂肪等生成ATP的速率不能满足肌纤维收缩的需要,同时也通过糖酵解途径为肌肉收缩供能,称为无氧-有氧混合供能。
糖酵解供能系统主要受运动强度和运动时间的影响,通过神经、激素、糖酵解催化酶活性调节糖酵解供能系统的供能。膳食糖类摄入和运动后糖类补充显著影响肌糖原和肝糖原含量,合理膳食和运动后及时补充糖类可以显著提高肌糖原含量,从而增加糖酵解系统供能能力。
一、运动中肌肉内糖有氧分解代谢的基本过程
肌肉内的肌糖原和肌肉从血液中摄取的葡萄糖,在利用氧的条件下,完全分解成二氧化碳和水,同时生成ATP的生化反应过程,称为糖的有氧分解代谢。
在肌细胞内的糖有氧分解代谢反应,是分为两部分的连续分解过程,分别在细胞浆和线粒体内进行,在人体其他组织细胞内也进行同样的过程,如图3.11所示。终产物为二氧化碳和水,同时生成ATP。生成的ATP通过肌酸-磷酸肌酸转运出线粒体才能被肌纤维收缩利用,ATP的生成速率只有糖酵解的约1/2,所以肌肉利用糖有氧氧化供能产生的力量小于利用糖酵解和磷酸原系统供能的力量。

图3.11 糖有氧氧化途径示意图
二、运动中肌肉内糖有氧代谢的调节
运动中的糖有氧代谢的调节同样包括3个方面,与糖的无氧代谢的调节相似。
1.肌糖原、肝糖原储备量的调节
长时间大强度有氧运动大量消耗肌糖原和肝糖原,因此其储备量的调节与糖无氧代谢相似。
2.运动强度对糖有氧分解过程的调节
①运动强度为无氧阈强度时(血乳酸浓度约为4 mmol/L),糖有氧分解代谢速率达到最高值。进一步增加运动强度,则糖酵解代谢增强,并且对糖有氧分解代谢产生抑制。
②长期通过糖有氧代谢供能的运动训练,可以提高所有催化糖有氧分解酶的活性,使糖有氧分解代谢产生ATP的速率增加,从而提高糖有氧分解代谢供能中的肌肉力量和输出功率。
3.运动后肌糖原恢复的调解
与糖无氧代谢调节原理相似。
4.运动强度与糖有氧代谢供能的关系
肌细胞内的肌糖原,以及肝脏内的肝糖原分解为葡萄糖进入血液循环后到达肌肉可以被肌肉摄取,均可以在运动中通过有氧分解代谢提供能量,在运动中,糖有氧代谢供能与运动强度关系密切,表3.1可以看出不同运动强度的项目运动中各种供能系统的供能比例,如图3.12和图3.13所示同样可以看出不同时间的最大强度运动中,肌肉内不同的供能系统随运动强度不同而起不同程度的作用。中等距离、2~10 min内的大强度运动中(65%~85%VO2max强度),糖有氧代谢供能占总供能比例较高。
表3.1 各种不同强度运动中不同的能量供应系统所占的供能比例
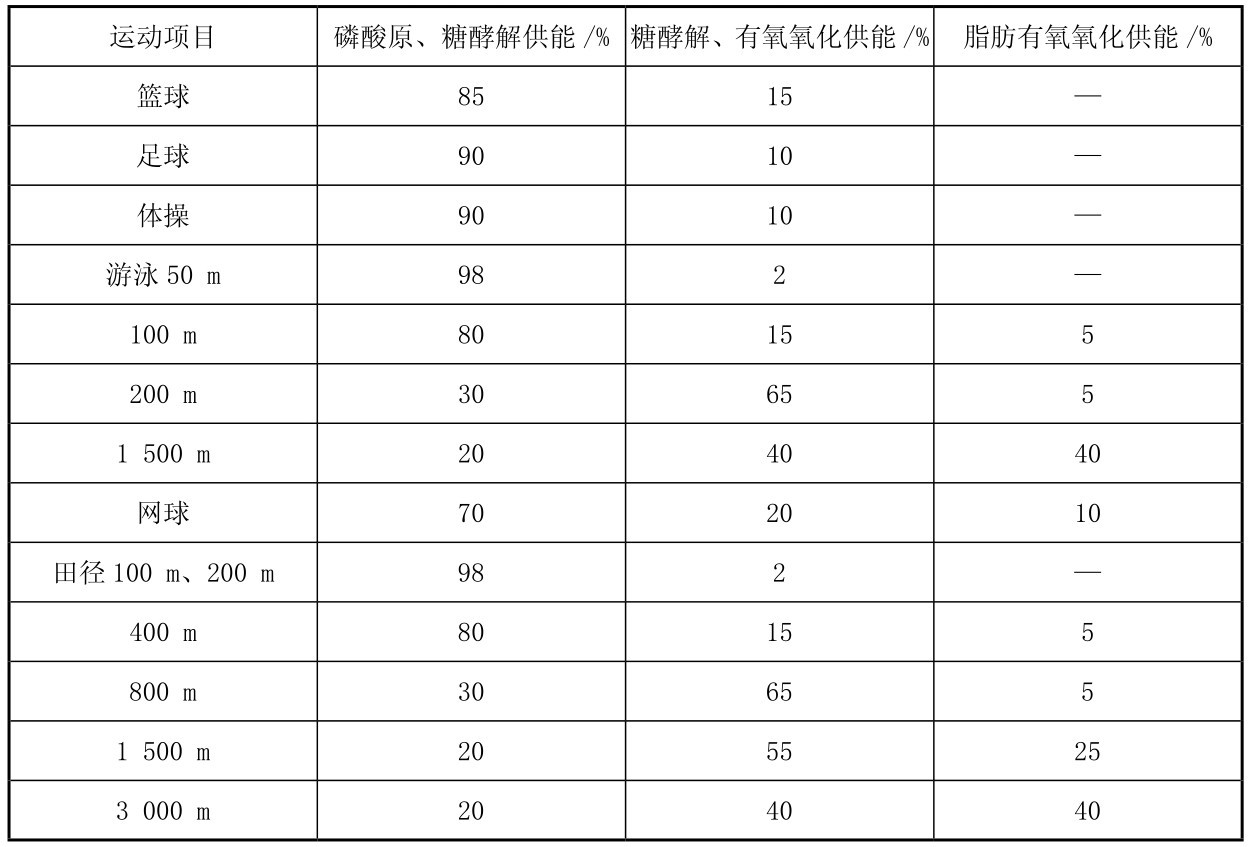
续表
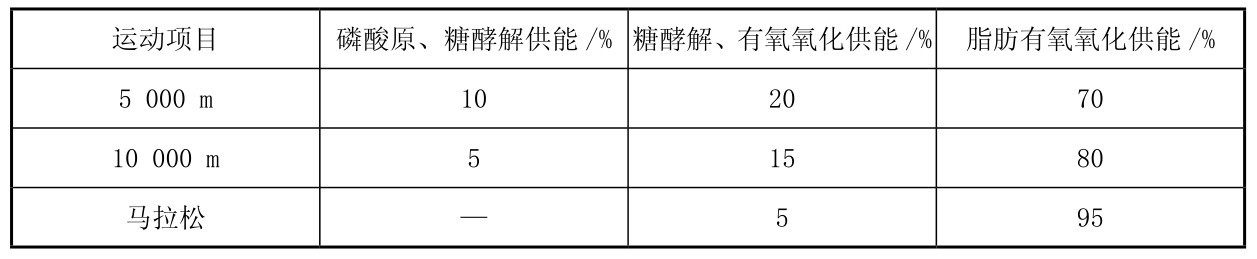

图3.12 运动时3个供能物质代谢系统在总能量中的比例示意图
(引自冯炜权等,《运动生物化学进展》,2006)
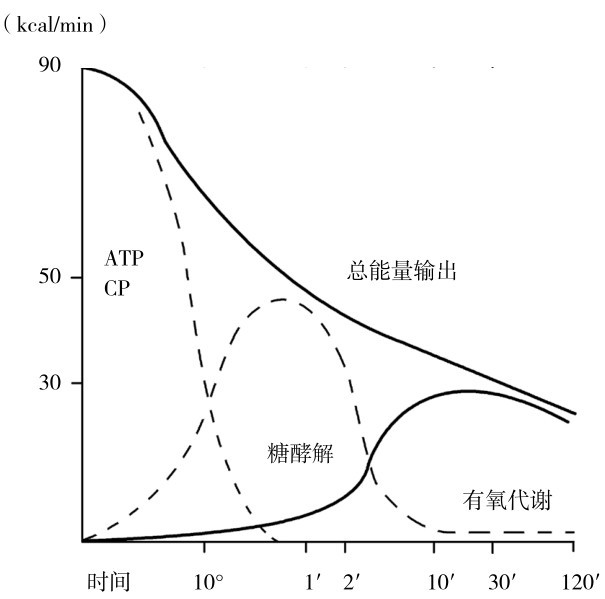
图3.13 人体骨骼肌运动时的供能生化过程和功率输出的相互关系
(引自冯炜权等,《运动生物化学进展》,2006)
知识拓展
糖的供能特点
人体在运动时,糖是重要的能源物质,因为它有以下供能特点:1.在不同氧供应状况下,糖都可以充分发挥其供能作用
像在100 m、400 m和800 m跑等运动项目中,氧供应不足,糖可以通过无氧酵解生成乳酸提供能量,以维持运动能力;在氧供应充足的耐力运动项目中,如马拉松跑、50 km竞走等,糖可以通过有氧代谢分解成二氧化碳和水,并提供能量以生成ATP维持运动能力。比较脂肪和蛋白质只有在有氧状况下才可以分解供能的方式,糖的这种分解供能无疑具有明显的优越性。而且在同样的供氧条件下,糖的能量生成效率是最高的,每消耗1 m L氧,糖可以释放能量生成6.2~6.5 mol的ATP,而脂肪只能生成5.6 mol的ATP。
2.糖的利用速率快
