



9.3.2 生物体内自由基的产生与清除
除了物理或化学因素可使生物体产生外源性自由基外,生物体内自由基尚可从非酶反应和酶反应中产生。在某些氧化还原反应中产生的中间物自由基如FMN或FAD半醌自由基,其寿命极为短暂,而且对机体无明显毒性。内源性自由基一般指氧自由基及其活性衍生物。
9.3.2.1 生物体内自由基的产生
A.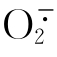 的产生
的产生
(1)非酶反应。在需氧的生物体内到处存在着O2。如果在O2参与的非酶反应中O2能从还原剂接受一个电子,就可以转变为。这类非酶反应在生物体内是存在的,现简述如下。
a)在O2存在的条件下,有些生物分子氧化时可产生。这些生物分子包括甘油醛、还原型核黄素、FMN与FAD、肾上腺素、四氢喋呤和半胱氨酸,GSH等巯基化合物。
b)在人体内,铁与蛋白质结合成血红蛋白、肌红蛋白、铁蛋白化合物。非蛋白质结合的铁是极少的,如在脑脊髓液中仅含有(2.2±1.3)μmol/L。在离体试验中已证明Fe2+可逐渐被空气中的O2氧化为Fe3+,同时O2转变为。
![]()
c)在非酶反应中以氧合血红蛋白转变为高铁血红蛋白时产生的最值得重视。因为每天人体中红细胞的氧合血红蛋白约有3%转变为高铁血红蛋白,表明循此途径可有相当量的产生。
![]()
d)在生物体内经常产生H2O2。如果H2O2不及时被清除,则有可能与过渡金属离子或其复合物如ADP-Fe2+反应,产生。
![]()
e)在氧化还原反应中醌类化合物产生的半醌自由基可与O2反应产生。
![]()
(2)酶反应。需要氧维持生命的生物包括细菌等微生物以及人,其体内一定会有氧代谢。在氧的代谢反应中常有酶促氧化与还原,这表明有些酶促反应中可能有的产生。
a)黄嘌呤或次黄嘌呤氧化为尿酸。在黄嘌呤氧化酶的催化下,黄嘌呤(或次黄嘌呤)可通过将单电子或双电子给予O2的方式氧化为尿酸。
![]()
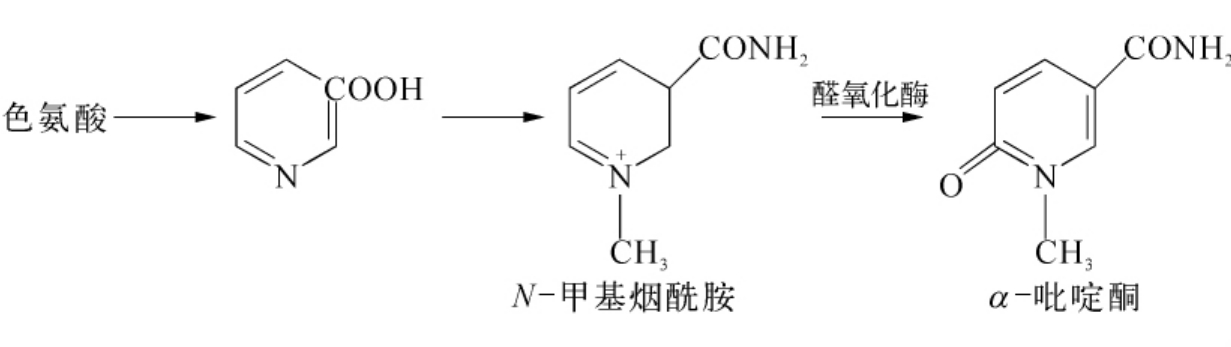
b)醛氧化酶。醛氧化酶也具有2个FAD、2个Mo及8个Fe。它还含有1个或2个分子的辅酶。
