



第二节 中心法则
孟德尔对遗传因子行为的论述使遗传的染色体假说迅速流传开来,而摩尔根为遗传的染色体假说提供了一系列证据。人们认识到,染色体就是遗传物质。然而染色体中包括两种物质:核酸和蛋白质,它们之中哪一种才是遗传信息的承载者?包含了遗传信息的遗传物质究竟如何发挥作用?
DNA是遗传物质的第一个证据来自1944年美国微生物学家艾弗里(Osward Avery)完成的肺炎链球菌转化实验(图6-3)。

图6-3 肺炎球菌转化实验
肺炎链球菌能引起人的肺炎和小鼠的败血症。这种细菌有很多不同菌株,光滑型(S)菌株能引起疾病。而粗糙型(R)菌株不能引起疾病。1928年,英国科学家Griffith发现用热杀死的S型细菌和活的无毒的R型细菌注射到小鼠中,能使很多小鼠患败血症死亡,而且从它们的心脏血液中找到活的S型细菌。活的R型细菌、死的S型细菌分别注射时,都不引起败血症。这说明热杀死的S型细菌有一种物质或转化因素能够进入R型细菌,使其转化为S型细菌。1944年,Avery和他的同事分离出热杀死的S型细菌中的各种成分,包括蛋白质、核酸、糖类、脂类等在离体条件下进行转化,发现只有DNA组分能够把R型细菌转变为S型。如果用DNA酶处理,使DNA分解,转化效率大大下降。这些实验结果第一次证实DNA就是遗传物质。
那么作为遗传物质的DNA分子究竟如何发挥它的作用?生物体内遗传信息究竟是如何流动的? 1958年,克里克提出了中心法则,描述了遗传信息传递的规律,包括由DNA到DNA的复制、由DNA到RNA的转录和由RNA到蛋白质的翻译等过程。20世纪70年代逆转录酶的发现,表明还有由RNA逆转录形成DNA的机制,是对中心法则的补充和丰富(图6-4)。
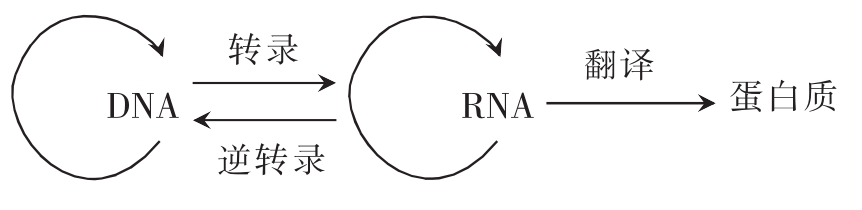
图6-4 修改后的中心法则
一、复制
DNA复制(DNA replication)是指以亲代DNA分子的一条单链为模板(template),根据碱基互补配对原则,合成一个与亲代DNA具有相同序列的新的子代DNA分子的过程。细胞分裂前均需要进行DNA复制,以保证分裂后每个子细胞中均有一套完整的遗传物质。
1.半保留复制与半不连续复制
DNA复制的特征之一是半保留复制(semiconservative replication)。沃森和克里克在提出DNA双螺旋结构模型时就对DNA的复制过程进行了探讨。他们认为DNA分子由两条链组成,两条链上的碱基G只能与C配对,A只能与T配对,所以两条链是互补的,每条链上都含有合成它的互补链所需要的全部信息。他们当时就推测,DAN分子在复制时将解开双螺旋结构,分别以两条单链为模板合成新链,这样新形成的两个DNA分子与原来的DNA分子的碱基顺序完全一样。现代生物学已经证明他们的推测是正确的:在DNA分子复制时,亲代DNA双链解开,每条单链作为模板指导合成其互补链,新合成的两个子代DNA分子与亲代DNA分子碱基序列完全一样。且其中一股单链来自亲代DNA,另一条单链是新合成的,这种复制方式称为半保留复制。