在一般情况下,任何运动都很少只动用一种代谢途径去补偿运动中消耗的ATP,因而运动时骨骼肌ATP的含量变化很少,但它的总利用量很大,在激烈的耐力运动中为每分钟1~2 mol,可见ATP的转化速率相当迅速。在大于2 min的大强度有氧耐力运动中,有氧代谢虽然不是唯一的,但被认为是具主导作用的ATP产生途径。而此时,有氧代谢的底物主要是糖类。运动开始时先利用糖原,如100 m跑、50 m游泳这样的短时间运动,在运动开始3~5 s糖就通过酵解参与供能,短跑时糖酵解供能速率比步行时可增加约120倍,由0.3 mmol/(kg·min)-1提高到40 mmol/(kg·min)-1;持续运动5~10 min后,血糖开始参与供能,最多可达到安静时速率的20倍,当运动强度达最大摄氧量强度时,可达安静时的50倍;随着血糖被骨骼肌、大脑等组织的利用,肝糖原分解速率可比安静时增加5倍。这说明,运动时糖供能速率快,其储量是影响运动能力的重要因素。
3.糖是运动时消耗最多的供能物质
Wilmore和Costill等(2004)计算的各项运动的能量消耗物质供能比例显示,糖对大强度运动项目极其重要,在这些项目中的供能比例往往在90%以上。表3.2是几项运动中糖的供能情况。从图3.1可以看出,不同强度运动中肌糖原、血糖、血浆游离脂肪酸及肌甘油三酯供能的状况,不难看出,随着运动强度的增加,糖类供能的比例也大幅度增加。当然,就脂肪酸的供能绝对值来讲,也有较大幅度的升高,尽管相对值减小。因此减肥或控体重者在有限的时间内要想消耗掉等重量的脂肪,65%VO2max的强度显然比25%VO2max要好一些。
表3.2 跑、游泳、自行车运动的能量消耗和糖供能

注:按70 kg体重计算。
(引自Willmore and Costill,2004)
探索与思考
1.总结:运动中糖作为供能物质,通过糖酵解供能系统和糖有氧氧化供能系统生成能量,如何利用运动中糖的供能特性,提高相关项目运动中糖的供能能力?
2.如何通过血乳酸测试,分析运动中糖的供能能力?
小结
通过糖的有氧分解代谢生成ATP的效率很高,但是ATP的生成速率只有糖酵解的约1/2,并且需要肌酸-磷酸肌酸转运出线粒体才能被肌纤维收缩利用。
在中等距离、2~10 min内的大强度运动中(65%~85%VO2max强度),糖有氧代谢供能占总供能比例较高,且在此运动强度范围内,随运动强度提高,糖有氧代谢供能逐渐加强。
一、运动中肌肉内脂肪有氧代谢的基本过程
在肌肉内,通过利用和消耗氧,分解脂肪和脂肪酸产生能量的生化反应过程,称为脂肪有氧氧化代谢系统。
在人体组织中,利用脂肪有氧氧化分解代谢产生能量的组织主要是心肌和骨骼肌的慢肌。脂肪有氧氧化分解代谢可以分为以下5个步骤:
1.脂肪动员
运动时交感神经兴奋,血液肾上腺素增多,激活脂肪细胞内的脂肪分解酶。储存在皮下或腹腔脂肪组织中的脂肪,在脂肪分解酶的催化下,分解为脂肪酸和甘油并释放进入血液,这一过程,称为脂肪动员。
2.血液运输
通过血液循环的转运,血液中的脂肪酸到达肌肉后被吸收进入肌细胞内。
3.L-肉碱转运,在肌细胞内转移至线粒体
如图3.14所示,进入肌细胞内的脂肪酸先转化为脂酰辅酶A,然后再转移至线粒体膜与L-肉碱结合,生成脂酰肉碱,然后再转运至线粒体膜内侧,再转化为脂酰辅酶A,因此L-肉碱为脂肪酸进入线粒体的转运载体,这一转运过程是肌细胞内脂肪酸氧化的限速步骤。中、短碳链脂肪酸可以不需要L-肉碱的转运直接进入线粒体,但是长碳链脂肪酸必须要L-肉碱的转运才能进入线粒体,因而中、短碳链脂肪酸的氧化利用速度比长链脂肪酸要快。
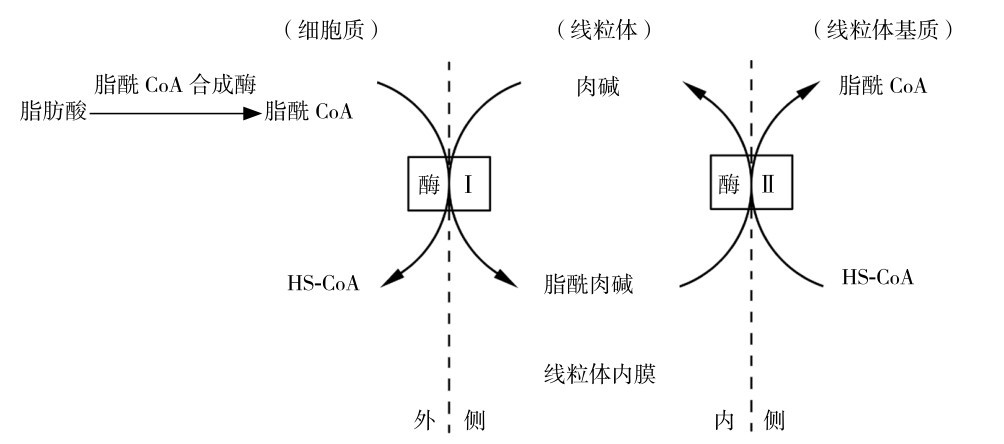
图3.14 脂肪酸在肌细胞内转移至线粒体
4.在线粒体内进一步分解
具体过程可以不了解。
5.在线粒体内氧化分解为二氧化碳和水,同时释放能量生成ATP
二、运动中肌肉内脂肪有氧代谢的调节
(一)运动中脂肪供能的特点
①储备量大,正常生理情况下,远远超出使用量。