它的系统名称为醛-氧-氧化还原酶,其底物为醛与氧。该酶可催化醛与氧反应生成酸和。它对底物的专一性较黄嘌呤氧化酶更差,它尚可催化N-甲基烟酰胺转变为α-吡啶酮。
c)与线粒体呼吸链有关的某些酶。线粒体呼吸链为由NADH、黄素蛋白、铁硫蛋白、泛醌与细胞色素类组成的一系列氧化还原系统。
辅酶Q(泛醌UQH2)可转变为泛半醌(UQH),如氧化型细胞色素c(cytc3+)可使UQH2氧化成UQH自由基。它可自氧化为UQ,同时产生。
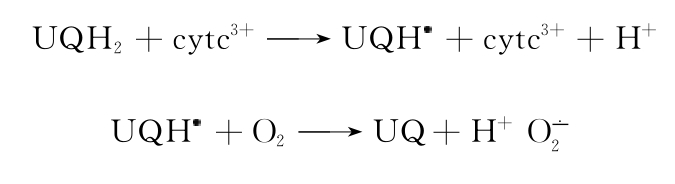
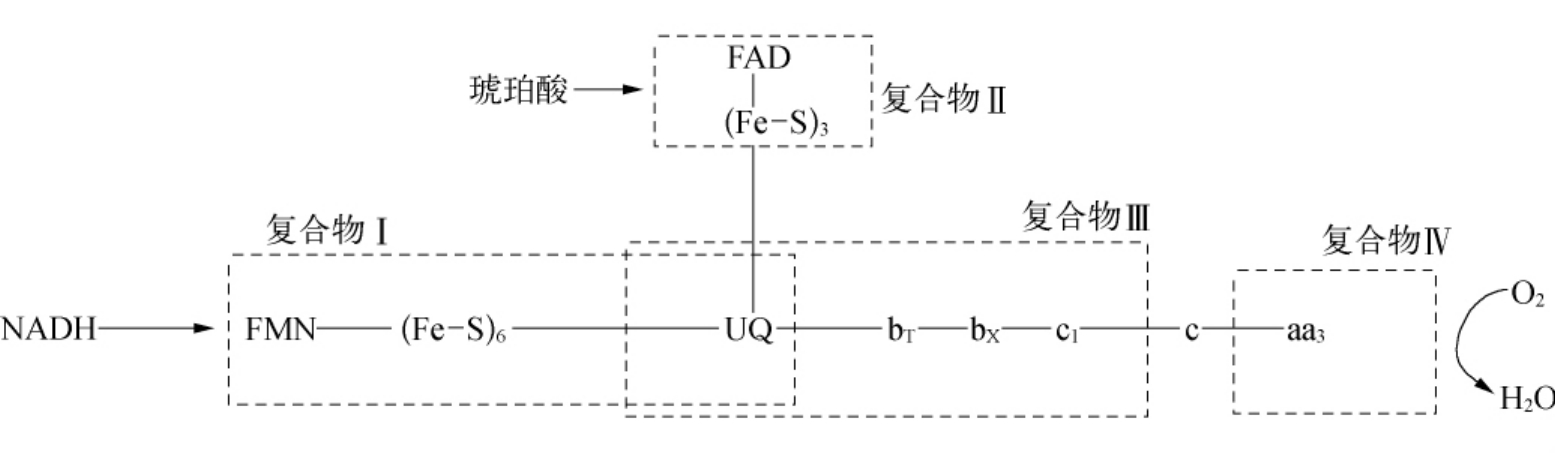
虽然复合物Ⅰ、Ⅱ、Ⅲ都涉及泛醌,可能产生,但其效能并不一致。一般认为,复合物Ⅰ与复合物Ⅲ都含有UQ,是产生的主要部位,而将电子传递至UQ的复合物Ⅱ却是次要的。
必须指出的是,的产生速率的大小决定于线粒体代谢的状态。在生理条件下,生物体内96%~99%的O2可通过细胞色素氧化酶的催化而还原为水,仅1%~4%的O2产生与H2O2。由于与H2O2可分别被SOD及过氧化氢酶、谷胱甘肽过氧化物酶清除,因此不会积累到足以损伤机体的浓度。但是在线粒体结构与功能受到影响时,与H2O2的生成可能增多,如果增加到超过生物体内的防御能力,就会发生活性氧对机体的损伤。
d)与微粒体电子传递系统有关的某些酶。微粒体不是细胞内存在的实体组织名称,而是用差速离心分离细胞器得到的核蛋白体与内质网。前者为蛋白质生物合成的场所。后者的功能虽随细胞不同而异,但都存在电子传递系统,其中某些酶反应产生了。
脂肪酸、甾醇、药物及许多异生素(xenobiotics)的氧化转变反应需要微粒体电子传递系统。后者包括了三种黄素蛋白(fp1、fp2及fp3)和其他黄素蛋白以及两种血红素蛋白(细胞色素b5与细胞色素P450)。
①黄素蛋白(fp1)。具有NADPH-细胞色素P450还原酶的功能,含有FMN与FAD两种辅基。在氧化还原过程中产生的自由基型黄素蛋白可在特殊情况下与O2反应生成,H或其衍生物如1O2可能参与脂类过氧化,或在血红素加氧酶作用下使血红素氧化裂解为胆色素。
②黄素蛋白(fp2)。又名NADH-细胞色素b5还原酶。它可将电子从NADH传递到细胞色素b5。