DNA复制的另一个特征是半不连续复制(semidiscontinuous replication)。半不连续复制是指DNA复制时,前导链上DNA的合成是连续的,后随链上是不连续的,故称为半不连续复制(图6-5)。

图6-5 半不连续复制
自然界中的DNA聚合酶只能以5'→3'方向合成子代DNA链,即模板DNA链的方向必须为3'→5'。而根据右手双螺旋结构模型,DNA分子中的两条单链是反向平行的。所以分别以两条亲代DNA单链作为模板合成子代DNA链时的方式是不同的。以3'→5'方向的亲代DNA链作模板的子代链在复制时基本上是连续进行的,其子代链的聚合方向为5'→3',这一条链被称为前导链(leading strand)。而以5'→3'方向的亲代DNA链为模板的子代链在复制时新链的聚合方向也是5'→3',因此子代DNA只能以短链的方式一段段合成,这些片段称为冈崎片段(Okazaki fragment)。这种新链合成的方式是不连续的,因此新DNA分子合成过程中,两条新生成的链一条是连续合成的,一条是不连续合成的,这种DNA复制方式被称为半不连续复制。以冈崎片段方式合成的新链需在前导链合成一短时间,模板单链拥有一定长度时才能开始合成冈崎片段,因此这条链被称为随后链(lagging strand)。
由于后随链只能以冈崎片段的方式合成,后随链成为中间掺有RNA片段的断裂的DNA分子,生物体首先利用DNA聚合酶的5'→3'外切酶活性清除冈崎片段的引物,其后利用DNA聚合酶的5'→3'聚合酶活性填补缺口,最后用DNA连接酶连接DNA分子的主链,完成复制过程。
2.参与DNA复制的主要蛋白质
为完成DNA分子的复制,大量蛋白质参与了复制过程。以下阐述几种参与DNA分子复制的主要蛋白质。
拓扑异构酶(topoisomerase)和解链酶(helicase):拓扑异构酶是生物体中用于处理空间拓扑结构的一种酶,通常认为它利用DNA分子的负超螺旋结构,在DNA分子第一个双螺旋结构被解开的过程中起着非常重要的作用。解链酶可通过水解ATP供能来解开双链,每解开一对碱基,需水解2分子ATP。
单链结合蛋白(single-strand binding protein,SSB):DNA双螺旋解开成单链后,SSB便牢固地结合到分开的单链上,防止它们重新形成双螺旋,同时也防止解链后的DNA分子被核酸内切酶水解。原核生物的SSB与DNA的结合表现出明显的协同效应,当第一个SSB结合后,其后的SSB与DNA的结合力可大大提高,且结合迅速扩展,直到全部单链DNA都被SSB覆盖。而真核生物的SSB没有协同效应,在DNA复制过程中不断地结合、脱离,直到复制完成。
RNA聚合酶(RNA polymerase)和引发酶(primase):DNA聚合酶没有从无到有合成DNA的能力,在有机体中,合成DNA首先必须合成一小截RNA引物。目前大多数研究者认为前导链和后随链的RNA引物合成是有区别的,前导链上的RNA引物由RNA聚合酶合成(部分研究者认为RNA聚合酶合成RNA分子的过程只是转录激活),后随链上冈崎片段的引物由引发酶与相关的蛋白结合形成引发体后合成。