②是长时间、中低强度运动时的主要供能的能源物质。由于脂肪动员、转运和脂肪酸的分解步骤复杂,需时较多,输出功率又不高,因此不是短时间、极限强度运动的能源物质,而只能作为长时间、中低强度运动时的主要能源物质(图3.15、图3.16)。
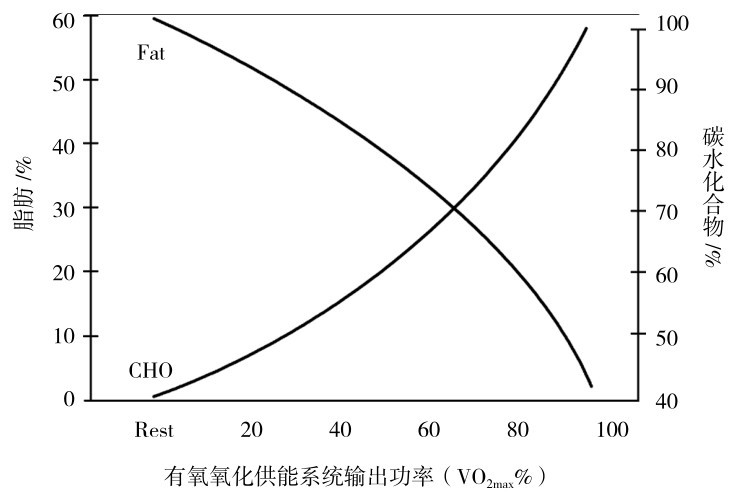
图3.15 运动中糖与脂肪作为能源的关系
引自Don Maclaren,2012)
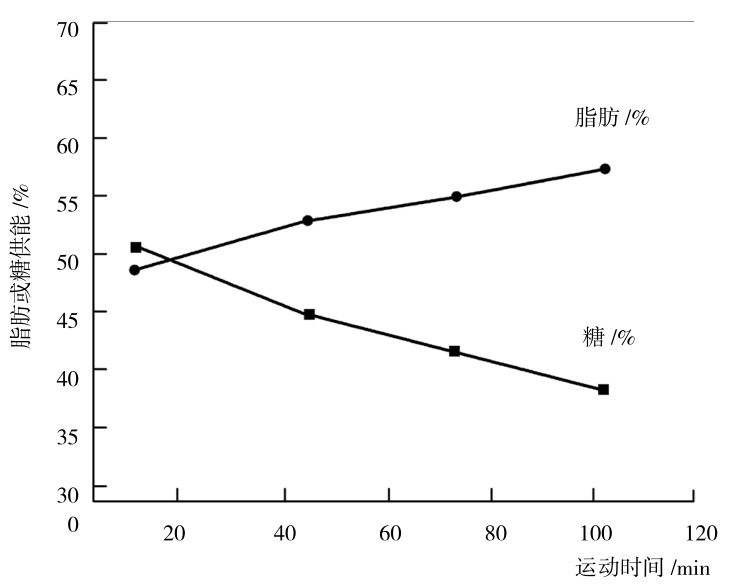
图3.16 在稳定强度运动中,糖与脂肪供能在总能量输出中的相对贡献
(引自Don Maclaren,2012)
在运动强度低于70%最大摄氧量、持续运动时间分别为40,90,180,240 min时,脂肪供能占总耗能的比例分别为37%,37%,50%,62%。但应该指出的是,只要在一定强度范围内,由于大强度运动时总能耗加大,因此,在相同运动时间的条件下,消耗脂肪的总量并不少。如分别以75%和25%最大摄氧量运动,脂肪的消耗量分别是110 cal/(min·kg)和70 cal/(min·kg)。可以看出,在两种运动时间相同的情况下,强度较大的运动,脂肪的总消耗量也大。
(二)影响运动时脂肪供能的因素
1.脂肪动员速度的激素调节
运动时交感肾上腺素能系统的活性提高,当运动强度在50%~70%最大摄氧量时,交感肾上腺素能系统的兴奋性就明显提高,血浆肾上腺素和去甲肾上腺素浓度也明显增高,提高脂肪酶的活性,加快脂肪动员。
2.乳酸对脂肪动员速度的调节
乳酸是糖酵解的代谢产物,由于糖酵解在运动中有重要的供能意义,但当时间超过30 s~1 min的最大强度运动,运动骨骼肌细胞中的乳酸产量都会增加。如果乳酸产生的速度超过它的消除速度,就会出现乳酸在骨骼肌细胞中堆积的现象。乳酸可阻断或抑制肾上腺素加速脂肪动员的作用。
耐力运动训练可提高骨骼肌细胞有氧氧化的能力,如增加骨骼肌细胞内线粒体的体积和数量,提高有氧代谢酶的活性等,减少相同运动强度下乳酸的产生数量,削弱乳酸对脂肪动员的阻断或抑制作用。同时,训练还可提高机体对乳酸的耐受能力,即使在乳酸增加的情况下,也能保持较高的脂肪动员速度。因此,长期的有氧运动训练可提高人体脂肪动员的能力。
3.运动中可利用糖量对脂肪利用的影响
运动中糖和脂类往往是骨骼肌细胞的主要供能物质。由于糖供能的输出功率较大,启动速度又较快,因此,在糖储量丰富时,骨骼肌细胞会更多地利用糖来供能。但由于体内糖的储备量相对较少,如果完全依赖糖代谢供能,则运动持续时间很有限。为了获得最大耐力,糖和脂肪必须合理利用。骨骼肌内可利用糖的量较少时,就尽可能多地利用脂肪酸供能,可以节约利用糖,使有限的糖储备维持较长的运动时间,直到运动结束,有利于维持长时间运动中的速度耐力。L-肉碱的数量是长链脂肪酸氧化有限速作用,因此,补充肉碱可能在一定程度上达到多利用脂肪供能,节约肌糖原的目的。
4.肌细胞糖储备对脂肪利用的影响
糖和脂肪都是人体的重要能源物质,糖储量的变化会影响脂肪的分解数量。高糖膳食使肌糖元储备增加,运动中能更多地利用糖供能,而脂肪氧化量较少。说明如果运动前提高骨骼肌细胞内糖的储备,可以减少运动中对脂肪的分解利用,同时也有利于维持较高的肌肉力量。
5.耐力训练水平对脂肪利用的影响
耐力训练能提高运动员利用脂肪供能:①耐力训练可以提高脂肪动员能力,加快运动中脂肪的动员;②耐力训练使骨骼肌中线粒体的数量和有氧氧化酶的活性均显著提高,能够加快脂肪供能。如图3.17所示,通过训练,在运动中可以很明显地提高肌肉利用脂肪酸的能力。
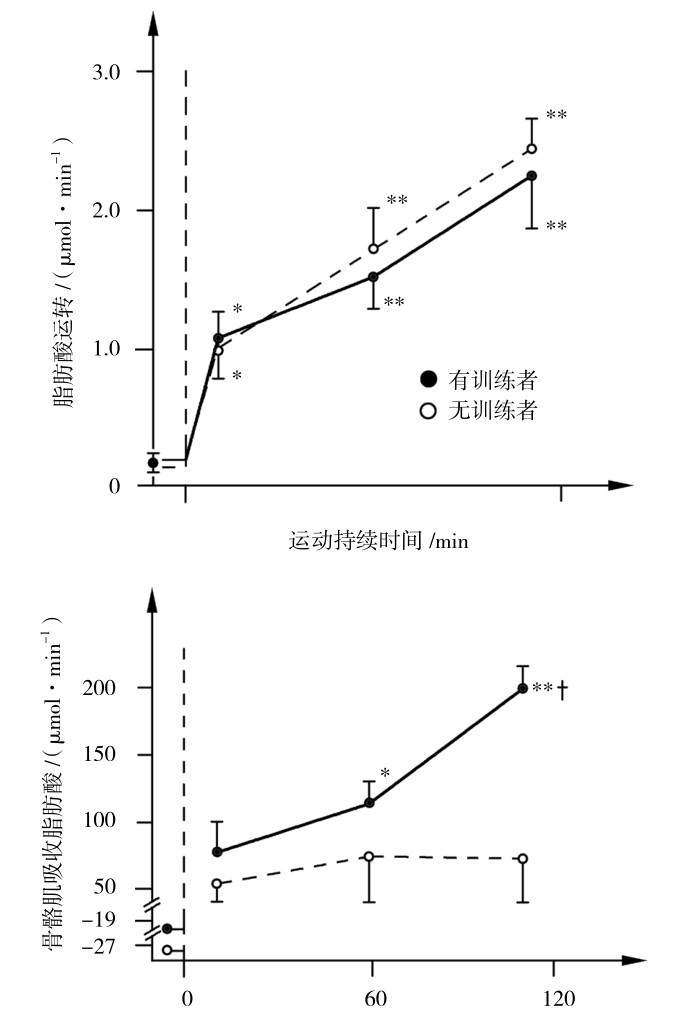
图3.17 运动中骨骼肌对脂肪酸的利用
(引自Don Maclaren,2012)
知识拓展
运动与脂肪代谢平衡
运动中可利用来分解供能的脂肪主要来源于两个途径:①骨骼肌细胞内储备的脂肪酸;②从脂肪组织中经过脂肪动员出来的血浆游离脂肪酸。