③黄素蛋白(fp3)。即所谓的混合功能胺氧化酶,其功能是参与许多二级及三级胺的氧化转变。
④其他黄素蛋白。在肝微粒体中除含有fp1、fp2与fp3三种黄素蛋白外,还含有其他的黄素蛋白,如DT黄递酶、葡萄糖酸丙酮氧化酶。
⑤细胞色素b5。为fp1与fp2两种黄素蛋白之间的电子传递蛋白。它具有脱饱和酶的作用,如可使硬脂酰-CoA转变为油酰-CoA。它在微粒体电子传递系统中的主要作用可能是作为氧化型细胞色素P450的电子供体。
⑥细胞色素P450。它是在许多有机化合物的氧化转变中起到氧活化与底物相互作用的关键酶。其辅基为Fe3+-血红素。
e)其他酶体系。产生的酶反应尚有多种酶。二氢乳清酸脱氢酶、NADPH氧化酶、铁氧化还原蛋白-NADPH还原酶、红氧化还原蛋白-NADPH还原酶、非特异性过氧化物酶、硝基丙烷双氧酶、吲哚胺双氧酶、色氨酸双氧酶、半乳糖氧化酶等。
①二氢乳清酸脱氢酶。在二氢乳清酸脱氢酶作用下,二氢乳清酸可转变为乳清酸。由于该系统含有泛醌,可在催化反应中转变为泛半醌,泛半醌与O2反应可产生氧化型泛醌及。

②NADPH氧化酶或NADH氧化酶。该酶存在于嗜中性粒细胞及巨噬细胞的浆膜内。在NADPH氧化酶或NADH氧化酶作用下,可产生,同时NADPH或NADH可氧化为NADP+或NAD+。
③铁氧化还原蛋白-NADPH还原酶。该酶的底物为铁氧化还原蛋白及NADPH。在O2存在时其他催化产物除外,尚有NADP+及还原型铁氧化还原蛋白。
④红氧化还原蛋白-NADPH还原酶。红氧化还原蛋白-NADPH还原酶的催化作用虽与铁氧化还原蛋白-NADPH还原酶类似,但底物之一红氧化还原蛋白与后者不同。在催化反应中都产生。
⑤过氧化物酶。在植物与细菌中含有非特异性过氧化物酶。该酶的辅基为Fe2+-原卟啉Ⅸ,其分子结构式如图9-9所示。
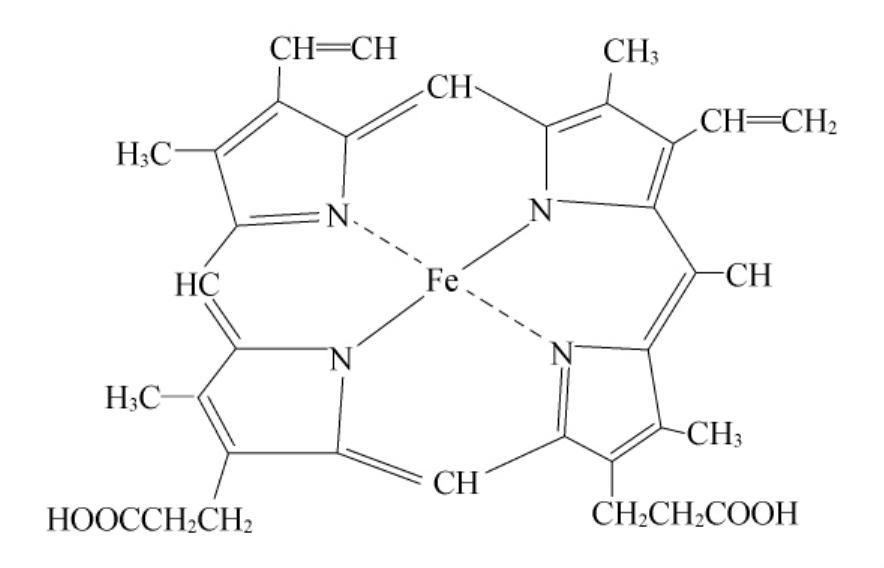
图9-9 Fe2+-原卟啉Ⅸ的分子结构
在H2O2存在的条件下,它可作用于愈创木脂醇、联苯胺与邻联二茴香胺以及其他底物。设底物为SH2,则其反应式可表示如下:
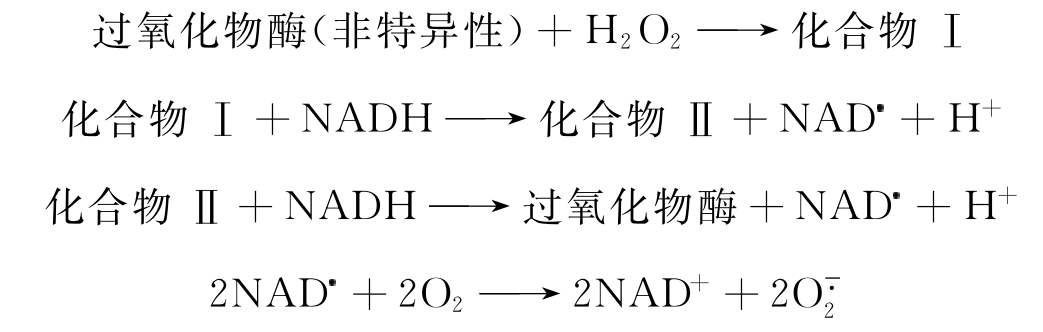