DNA聚合酶(DNA polymerase):催化dNTP聚合到核酸链上的酶。DNA聚合酶具有5'→3'的聚合活性,能催化四种dNTP以磷酸二酯键一个一个地接到DNA链上去。此外,DNA聚合酶还具有3'-5'外切核酸酶活性,当新合成的DNA单链3'端出现非配对碱基时,该功能可以切除错误碱基起到校对功能。大肠杆菌DNA聚合酶I还具备5'→3'核酸外切酶活性,可以用以去除冈崎片段中的引物。
DNA连接酶(DNA ligase):催化相邻DNA片段间的连接,把有缺口的3'-OH末端与相邻核苷酸5'-磷酸连接起来形成磷酸二酯键,连接反应是耗能的。大肠杆菌的DNA连接酶以NAD+为能量来源,动物细胞和某些噬菌体以ATP为能量来源。DNA连接酶在后随链的合成过程中起着重要的作用。
3.复制起始位点与复制叉
复制起始位点(origin of replication)是指DNA分子上开始进行复制的部位。DNA分子的复制起始不是随机的,只能从特定的区域开始。作为第一个DNA分子双链被打开的区域,复制起始原点在结构上有很多特点,是分子生物学家特别关注的区域之一。原核生物每个DNA分子只有一个复制起始点,真核生物DNA分子有多个复制起始点。每个复制起点到终点间的DNA复制区域称复制子(replicon)。原核生物的基因组只有一个复制子,而真核生物染色体为多复制子。复制时,双链DNA由复制起始位点处打开双螺旋结构,沿两条张开的单链模板合成DNA新链,两侧形成的Y型结构称为复制叉(replication fork)(图6-6)。

图6-6 复制起始位点与复制叉
大肠杆菌在对数生长期时,DNA分子上经常出现多复制叉结构。DNA分子的一轮复制还没有结束,新生成的DNA分子双链已经打开,开始新一轮的复制。这是由于在对数生长期大肠杆菌DNA复制速度极快造成的。实际上,对数生长期的大肠杆菌内常有多个主DNA分子,原因是DNA分子复制的速度快于细胞分裂的速度。
二、转录
转录(transcription)是遗传信息从DNA到RNA的转移。即以双链DNA中的一条链为模板,以腺三磷(ATP)、胞三磷(CTP)、鸟三磷(GTP)和尿三磷(UTP)4种核苷三磷酸为原料,在RNA聚合酶催化下合成RNA的过程。转录的产物包括mRNA以及非编码RNA(tRNA、rRNA)等。同时我们将作为模板的DNA单链称为模板链(template strand),与转录出的RNA序列相同的DNA单链称为编码链(coding strand)。转录起始位点是指DNA分子中开始RNA转录的位置。常把起点前面,即5'末端的序列称为上游,把其后3'末端的序列称为下游。在描述碱基位置时,起点为+1,下游方向依次为+ 2、+3,上游方向依次为-1、-2、-3等等。
1.RNA聚合酶和启动子
RNA聚合酶(RNA polymerase)是以一条DNA链或RNA链为模板催化由核苷-5'-三磷酸合成RNA的酶。催化转录的RNA聚合酶是一种由多个蛋白亚基组成的复合酶,通常体积非常庞大,能横跨DNA分子上的几十个碱基,这种空间结构上的庞大性有利于进行基因表达调控。