运动强度在25%最大摄氧量时,血浆游离脂肪酸浓度增加的速度是安静时的5倍;运动强度达25%~65%最大摄氧量时,肌糖原分解成了重要的能量来源,血浆游离脂肪酸浓度增加的速度不变,脂肪分解的总量上升,其来源应该是骨骼肌细胞中储备的脂肪酸;运动强度达85%最大摄氧量时,骨骼肌收缩主要靠肌糖原分解成供能,这时,血浆游离脂肪酸浓度下降。因此,如果只强调运动中的能量消耗,对减体重、降体脂而言,低强度运动无疑较高强度运动效果更好。但这又无法获得大强度运动对提高运动后能量消耗的益处,如何将两者结合起来,是很值得研究的。
长期的耐力训练,可以改变食物脂肪在体内的流向。经过长期耐力训练的人进食脂肪后,消化吸收的脂肪酸有更多的储备在骨骼肌细胞中,以便运动时分解供能。而其吸收的食物脂肪酸则较少进入脂肪组织,因此,长期进行耐力运动,对改善全身脂代谢和血脂成分调整体脂分布和体成分,都是非常有益的。
探索与思考
查阅资料,讨论补充L-肉碱并结合运动能否增加身体脂肪的氧化分解、减少体脂含量?为什么?
前沿话题
不同的供能代谢系统在运动训练中是相互联系的
需要说明的是,运动中通过大脑运动皮层甚至心理意志的作用,调节、维持或加强运动神经元的活动,运动神经元则决定了动员募集的肌纤维数量和肌纤维收缩频率的动态变化,从而调节、维持或加强肌肉的力量。这一复杂的神经过程,对运动中能量消耗的速率、总能量消耗起决定作用;相应地,运动中人体的呼吸和氧供应、肾上腺素和皮质激素等运动中促进能量生成的激素与神经作用协调,调节运动中能量的生成方式和供能速率,整合能量代谢系统,使不同的能量代谢系统相互促进,而非单独起作用,如肌酸-磷酸肌酸系统促进糖酵解系统的供能,适量的糖酵解系统的产物可以激活有氧氧化系统,肌酸-磷酸肌酸系统通过能量转运促进有氧氧化系统的供能。另外,运动中肌肉内蛋白质分解增多,包括收缩蛋白质成分和其他功能性蛋白质成分,虽然其中某些氨基酸如枝链氨基酸可以通过三羧酸循环分解产生能量,但是在正常情况下其提供的能量只是总能量消耗的极少部分,因此一般不考虑蛋白质在运动中的供能。
在运动后人体的恢复中,能源物质特别是磷酸肌酸和肌糖元恢复合成、蛋白质的恢复合成以及其他多种生理机能的恢复和细胞内环境平衡的恢复,均需要消耗能量,肌肉中这些能量一般大部分来自于脂肪酸的有氧氧化,其他器官中主要来自于葡萄糖(从血液中摄取)的有氧氧化,这是运动生理学中提到的“运动后过量氧耗”的主要原因。
胰岛素在运动后恢复中促进各个器官包括骨骼肌组织从血液中吸收各种营养物质(葡萄糖、氨基酸、脂肪酸、维生素等),用于能源物质和蛋白质的恢复合成,促进蛋白质合成激素主要是雄性激素(如睾酮)以及生长激素促进运动后蛋白质的恢复合成和超量代偿性恢复。这些是激素在运动后对物质合成代谢恢复的调节作用。
一、人体运动中的水盐代谢与体温调节
(一)运动中的体温调节
根据外界环境的温度,皮肤的温度可以在较大的范围内变动,但深部组织的温度则必须保持在正常体温37 ℃左右很小的范围内。因此,身体的产热率和散热率必须保持平衡,否则就会导致体温的变化。人体内发生的所有化学反应产生的化学能大部分表现为热能,因此,产热率和代谢率是成正比的。运动中代谢率的增加是和能量转化成比例的。优秀马拉松运动员能够用2 h多一点的时间完成马拉松比赛,这样的速度会导致机体的产热率达到1 200 W,但是其体温的上升却很少超过2~3 ℃,这表明机体散热率也随着产热率的上升而升高。一般来说,运动中体温的上升和运动强度是成比例关系的。
人体运动中体温的调节方式中,人体体表与环境之间的物理热交换主要以传导、对流、辐射3种方式进行,通过汗液蒸发散发身体热量是人体主动进行的散热方式。环境温度是人体散热最重要的影响因素。
1.传导
空气的导热性较差,水的导热性是很好的,因此,水温28 ℃时,人们感觉暖和,而水温低于28 ℃时,人们却感到凉,甚至冷。因此,水温对游泳运动员的成绩有相当的影响。
2.对流、辐射
当人体皮肤的温度远远高于环境温度时,对流、辐射是非常有效的散热方式,在这种条件下,即使在大强度运动中,这两种方式也是主要的散热方式。反之,随着皮肤与环境的温差减小,当环境温度高于35 ℃时,机体开始从环境中吸收热量。