NAD是一种自由基。它与O2反应可产生。其反应速率是很高的,k2为1.9×109mol·L-1·cm-1。
⑥硝基丙烷双氧酶。该酶可催化2-硝基丙烷氧化为丙酮,同时产生。SOD可抑制其氧化反应。
⑦色氨酸双氧酶。该酶可作用色氨酸,但不能作用其他底物。在催化过程中既产生了,又利用了O2,其产物为N-甲酰-犬尿氨酸。
B. 的产生
的产生
的化学性质非常活泼,寿命极短,产生部位常为其作用部位。如果在生物体内重要生物分子如核酸不处在产生部位,则不会受到损伤。虽然至今难于用现有的技术直接检查出生物体内的,但是已有很多事实间接证明生物体内确有产生。
b)某些药物在体内代谢中可能产生。
①羟化的化学药物,如6-羟基多巴胺、6-氨基多巴胺、6,7-二羟基色胺、5-羟基巴比妥酸、四氧嘧啶进入生物体内,可产生。证据是:其一,与甲硫基丙醛反应可产生C2H4;其二,被清除剂抑制。
②带有醌型的抗肿瘤药物,如博莱霉素、丝霉素B、C与链黑霉素进入生物体内,可产生。其证据是它们都可与捕获剂DMPO或PBN(phenyl butyl nitrone)结合。
c)某些酶或其复合体系可产生。
②前列腺素合成酶。在该酶的催化反应中产生的可使酶发生自身失活,但加入甲硫基丙醛不仅能除去,使酶自身失活受到抑制,而且通过甲硫基丙醛与反应产生的C2H4,间接证明酶促反应中确有产生。
③鸟嘌呤环化酶。在酶促过程中所需的可在反应中产生,其主要证据是其反应可被清除剂抑制。
④微粒体乙醇氧化体系。在肝微粒体中存在乙醇脱氢酶,它可使乙醇氧化为乙醛,同时产生H2O2。后者通过Fenton反应产生,证据是:其一,用脱铁敏(desferrioxamine)使Fe不能参与产生的反应,致反应受到抑制;其二,加入清除剂可使反应受到抑制;其三,由于叠氮化物可抑制过氧化氢酶,加入叠氮化物使H2O2与Fe2+之间反应更加强。
d)由某些生理或病理作用产生的。
①嗜中性白细胞及巨噬细胞吞噬细菌时可产生。其证据是,加入甲硫基丙醛可产生C2H4。
②发炎。在发炎过程中有产生,证据是:其一,可被清除剂抑制;其二,可被过氧化氢酶与SOD抑制;其三,加入H2O2可增强发炎作用。这些证据表明的产生可能通过Fenton型Harber-Weiss反应。