大肠杆菌的RNA聚合酶由五个亚基组成(图6-7),含有α,β,β',δ4种不同的多肽,其中α为两个分子。大肠杆菌的RNA聚合酶可分为核心酶和δ因子两部分,δ因子的作用是识别转录的起始位置,并使RNA聚合酶结合在启动子部位。α2ββ'四个亚基组成核心酶(core en-zyme)负责链的延伸。转录开始时,δ因子首先识别启动子并与启动子相结合,召唤核心酶组成全酶(holoenzyme)。转录开始后,δ因子被释放,链的延伸则由四聚体核心酶催化。核心酶的四个亚基中,α亚基与RNA聚合酶的四聚体核心(α2ββ')的形成有关,β亚基含有核苷三磷酸的结合位点,β'亚基含有与DNA模板的结合位点。利福平(rifampicin)和利福霉素(rifamycin)能结合在β亚基上而对此酶发生强烈的抑制作用。它抑制RNA合成的起始而不抑制其延长。链霉素(streptolydigin)能抑制RNA链的延长。肝素能和β'亚基结合,从而在体外抑制转录作用。

图6-7 大肠杆菌的RNA聚合酶
真核生物的RNA聚合酶更为庞大,主要分三类:RNA聚合酶Ⅰ存在于核仁中,转录rRNA;RNA聚合酶Ⅱ存在于核质中,转录大多数基因,需要“TATA”框;RNA聚合酶Ⅲ存在于核质中,转录很少几种基因,如部分tRNA基因、5SrRNA基因等。
启动子(promoter,P)指DNA分子上能与RNA聚合酶结合并形成转录起始复合体的区域。原核生物的启动子有一系列保守结构:
(1)-10区,又称Pribnow box或TATA box,位于转录起始位点上游5~10bp,一般由6~8个碱基组成,富含A和T。启动子来源不同,Pribnow盒的碱基顺序稍有变化。它是RNA聚合酶的牢固结合部位。
(2)-35区,位于转录起始位点上游35bp处,故称-35区,一般由10个碱基组成。它是RNA聚合酶的初始结合部位。
(3)-10区与-35区之间核苷酸数目,强启动子一般为17±1bp。
真核生物的启动子结构复杂,结构保守性相对较弱,但也有一些相对保守的元件,如-25~-35区含有TATA box、-70~-80区含有CAAT box、-80~-110区含有GC box等。习惯上TATA区上游的保守序列称为上游启动子元件(upstream promoter element,UPE)或上游激活序列(upstream activating sequence,UAS)。真核的RNA聚合酶通常需要和转录因子(transcription factor)相协调才能和启动子结合。
2.原核生物的转录
原核生物没有成形的细胞核,其转录过程特点鲜明。
(1)转录单元为多顺反子结构
转录单元(transcription unit)是一段从启动子开始到终止子(terminator)结束的DNA序列。真核生物每个基因都有自己单独的启动子,每个转录单元通常只包含一个基因。而原核生物的结构基因经常呈多顺反子(polycistron)形式排列。将几种不同基因的mRNA连在一起,相互之间由一段短的不编码蛋白质的间隔序列所隔开,这种mRNA叫做多顺反子mRNA。这样的一条mRNA链含有指导合成几种蛋白质的信息。如乳糖操纵子中3个与代谢乳糖相关的结构基因被顺序排列在一起,在其前端加上一个启动子,三个基因的转录顺序进行,转录出的mRNA经过翻译生成三种蛋白质。