3.蒸发
运动中体内大量快速产热,特别是环境温度也同时较高时,体内蒸发就成了散热的唯一方式。当机体代谢产热量很高,物理散热很有限,有可能导致体温上升时,快速蒸发散热是十分重要的。由体表每蒸发1 L汗液,可以散发身体热量2.6 MJ(620 kcal)。
以马拉松运动员为例,假设其产热率为1 200 W,蒸发散热的作用就非常明显。如果没有热交换机制,体温会快速上升并在运动开始后20 min内达到人体耐受的上限。1 L/h的汗液蒸发速度可以使散热量达到2.6 MJ/h(620 kcal/h),相当于722 J/s(172 cal/s)或722 W。因此,身体代谢产热的总量相当于1.7 L/h的汗液蒸发率所散失的热能。高蒸发率要求高的汗液分泌率,这就导致随着出汗的增多,水和电解质进行性丢失。
尽管人体通过皮肤的汗液蒸发散热能力很高,但人体必须不停地排汗,保持皮肤的湿润。当皮肤与环境之间的蒸气压差很低时,蒸发就会被阻碍。这种情况在环境水蒸气压很高时更明显。如衣服妨碍空气的流通,使皮肤周围的水蒸气达到饱和,从而限制汗液的蒸发。大的体表面积、高的风速都可以促进蒸发散热,但当环境温度高于皮温时,它们也会因为辐射、对流的提高使身体从环境中吸收的热量增多。
(二)运动中的水盐代谢与体温调节
1.运动时的脱水
运动时代谢率增加,除25%的代谢产能用于参与外部工作外,其余的都以热量的形式散失。在运动过程中,能量需求增加后,随之而来的是产热的增加。体重70 kg的人,正常安静的氧耗是千克体重每分钟4 m L,产热率为60~70 W(相当于一个小电灯泡的热量),维持2.5 h马拉松跑的需氧量约4 L/min,相应的产热率可达1 100 W,体温也开始急剧升高。
①散热和脱水方式。为了避免体温升高的有害影响,机体的散热率也需相应增加。运动员主要是通过排汗来调节体热平衡。运动时,皮肤温度保持在较高水平有利于体热的辐射和传导,但这些机制只有在周围温度低并且风速快时才发挥积极作用。当环境温度高于皮肤温度时(大约超过35 ℃),机体只能通过蒸发的形式散热。每蒸发1 L水就有2.4 MJ的体热散失。一个70 kg体重的人进行2.5 h的马拉松跑,如果仅以蒸发的形式维持产热与散热平衡,就需要汗液以1.6 L/h的速度蒸发。在如此高的出汗率情况下,有部分汗液未经蒸发即从皮肤流失,汗液分泌量达到2 L/h才可能满足蒸发散热率的需要。这种情况可能发生,但会导致5 L的水分丢失,对于70 kg的人来说相当于7%的体重。此外,水分还可以通过呼吸道蒸发,尽管这不是散热的主要渠道,但在炎热、干燥的环境中进行激烈运动时,经呼吸散发的热量可占相当比重。
②项目差异。在不同项目运动中,能量需求是不断变化的,排汗量差异也较悬殊,主要取决于当时的气温、热辐射强度、气压、湿度、运动强度、饮食含盐量以及血浆中的含盐量等因素。表3.3列出了不同运动项目的平均出汗量,但平均值往往掩盖了两极反应。即使在很低的环境温度下也可能观察到高出汗率的情况,比如像马拉松这种能量需求很高的项目,因此,不能认为脱水是仅仅存在于高温环境中。但是,汗液的丢失确实与环境状况息息相关,体液的大量流失在夏季和热带气候更常见。据报道,夏季参加比赛的马拉松运动员体液流失可达6 L甚至更多,相当于8%体重或12%~15%的总体液量。尽管个体差异明显,但对不同组的马拉松运动员的研究发现,跑速与出汗率有关,但与总出汗量无关。
表3.3 不同项目运动的出汗率
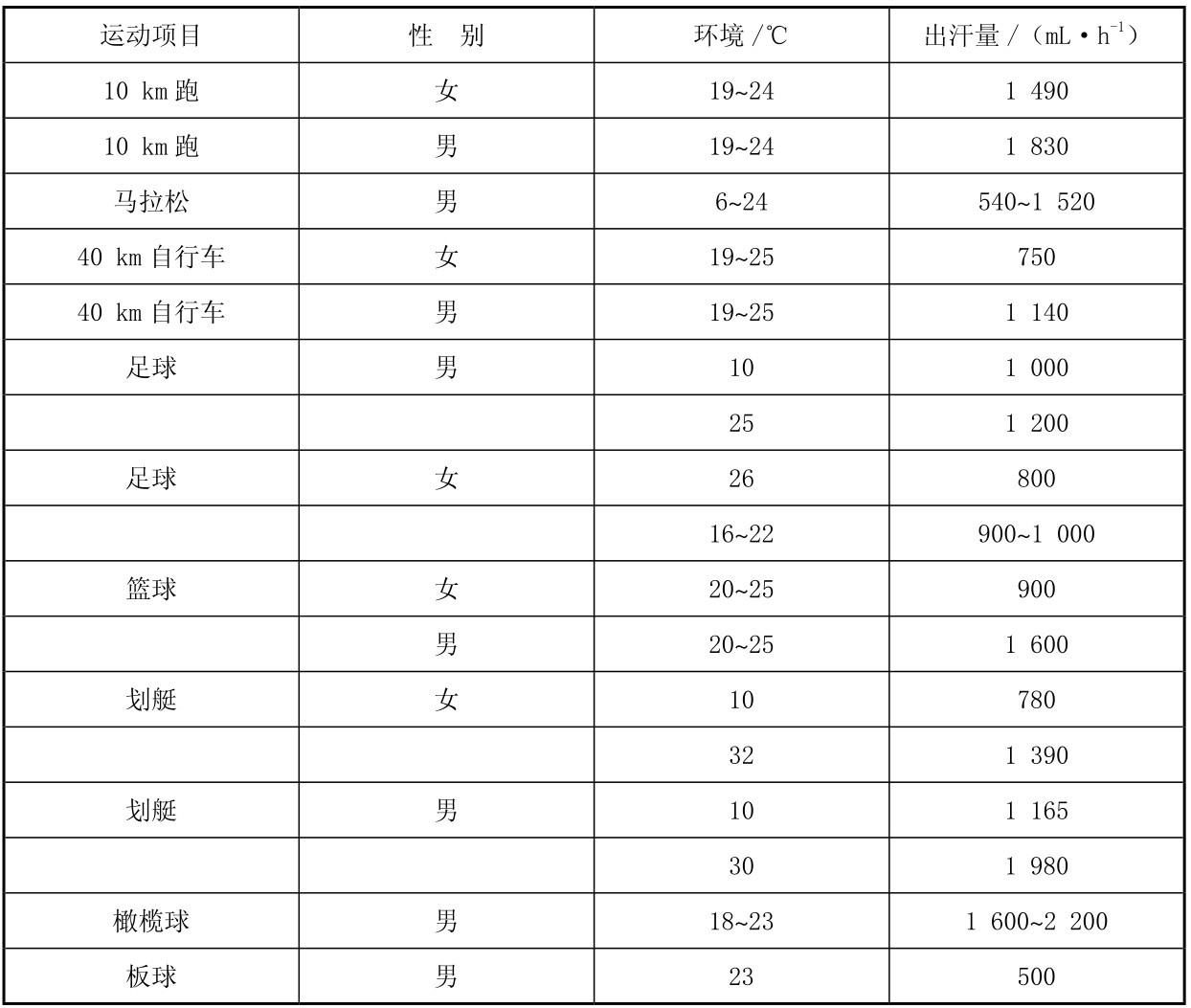
注:出汗量为均值,不能反映个体差异。 (引自Reher and Burke,1996)
③体重、温度影响。运动员在训练或比赛时的排汗量和运动负荷强度及温度有关,在热环境下运动会增加排汗量。表3.4显示,当运动员在不同气温下以不同速度跑时,随负荷强度增加和温度的上升,排汗量相应增加。
表3.4 不同体重运动员在不同温度环境下不同跑速时的出汗量

续表