③O3中毒。从肺部吸入O3后,O3可以诱发产生。
④电离辐射。X射线或γ射线照射生物体可产生。
C.H2O2的产生
在生物体内产生H2O2的酶促反应是比较多的,如黄嘌呤氧化为尿酸,D-氨基酸氧化为酮酸,D-葡萄糖氧化为D-葡萄糖酸内酯,胺类化合物氧化为醛类以及的歧化反应。
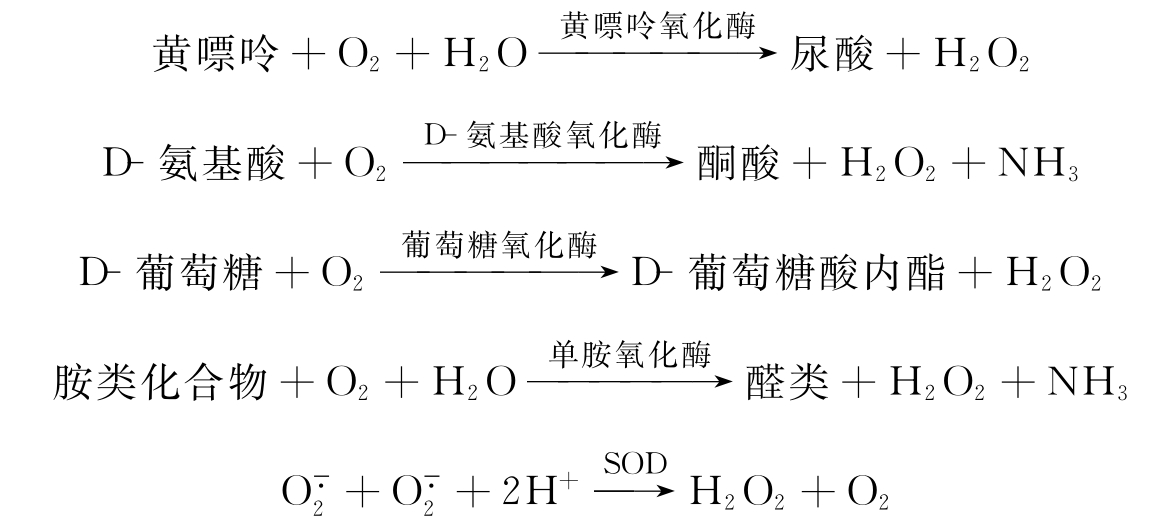
在这些酶促反应中,除了SOD催化歧化为H2O2外,其他酶均直接使O2转变为H2O2。线粒体与内质网产生的是H2O2产生的主要来源。其H2O2产量约占氧消耗量的1.7%。
D.1O2的产生
虽然有些学者认为的自动歧化反应中会产生1O2,但证据是不足的。Fenton反应产生1O2虽然有一些间接证据,然而至今尚无定论。在生物体内1O2产生的来源如下。
(1)嗜中性白细胞与巨噬细胞吞噬细菌时髓过氧化物酶的催化反应。
(2)脂类过氧化过程中RO2与RO2的相互作用。
![]()
(3)光敏反应。例如眼角膜中柱细胞含有的视黄醛就是一种光敏剂,在光敏反应中可产生1O2。
(4)某些植物和细菌中过氧化物酶的催化反应,例如:
E.脂类过氧化物的产生
RO, 与ROOH为脂类过氧化产物,但在正常生理情况下,脂类过氧化产物的含量是极低的,即使有机会发生脂类过氧化,其产物也会被转变为无害物质。
与ROOH为脂类过氧化产物,但在正常生理情况下,脂类过氧化产物的含量是极低的,即使有机会发生脂类过氧化,其产物也会被转变为无害物质。
在病理情况下,外源性物质如O3、药物与毒物都可间接引发脂类过氧化。内源性物质如活性氧增多与体内清除活性氧或抗脂类过氧化的能力下降,均可使脂类过氧化发生或加强。
9.3.2.2 自由基的清除
A.酶对自由基的清除
(2)过氧化氢酶。过氧化氢酶可清除H2O2,但在细胞内主要存在于过氧化体。在后者中,酶促反应产生的H2O2可被过氧化氢酶清除,其他细胞器如线粒体产生的H2O2能透过细胞器膜,进入胞浆。其中尚未得到清除的可进入过氧化体,最终被过氧化氢酶清除。