(2)mRNA寿命短
原核生物mRNA寿命很短,一般只有几分钟,最长也只有数小时(RNA噬菌体中的RNA除外)。有观点认为,转录开始后1分钟,mRNA的降解就开始了。而真核生物mRNA的寿命则较长。
(3)转录和翻译偶联
原核生物mRNA的转录与翻译一般是偶联的,即转录尚未完毕,蛋白质的翻译、合成就已开始。而真核生物转录的mRNA前体则需经后加工后才可以进行翻译。
由于没有核膜,原核生物的核糖体可以进入核区,与刚刚从RNA聚合酶中释放出来的mRNA分子相接触,在转录还没有结束时就开始翻译。翻译的过程中核糖体会延着mRNA向前移动,空出mRNA分子和核糖体结合的部位,第二个核糖体又可以和mRNA相结合开始翻译。这样,在新生成的mRNA链上将排列一系列核糖体,形成多核糖体结构。在某个瞬间,正在转录的mRNA分子的核糖体结合部位遭到攻击而被降解后,下一个核糖体就无法再和mRNA分子相结合。因此,原核生物转录和翻译的偶联对原核生物来说,有着重要的意义。
3.真核生物的转录
由于存在核膜,核糖体无法和正在转录的mRNA分子相接触,真核生物的转录与翻译无法偶联。因此,真核生物的转录与原核生物有很大区别,特别是真核生物mRNA分子在转录后有一个复杂的加工过程(图6-8):
(1)真核生物mRNA分子的5'端存在“帽子”结构。虽然真核生物的mRNA的转录通常以嘌呤核苷酸三磷酸(pppA或pppG)开始,但成熟mRNA 5'端的一个核苷酸总是7-甲基鸟核苷三磷酸(m7GpppAGpNp)。这是因为在转录开始后很短的时间内,正在转录的mRNA分子的5'端在就在鸟苷酸转移酶的催化下,加上一个倒扣上去的甲基化的鸟嘌呤。根据+ 1和+2位点两个核糖核苷酸上甲基化情况的差异,真核生物的5'端“帽子”可分为零号帽(cap 0)、1号帽(cap 1)和2号帽(cap 2)。5'加帽的功能主要有:①使mRNA免遭末端核酸酶的破坏,增加mRNA分子的稳定性。②能与核糖体小亚基识别,促使mRNA和核糖体结合。
(2)绝大多数真核生物的mRNA分子具有poly(A)尾巴。除组蛋白基因外,真核生物mRNA分子的3'末端都有poly(A)序列,其长度因mRNA种类不同而变化,长度通常在100~200个碱基左右。poly(A)尾不是由DNA编码的,而是转录完成后加上去的。有研究表明,转录完成后的mRNA分子首先由一个特异性酶识别切点转录末端上游方向13~20碱基的加尾识别信号AAUAAA以及切点下游的保守顺序GUGUGUG,把切点下游的一段切除,再由poly(A)聚合酶(又称RNA末端腺苷酸转移酶)催化,以ATP为前体,合成poly(A)尾巴,如果这一识别信号发生突变,则切除作用和多聚腺苷酸化作用均显著降低。mRNAPoly(A)尾的功能是:①可能有助mRNA从核到细胞质转运;②避免在细胞中受到核酶降解,增强mRNA的稳定性。
有趣的是,虽然除了组蛋白基因编码的产物外,真核生物mRNA分子的3'末端都有poly(A)序列,但由于组蛋白在细胞内的用量较大,约有1/3的真核mRNA3'末端没有poly(A)序列。