(引自Nissen等,1992)
④性别、年龄差异。在一般情况下,女性比男性出汗少,即便在经过一段适应期后也是如此。但这种所谓的性别差异可能在很大程度上是由于训练和适应程度不同造成的。关于年龄对排汗能力影响的研究有限,健康及适应水平是很重要的影响因素,但一般而言,排汗反应是随年龄而下降的。这并不代表女性和老人无法适应热环境中的运动,也不意味着即使在热环境下他们也无须特别注意液体的补充。儿童与成人在运动排汗反应及汗液成分上有所不同。儿童的排汗能力及汗液中的电解质浓度(以体表面积计)均低于成人,但儿童补充液体和电解质的需要与成人同样重要。实际上,发生相同程度的脱水时,儿童的体温增加较成人明显,因此他们补液的需求可能更高些。
2.运动中的电解质丢失
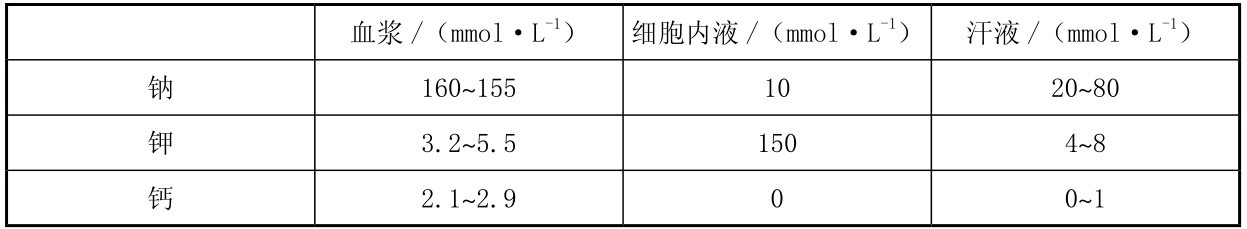
机体丢失的水分首先来自细胞外液,因此,汗液中的氯化钠浓度较高。汗液中电解质的流失实际上反映了发汗率和汗液成分的变化,而后两者除了随时间变化外,还受运动强度﹑环境及生理机能的影响。与电解质的易变相比,血浆的等渗环境相对稳定,其主要电解质是钠和氯(表3.5)。离子的浓度通常以mmol/L计算,而钠盐的丢失量却是以g为单位评定的。以每升汗液含钠50 mmol计算,那么,每失汗1 L,即失钠2.9 g。如果运动员在一天的训练中出汗达5 L,其失钠量即达到15 g。即使由于大量出汗而使汗液中钠的浓度及排尿量减少,盐的丢失量对一般的摄入量来说是太多了,而且在高温环境中钠盐的代谢更加变化不定。因此,运动中补液应考虑高温环境的影响。
机体经汗液可丢失大量的盐。当大量出汗时,电解质的丢失十分明显。汗液中各种电解质成分的丢失量受其浓度和总出汗量的影响。表3.5列出了血浆﹑细胞内液及汗液的电解质成分。
表3.5 细胞内、外液正常电解质浓度
续表
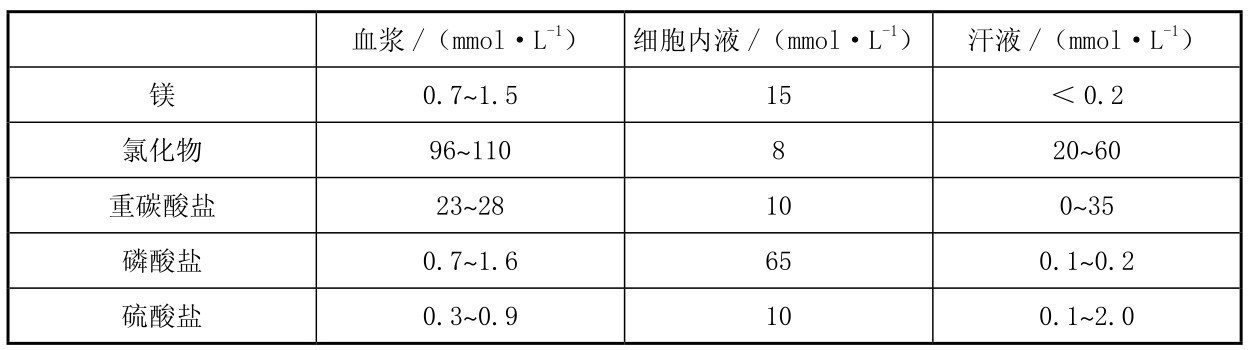
汗液的成分是存在个体差异的,但即使是同一个体也会因排泄率、训练程度及热适应性的不同而变化。随着训练水平的提高和热适应性的增强,热刺激时的排汗率增加而电解质丢失减少。机体在保存电解质的同时增加了热调节能力,称为热习服。汗液的主要成分与细胞外液相同,主要是钠和氯,当然其浓度低于血浆。经过适应性训练后,增加排汗率的同时可保证汗液中钠盐的浓度下降。相对而言,汗液中钾的浓度受出汗率的影响小,镁的浓度变化很少或轻微下降。
二、运动中水盐代谢调节与运动能力
(一)失水对运动能力的影响
失水对运动能力的影响程度与失水量的多少及训练水平有关。不论是耐力性有氧运动还是高强度无氧运动,脱水通常会损害运动能力。一般来说,脱水达体重的2%时,运动能力即受到轻微损害;当脱水超过体重的5%时,运动能力可下降30%。研究发现,长时间运动使机体丢失约2.5%体重的体液,进行大强度的能力下降了45%,而预防脱水发生可以增强运动能力。但对于训练水平较高的运动员,特别是经过多年训练对脱水有一定耐受的运动员来说,失水即使超过5%,摄氧量、心输出量和运动能力均无明显影响。
1.脱水影响机体的最大摄氧能力和运动能力
在温和的气候中,身体丢失水分少于体重3%的水分时,不会改变最大摄氧能力;身体丢失水分超过体重的3%时,最大摄氧能力下降。在炎热的气候中,小量的或中等的水量减少就能够导致最大摄氧能力的大幅下降,热环境加重缺水导致的最大摄氧能力下降。低水合时,递增强度运动时机体运动到疲劳时的能力降低。少量(体重的1%~2%)的脱水降低体能,但不降低最大摄氧能力。在炎热的气候里,低水合降低最大摄氧能力的幅度更大。
2.低水合降低运动员的耐力和力量
如缺水对长跑运动员耐力的影响。让运动员分别在正常水合和低水合时完成1 500,5 000, 10 000 m跑步。他们用速尿使运动员体重减少2%,血容量下降11%。各种距离的跑步项目,运动能力都下降了,只是长距离(5 000,10 000 m下降5%)比短距离(1 500 m下降3%)下降程度更大。脱水(相当体重的3%)对2 000 m划船的影响研究中,结果是低水合比正常水合时慢22 s,力量下降5%。