(3)谷胱甘肽过氧化物酶、谷胱甘肽转硫酶与其他有关的酶如葡萄糖-6-磷酸脱氢酶。在线粒体及胞浆中产生的H2O2常靠谷胱甘肽过氧化物酶清除。脂类氢过氧化物可被谷胱甘肽过氧化物酶或谷胱甘肽转硫酶清除。在这两种酶作用中所产生的GSSG在谷胱甘肽还原酶作用下又可还原为GSH,继续参加清除活性氧的反应。同时由NADPH氧化的NADP+需要通过葡萄糖-6-磷酸脱氢酶的催化作用才能再还原为NADPH,以维持体内清除活性氧的能力。
B.抗氧剂
(1)维生素E。维生素E可以猝灭1O2,又能与1O2起反应,故它是1O2的有效清除剂。依靠维生素E的这一作用,生物膜可免受1O2的损害。它还可与反应,但反应是缓慢的。在脂类过氧化的过程中起到连锁反应关键作用的RO2遇到维生素E时,可转变为化学性质不活泼的ROOH,同时维生素E转变为维生素E自由基,再变成α-妊娠醌及其他产物。这样一来,脂类过氧化的链式反应可以中断,从而起到终止脂类过氧化的作用。在抗坏血酸作用下,维生素E自由基可转变为维生素E。此反应似乎发生于膜的表面,因为水溶性维生素C不能进入膜的疏水层。
(2)谷胱甘肽。在生物体内含有SH化合物的有蛋白质与非蛋白质。在含有SH基的非蛋白质中以谷胱甘肽的含量为最高(见表9-11)。因此在防护自由基损伤作用方面比其他SH基化合物显得特别重要。

表9-11 哺乳类组织某些SH基化合物的含量

谷胱甘肽生物学作用的重要性还表现在很多方面,如它为某些酶如乙二醛酶、顺丁烯二酸单酰乙酰乙酸异构酶、前列腺素过氧化物酶的辅助因子;参与甲状腺素的生物合成、胰岛素的降解以及某些药物或毒物的代谢。
现着重介绍它防护自由基或活性氧损伤机体的作用。
a)清除H2O2。
1)H2O2+→─2GSHGSSG+2H2O
2)参加叶绿体抗坏血酸、谷胱甘肽循环以清除H2O2。
注:①抗坏血酸;②脱氢抗坏血酸;③谷胱甘肽还原酶
b)修复损伤。例如使有机自由基恢复为原有机化合物
![]()
c)防护脂类过氧化物造成的损伤。

(3)抗坏血酸。抗坏血酸是电子供体,为还原剂。当它供给一个电子时就成为半脱氢抗坏血酸自由基,若再供给一个电子,则成为脱氢抗坏血酸。
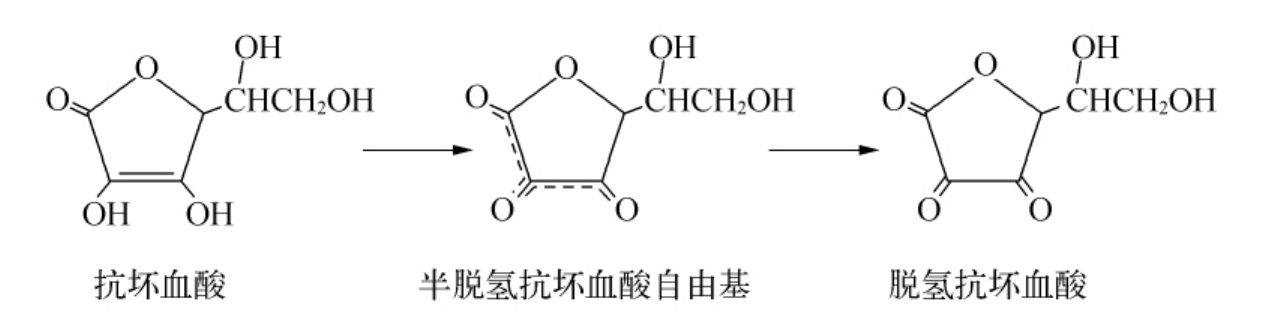
由于抗坏血酸有这种特性,在低浓度抗坏血酸中它可使Fe3+还原为Fe2+,Fe3+与H2O2反应生成,显示有害作用,但在生物体内抗坏血酸浓度比较高,它可作为自由基清除剂,如:

(4)尿酸。Ames报道,在人的血浆中含有0.12~0.45mmol/L的尿酸。它是单线态氧和的有力清除剂。有些学者还指出,尿酸可抑制脂类过氧化,而在细胞外液中SOD与过氧化氢酶都很少。Ames认为,尿酸可能和抗坏血酸一样能起到抗氧剂的作用。在人血中尚存在4.5mmol/L葡萄糖。后者也可清除。与尿酸反应产生的自由基可迅速与O2反应生成有机O2。有机O2虽为自由基也能损伤某些酶,但其反应性较差很多。
(6)血浆铜蓝蛋白。在正常成人血浆中血浆铜蓝蛋白含量为300mg/100mL,这是一种α糖蛋白。最初以为它的功能是转运Cu,但后来发现它可催化Fe2+氧化为Fe3+,而且对于许多芳香烃族胺类与酚类也有类似作用。有些学者指出,它可动员贮藏的胺类,以调节循环血液的生物活性胺如肾上腺素与羟吲哚。近来发现血浆铜蓝蛋白可能是一种抗氧剂,如能抑制由抗坏血酸或无机铁引起的脂类自氧化。它还能清除。当患发炎性疾病如风湿性关节炎、感染或怀孕时,血浆铜蓝蛋白都会增高,甚至在任何损伤情况下也有同样情况。它可能是抗御活性氧的抗炎性内生蛋白质。
血浆铜蓝蛋白清除的能力仅为SOD的1/30 000,但由于血浆中该蛋白含量较高,实际上相当于在100mL血浆中含30μg SOD,故清除的作用不可忽视。
(7)茶多酚。茶叶在中国和世界已有几千年栽种和饮用的历史。喝茶有益于健康已被广大人民所接受,喝茶可以预防一些疾病也被流行病学所证实。过去一般认为喝茶有助于健康是因为茶叶中含有维生素和一些微量元素,但是这一看法并不全面。近来研究表明,茶叶中对身体健康最有益的物质是具有很强抗氧化能力的茶多酚。茶多酚占茶叶干重的30%左右。科学研究证明,茶多酚可以有效地清除氧自由基和脂类自由基,防止脂质过氧化。茶多酚是一种多酚类物质,它包含四种单体成分,其结构如图9-10所示。
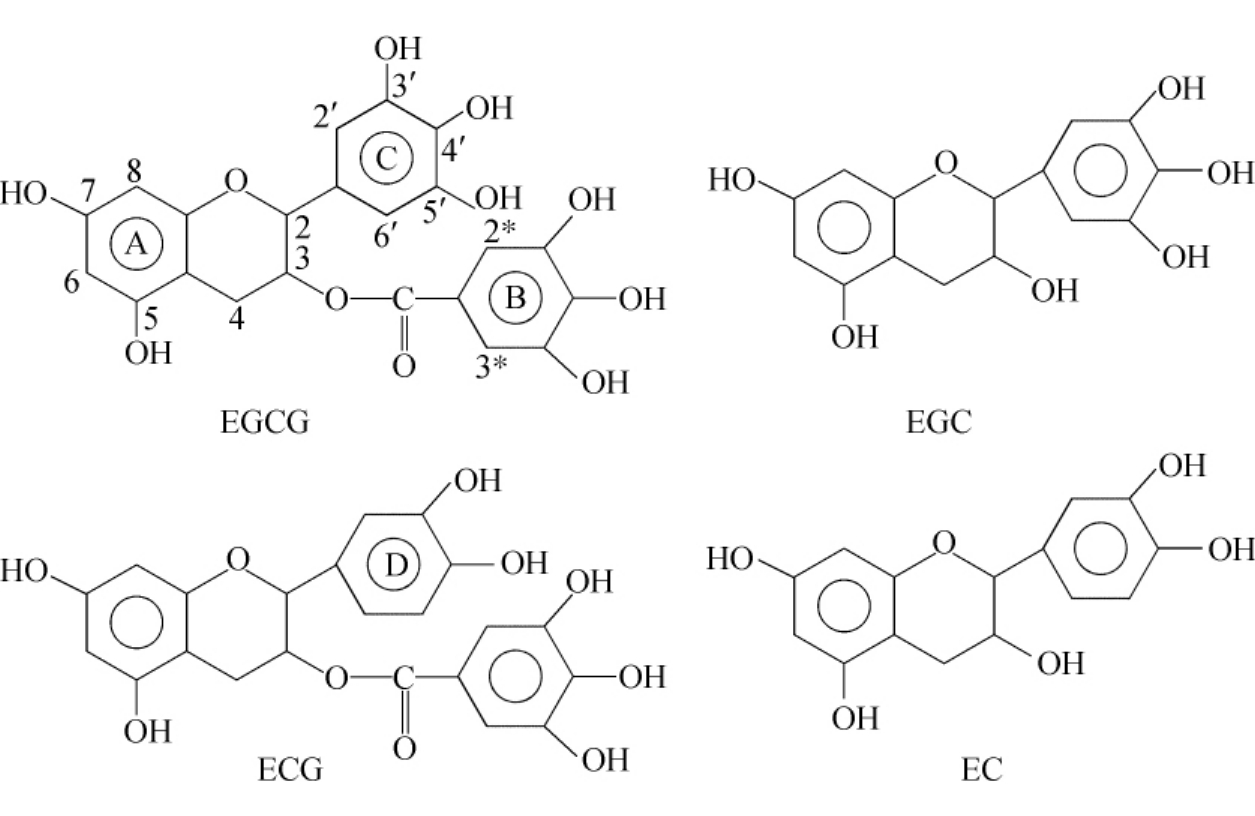
图9-10 茶多酚的四个单体的结构式
由于这四种儿茶素的结构不同,它们对氧自由基的清除效率也不一样,表9-12给出了用化学发光法和ESR技术测量的结果。
表9-12 茶多酚四种单体对超氧阴离子自由基的清除率(nmol/L)