(3)内含子的剪切
真核生物的基因通常是断裂基因,意为真核生物基因中编码产物的部分常常被不编码产物的部分分隔开。因此在基因表达的过程中,伴随着RNA的剪接过程。在mRNA前体分子剪接过程中被切除的部分所编码的信息不出现在蛋白质产物中,称之为内含子(intron);而在剪接过程中被保留下来,所编码的信息出现在蛋白质产物中的序列,称之为外显子(exon)。因此,在真核生物中,由转录生成的原初转录产物——核不均一RNA(heterogeneous nuclear RNA,hnRNA),需经过5'加帽、3'加尾、内含子的剪接等步骤才能成为成熟的mRNA,在细胞质中进行翻译。
可变剪接(alternative splicing)是近年来越来越引起分子生物学家注意的现象。它是指同一前体mRNA分子,可以在不同的剪接位点发生剪接反应,生成不同的mRNA分子,最终产生不同的蛋白质分子的一种RNA剪切方式。可变剪接的存在使一条DNA序列可最终编码2个以上的蛋白质,使基因编码产物的效率大大增强。可变剪接刚被发现时,被认为是在少数基因中出现的个别现象,随着人类基因组计划的完成,研究者吃惊地发现,人类基因组中有50%左右的基因可能存在可变剪接现象,而部分学者甚至认为这一比例高达75%。
RNA编辑(RNA editing)是RNA分子加工时出现的修饰现象。mRNA因核苷酸的插入、缺失或替换而改变了源自DNA模板的遗传信息,翻译出不同于基因编码的氨基酸序列。RNA编辑在mRNA水平上改变遗传信息,同样可使一个基因编码几个不同的蛋白质。
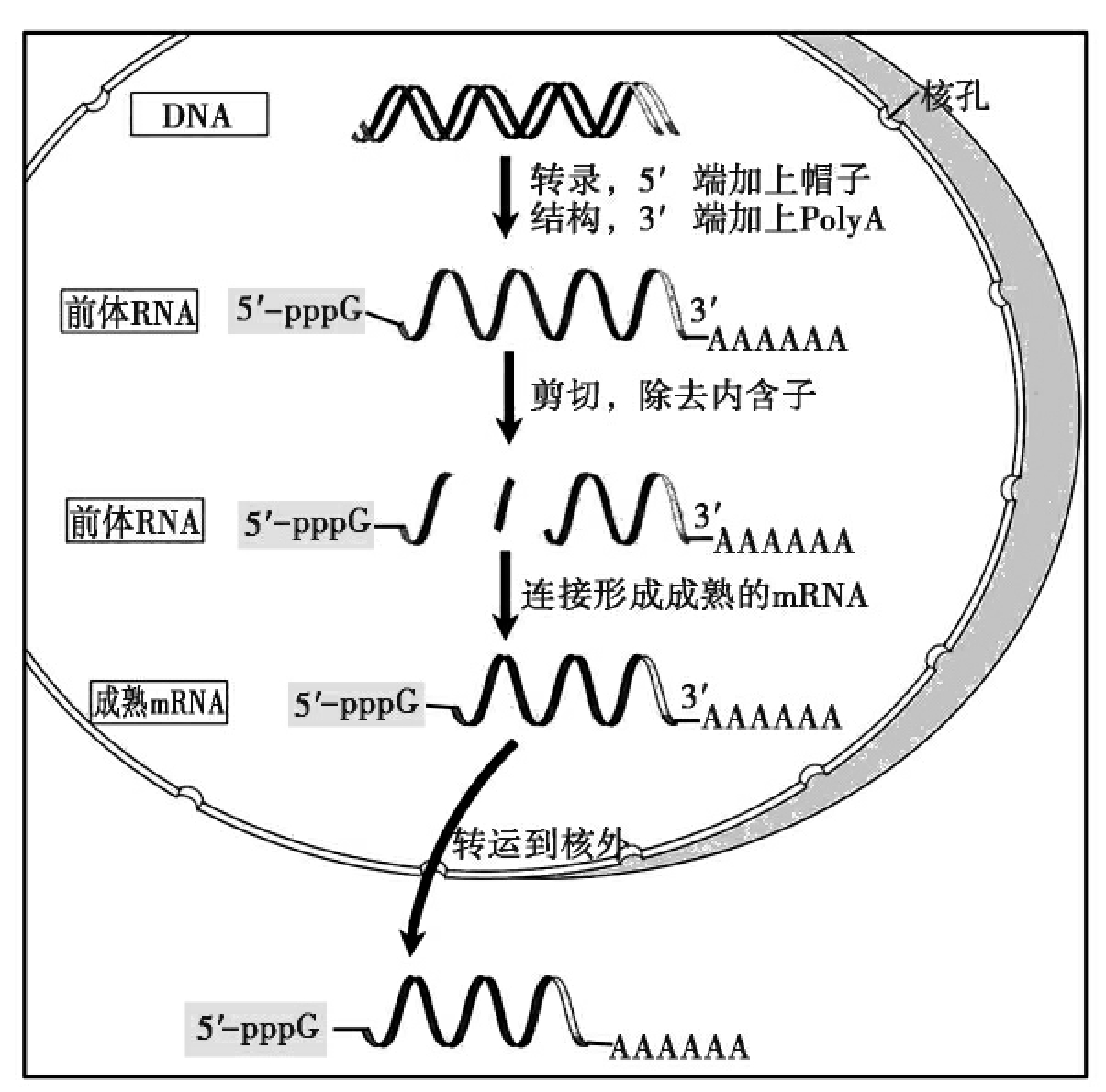
图6-8 真核基因转录后加工过程
二、蛋白质的生物合成——翻译
翻译(translation)是指将mRNA分子中由碱基排列顺序组成的遗传信息通过密码子破译的方式转变成蛋白质中的氨基酸排列顺序,合成蛋白质的过程。翻译是一个复杂的过程。细胞中的很多组分参与了翻译,大量能量也消耗于蛋白质合成的过程中。总的来说,核糖体是蛋白质合成的场所,mRNA是蛋白质合成的模板,转移RNA(transfer ribonucleic acid,tRNA)是模板与氨基酸之间的结合体。
1.三联体遗传密码子
mRNA上每3个核苷酸翻译成蛋白质链上的一个氨基酸,这3个核苷酸排列顺序称作遗传密码,也叫三联体密码(triplet code)。三联体密码子碱基的组合的可能性有64种,64种碱基的组合对应64种密码子(表6-3)。
64个密码子中,61个是编码氨基酸的密码子,其中包括两个起始密码子:一种是AUG(编码甲硫氨酸),一种是GUG(编码缬氨酸),另外还有UAA、UAG、UGA三个密码子不能编码任何氨基酸,是蛋白质合成的终止密码子。
表6-3 密码子表