3.脱水使运动员更容易疲劳
通过自行车测力计来研究低水合对运动能力的负面影响。在两个实验中,志愿者分成补水和不补水两组,蹬车55~60 min后立即测量其运动能力。一个实验中以90%最大摄氧量强度蹬车时,补水组发生疲劳的时间比不补水组长51%(6.5∶9.8 min)。另一个试验中发现,补水组比不补水组蹬车速度快6.5%。
(二)运动中水、电解质补充
当因出汗导致水和电解质大量流失出现脱水时,会降低机体体温调节的能力,肌肉耐力和力量也会随之下降。冯炜权等总结(表3.6)认为,在以下情况中需特别注意补水,以预防疲劳的发生,防止运动能力下降。
表3.6 运动员需要补液的情况
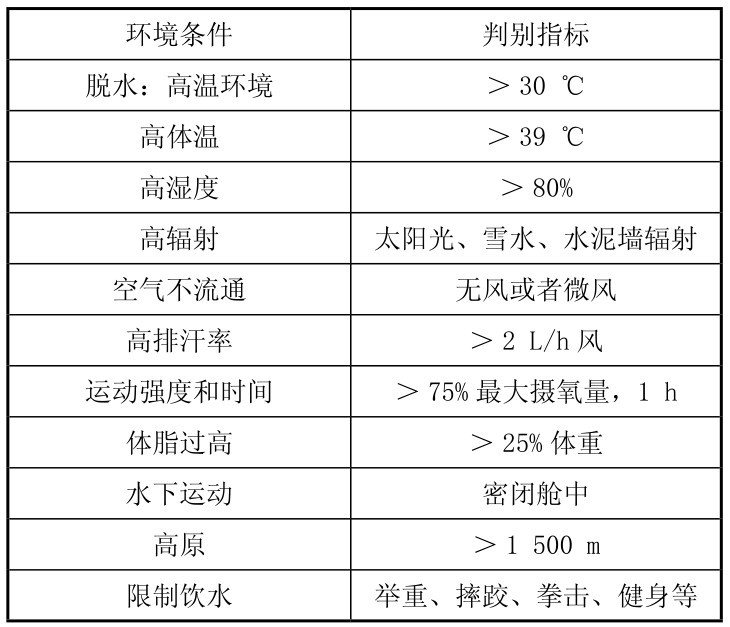
(引自冯炜权,1997)
1.合理的补水方法
运动员水的供给量应以补足失水量以保持水平衡为原则。补水的同时适当补充盐,目前,多补充低浓度的糖—电解质溶液。综合目前研究,认为结合运动前、中、后补水可以达到最佳效果。
①运动前。运动前30 min补水300~500 m L,可减少体温升高,延缓脱水发生;运动员在运动中会感到比较舒适;赛前补水,能够减轻途中的脱水程度。
②运动中。运动中补液一般是不够的,同时还会受到比赛规则和比赛条件的限制;在不同的项目和运动强度下,补液后的胃肠反应也是必须考虑的因素;在长时间耐力运动中,每隔15~20 min补液100~300 m L,但每小时补液总量不宜超过800 m L,否则,会因为吸收不了而出现恶心、呕吐现象。
③运动后。运动后补液,应该遵循少量多次的原则。如果饮水过多,容易增加心、肾负担,并影响食欲。此时的补液应以糖—电解质溶液为主,可以帮助血浆容量尽快恢复。如果只是补充凉开水,容易引起尿量增加,反而延缓身体复水和体内电解质的平衡过程。要达到机体短时间内(<6 h)完全复水,通常需补充体液流失量1.5倍的液体,同时液体中应含有一定量的电解质,特别是氯化钠,以补充运动中通过汗液丢失的盐分。
2.补充水、电解质的饮料
运动散热的主要方式是汗液,大量出汗导致的脱水会不同程度地损害运动能力,并会增加热疾病发生的危险性,补充糖—电解质饮料可以改善运动能力,使用运动饮料进行补充是最佳选择。运动饮料中的电解质主要是钠离子和氯离子。运动饮料的定义为:营养素及其含量能适应运动或体力活动人群的运动生理特点,能为机体补充水分、电解质和能量,可被迅速吸收的饮料(GB 15266—2009),并规定了理化指标和维生素添加剂标准,见表3.7。
表3.7 运动饮料国家标准理化指标
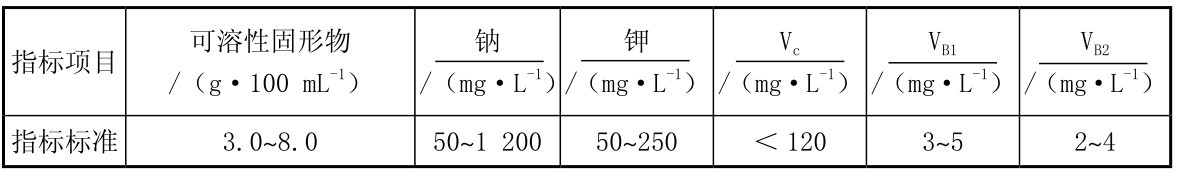
运动中补液的目的是为了保持和提高运动能力,运动饮料提供糖类、水和电解质。还要考虑饮料的风味、口感,它不但影响运动员饮用该饮料的数量,还影响运动员对该饮料的喜爱程度。饮用低浓度的糖—电解质溶液比单独补水的效果好,最佳的钠离子浓度为40~60 mmol/L,可以促进水的吸收和维持血容量。长时间运动时补钠可以补充汗液中钠的流失,维护血钠浓度和血浆渗透压。
知识拓展
运动性低钠血症
在长时间、长距离的运动中发生的低钠血症,称为运动性低钠血症(exertion blood hyponadium)。有症状的低钠血症可见于马拉松、超长马拉松、公路自行车、军事训练以及一些娱乐活动。在体育活动中,发生低钠血症的症状严重程度与血清钠离子浓度降低的程度和速度有关,表现为恶心、呕吐、头痛,甚至抽搐、昏迷。如果低钠血症是经过数小时缓慢形成,一般很少导致脑水肿,也较少出现严重不良症状。长时间运动产生的低钠血症,主要原因是在多个小时内,在大量出汗的情况下过量饮用低渗透压液体,钠的损失不能得到补充,因此属于稀释性低钠血症。饮用含盐饮料或食用含盐食物可以预防运动性低钠血症。
探索与思考
1.人体运动中为什么体温会升高?人体如何在运动中进行体温调节?