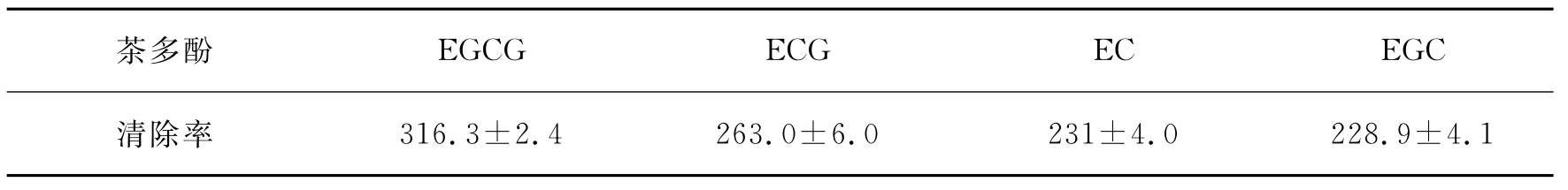
由表9-12的数据可以看出具有较多酚羟基的儿茶素具有较强的抗氧化能力。这几种儿茶素对氧自由基的清除能力按以下关系递减:EGCG>ECG>EC>EGC。
(8)银杏叶提取物(EGb)。银杏树(Ginkgo biloba)为银杏科银杏属植物,又名公孙树,为中国特有的古老植物,其叶作药用在我国已有5 000年的历史,《神农本草》就有记载,银杏具有治疗哮喘和气管炎的作用。自20世纪60年代以来,国内外学者对银杏的化学成分、药理作用及其应用作了大量的研究工作。
银杏提取物(extract of ginkgo biloba)的种类很多,提取方法不同,其效果也不同。最早注册的银杏提取物是1965年德国的商品Tebonin,后被法国研究发展注册为Tanakan(1974),现定名为EGb761,已在三十多个国家中得到应用。
EGb有清除多种自由基的作用,是一种广谱的自由基清除剂。人们的实验表明:它具有清除羟自由基、超氧阴离子自由基和一氧化氮,抑制脂质过氧化的作用。其作用强度与自由的酚羟基数目有关,并与结构中吡喃环C2~C3位间双键和B环两个邻位羟基基团有关。
C.自由基修复机理
在正常生物体内,自由基的产生与清除可维持低水平的、有利无害的平衡,但有时可能有少数自由基会损伤生物分子,使后者成为自由基。然而这些自由基又能在抗氧剂的作用下转变为原来的生物分子。这种现象称为自由基修复。通过酶反应的生物分子修复,常称为生化修复。
(2)自由基的清除。生物体具有的清除自由基的能力,可使生物分子避免受到自由基的损伤。虽然防止自由基的产生或自由基的清除都不直接关系到自由基修复,但可减少生物分子受到更多自由基的损害,有助于自由基修复效能。
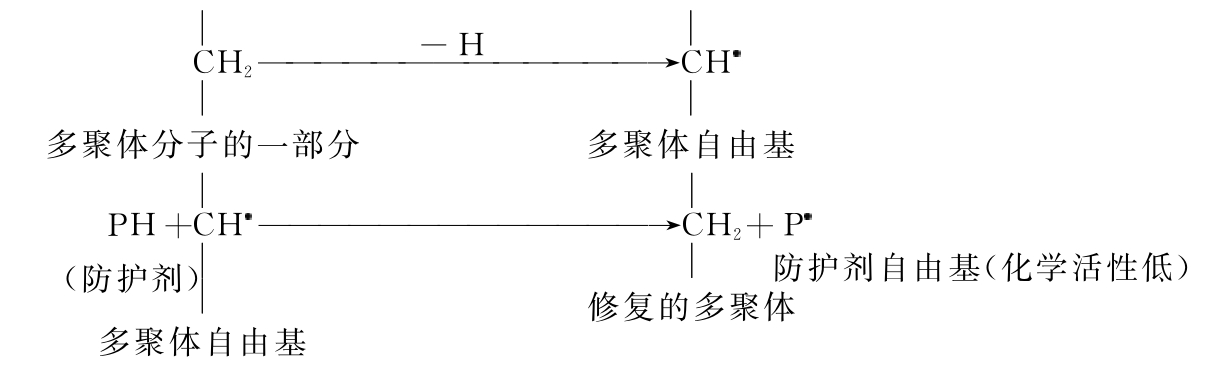
(3)自由基的修复。Alexander与Charlesby早就对多聚体的辐射防护提出了自由基修复的学术思想。他们认为巯基化合物抗辐射的机理可能就是自由基修复。其代表性反应如下所示:
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