从遗传密码子表中可以看到,61个遗传密码子除了甲硫氨酸和色氨酸外,每一个氨基酸都至少有两个密码子。同一种氨基酸具有两个或更多个密码子的现象称为密码子的简并性(degeneracy)。简并性使密码子中碱基即使被改变,仍然编码原来氨基酸的可能性大为提高,也使DNA分子上碱基组成有较大的变动余地。我们注意到,在兼并密码子中,第一位和第二位的碱基经常是相同的,不同的是第三位碱基。1966年,克里克根据立体化学原理提出的摆动学说(webblehypothesis)很好地解释了这一现象。摆动学说认为:处于密码子3'端的碱基与之互补的反密码子5'端的碱基(也称为摆动位置),由于存在摆动现象,所以使得一个tRNA反密码子可以和一个以上的mRNA密码子结合。
在简并密码子中,某些生物体似乎更偏爱使用其中某一个,这一现象称为密码子偏好性(codon usage bias)。例如,在酵母基因组中,48%的精氨酸由密码子AGA编码,而其余五种编码精氨酸的同义密码子(CGT、CGC、CGA、CGG和AGG)则以较低的频率被使用(每种10%左右)。果蝇使用编码精氨酸的密码子偏好性完全不同,使用密码子CGC的概率为33%,明显高于其他五种密码子(每一种的出现频率约为13%)。
除极少数情况外(线粒体基因组、极端环境中的微生物)外,自然界的生物体,无论是原核生物还是真核生物,都使用一套通用的遗传密码子。这是进行基因工程研究的基础,也表明生物起源于共同的祖先。
2.转移RNA
转运RNA(transfer ribonucleic acid,tRNA)是具有携带并转运氨基酸功能的一类小分子核糖核酸,主要是携带氨基酸进入核糖体,在mRNA指导下合成蛋白质。
tRNA分子的二级结构为三叶草模型。它由3个环和5个臂组成,包括:D环、反密码环、TΨC环,D臂、反密码臂、TΨC臂、受体臂臂、可变臂等。除可变臂和D环外,各种tRNA分子各个部位的核苷酸数目和碱基对基本上是恒定的。在这些环和臂中,最重要的是反密码子环(anticodon loop)和受体臂(accept stem)。
受体臂对面的单链环称反密码子环,该环含有由三个核苷酸残基组成的反密码子。翻译过程中反密码子在核糖体内与信使核糖核酸的密码子配对识别,参与蛋白质生物合成。自然界中稀有碱基出现的频率很低,但在tRNA中,特别是在反密码子环中反密码子的3'端邻近部位出现的频率较高,这有利于维持反密码子环的稳定以及密码子和反密码子形成正确的配对结合。
受体臂是由7个碱基对的配对区加上3'末端3~4个未配对碱基所组成的,3'末端的最后3个碱基永远是CCA。在携带氨基酸时,氨酰-tRNA合成酶在水解ATP的情况下催化氨基酸和tRNA结合,氨基酸结合部位在受体臂末端腺苷酸残基的2'或3'-羟基上。携带了氨基酸的tRNA叫氨酰tRNA。
第一个进入核糖体与mRNA起始密码子结合的tRNA叫起始tRNA,其余tRNA参与肽链延伸,称为延伸tRNA。结合相同氨基酸的tRNA称为同工tRNA。
3.核糖体
核糖体是细胞内一种核糖核蛋白颗粒(ribonucleoprotein particle),核糖体无膜结构,主要由RNA和蛋白质构成,功能是按照mRNA上的遗传密码将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。无论是原核生物还是真核生物,细胞质内均有大量的核糖体。原核细胞的核糖体较小,沉降系数为70S,相对分子质量为2.5×103 kDa,由50S和30S两个亚基组成;而真核细胞的核糖体体积较大,沉降系数是80S,相对分子质量为3.9×103~4.5×103 kDa,由60S和40S两个亚基组成。此外,脊椎动物的线粒体内还存在55S的核糖体。