2.运动饮料国家标准中,为什么要含有这些成分?每一种成分都在运动中起什么作用?
小结
在运动中,人体通过体表蒸发汗液,是调节运动中体温升高的主要途径。特别是在长时间大强度运动中,随着汗液的蒸发,伴随体内水分和大量电解质丢失,造成运动脱水和电解质特别是钠和钾大量丢失,降低肌肉力量,降低运动能力;在炎热高湿度的环境中运动,脱水和电解质紊乱会发生得更快,并易造成中暑;长时间的运动还会由于钠的大量丢失诱发“运动性低钠血症”,危及生命。因此,在训练和比赛中,运动员应该养成习惯,规范地补充运动饮料。
本章小结
通过这一章知识内容的学习,我们初步了解了运动中骨骼肌内能量生成的基本过程,包括ATP-CP系统、糖酵解系统(合称无氧代谢供能系统),糖有氧氧化系统、脂肪有氧氧化系统(合称有氧代谢供能系统),不同的能量供应方式主要取决于运动强度(包括肌肉力量大小、肌肉收缩速度)和运动时间,与肌纤维动员类型密切相关,无氧代谢供能主要发生在快肌纤维,有氧代谢供能主要发生在慢肌纤维。在各种运动方式的实际运动中,各种能量供应方式均在运动中起重要作用,区别主要在于分别在不同的强度和时间内所起作用的比例不同,因此在实际运动中是连续的供能过程。
运动中伴随能量生成与释放,骨骼肌有大量热量释放,通过血液循环散发,最后主要以出汗的方式散发热量,造成体内水分大量丢失,同时伴随大量电解质特别是钠离子的丢失,维持体内水分和电解质平衡是维持内环境稳定、保持运动能力、减轻运动疲劳的重要生理机制。
维生素在运动训练中与心脏、大脑、骨骼肌等器官中的能量代谢和物质代谢密切相关。特别是B族维生素和维生素C直接参与运动中骨骼肌内的能量代谢,相关内容可以通过查阅资料进一步学习。
需要说明的是,运动中,通过大脑运动皮层甚至心理意志的作用,调节、维持或加强运动神经元的活动,运动神经元则决定了动员募集的肌纤维数量和肌纤维收缩频率的动态变化,从而调节、维持或加强肌肉的力量。这一复杂的神经过程,对运动中能量消耗的速率、总能量消耗起决定作用;相应地,运动中人体的呼吸和氧供应、肾上腺素和皮质激素等运动中促进能量生成的激素与神经作用协调,调节运动中能量的生成方式和供能速率,整合能量代谢系统,使不同的能量代谢系统相互促进,而非单独起作用,如肌酸-磷酸肌酸代谢供能系统促进糖酵解系统的供能,糖酵解代谢供能系统的产物可以激活有氧氧化系统,肌酸-磷酸肌酸代谢供能系统通过能量运转促进有氧氧化代谢供能系统的供能。另外,运动中肌肉内蛋白质分解增多,包括收缩蛋白质成分和其他功能性蛋白质成分,虽然其中某些氨基酸如枝链氨基酸可以通过三羧酸循环分解产生能量,但是在正常情况下其提供的能量只是总能量消耗的极少部分,因此一般不考虑蛋白质在运动中的供能。
在运动后人体的恢复中,能源物质特别是磷酸肌酸和肌糖原再合成、蛋白质的恢复合成以及其他多种生理机能的恢复和细胞内环境平衡的恢复和提高,均需要消耗能量,肌肉中这些能量一般大部分来自于脂肪酸的有氧氧化,其他器官中主要来自于葡萄糖(从血液中摄取)的有氧氧化,这是运动生理学中提到的“运动后过量氧耗”的主要原因。同样,胰岛素在运动后恢复中促进各个器官包括骨骼肌组织从血液中吸收各种营养物质(葡萄糖、氨基酸、脂肪酸、维生素等),用于能源物质和蛋白质的恢复合成,促蛋白质合成激素主要是雄性激素(如睾酮)以及生长激素促进运动后蛋白质的恢复合成和超代偿。这些是激素在运动后对物质合成代谢恢复的调节作用的重要表现。
学习这一章的知识内容后,可以结合前两章的内容进行回顾,进一步加深对相关内容的理解。同时,也为后面章节的知识内容学习奠定了基础。
练习题
一、名词解释
1.磷酸原
2.糖酵解
3.超代偿
4.糖的有氧分解代谢
5.脂肪动员
二、填空题
1.人体肌肉内存在3种不同类型的能量生成方式______、_______和______,供能过程消耗的主要底物分别为______、________和__________。
2.骨骼肌在运动过程中的直接能量供应者是________。
3.磷酸原系统供能可维持最大强度运动的时间是_______;糖酵解功能系统是最大强度运动时间范围为______的最主要供能方式。
4.运动员的无氧代谢调节包括3个方面_______、_________和________。
5.脂肪有氧氧化分解代谢可以分为5个步骤:_______、_________、________、_______和_______。
6.糖酵解的终产物有_______、________。
7.糖有氧氧化的终产物有_______、_________和_________。
8.人体内糖的存在形式有_______、_________和_______。
9.汗液的主要成分与细胞外液相同,主要是_____和________。
10.运动中补充的运动饮料主要含有_______、_______和________,补充的目的是为了保持和提高运动能力。
三、问答题
1.举例说明运动训练能否提高磷酸原供能系统能力并解释原因及其调节过程。
2.糖原的储备和恢复是决定运动员无氧代谢能力的关键环节,在实际训练和比赛中,运动员应该如何做才能提升糖原的储备量及其恢复能力?
3.试比较100 m运动员和马拉松运动员在比赛过程中身体能量供应方式的异同点。
4.为什么补充肉碱在一定程度上可以达到节省肌糖原的目的?
5.为什么说运动员乳酸清除能力的提高可以缓解疲劳和提高运动成绩?
6.影响运动时脂肪供能的因素有哪些?
7.简述运动后糖原的恢复规律。
8.简述失水对运动能力的影响。
9.论述运动中水盐代谢与体温调节的关系。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