要完成蛋白质的合成,核糖体上至少有5个活性中心:mRNA结合部位、结合或接受氨酰-tRNA的部位、结合或接受肽基tRNA的部位、肽基转移部位、形成肽键的部位(转肽酶中心)。从分工上来说,核糖体小亚基主要负责对模板mRNA的特异性识别与结合,核糖体大亚基主要负责携带氨基酸及tRNA、形成肽键。
4.肽链起始、延伸和终止
以细菌为例介绍肽链起始和延伸的基本过程:
(1)多肽链的起始:在起始因子等一系列蛋白质的作用下,核糖体小亚基识别mRNA的起始部位并和mRNA结合,在起始因子和GTP的帮助下,甲硫氨酰(蛋氨酸)—tRNA的反密码子识别mRNA上的起始密码AUG,与之互补结合进入小亚基的P位,接着大亚基结合上去,GTP水解,释放翻译起始因子。
(2)多肽链的延长:在延伸因子和GTP的帮助下,第二个密码子对应的氨酰—tRNA进入核糖体的A位,密码子与反密码子的氢键互补结合。在大亚基上的多肽链转移酶(转肽酶)作用下,供位(P位)的tRNA携带的氨基酸转移到A位的氨基酸后并与之形成肽键(—CO-NH—)。核糖体沿mRNA往前移动一位,失去氨基酸的tRNA脱离P位,随后会进入细胞质中。同时,原本处于A位的二肽基tRNA进入P位,将A位空出,与A位新的密码对应的新氨基酰-tRNA进入A位,转肽键把二肽转到此氨基酸上后形成三肽,核糖体又往前移动,由此渐进渐进,反复循环,多肽链得以顺利延伸。
(3)多肽链的终止与释放:肽链的延长不是无限止的,当mRNA上出现终止密码时(UAA、UAG和UGA),无对应的氨酰tRNA进入核糖体,肽链的合成停止,被终止因子识别,进入A位,抑制转肽酶作用,使多肽链与tRNA之间水解脱下,从大亚基中释放出来,离开核糖体,同时大小亚基与mRNA分离,小亚基可再与mRNA起始密码处结合,也可游离于胞质中,完成翻译的mRNA也可被降解。
5.翻译后加工
翻译后加工(posttranslational processing)指多肽链从核糖体释放后经各种修饰、处理,成为有活性的成熟蛋白的过程。翻译后加工主要包括一级结构的修饰和高级结构的修饰。
一级结构的修饰主要包括:(1)N端fMet或Met的切除。大多数生物的N端甲硫氨酸会在多肽链合成完毕前被切除。(2)特定氨基酸的修饰。包括磷酸化、糖基化、甲基化、乙基化、羟基化、羧基化等。(3)切除某些非功能片段。如信号肽等。蛋白质前体通过自我剪接去除内含子、连接外显子的过程称为蛋白质剪接(protein splicing),是目前分子生物学的一大研究热点。
高级结构的修饰主要包括:(1)二硫键的形成和辅基连接。(2)多亚基蛋白的亚基聚合。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










