



3.3.1 安徽巢县马家山早三叠世生态地层剖面描述
剖面位于安徽巢湖北岸,距巢县县城约5km,巢县水泥厂采石公路旁。早三叠世地层发育完整,出露良好,化石丰富,是研究生态地层的良好场所。本剖面可划分出5个群落序列,现按剖面自新到老描述,如图3.4。
中三叠统
早三叠世末,Periclaraia和Neospathodus homeri等(早三叠世生物)在本区消失,中三叠统下部未见任何化石,其岩性以白云岩为主夹泥晶灰岩,并含石膏假晶,另有垮塌角砾。这说明中三叠世环境发生了突变,故生物群落应为突变取代。
~~~~~突变取代~~~~~
下三叠统
Periclaraia群落序列
![]()
中层至薄层生物碎屑、岩屑和砂砾屑微晶灰岩,夹有微-粉晶灰岩。生物碎屑以双壳动物和钙质海绵骨针为主,在微-粉晶灰岩中发育水平纹层。岩屑、砂砾屑和生物碎屑的存在,说明水体较浅,而水平纹层的存在,又可说明水体较平静,故推测为上部浅海上部环境,相当于Ⅲ1生境型。
![]()
本群落带有双壳动物Periclaraia circularis,P.chaoxianensis;菊石Subcolumbites;牙形石Neospathodus homeri,Cypridodella conflexa,Ellisonia sp.和裂齿鱼目及鱼碎片。典型群落分布于71,72,75和76层,尤以75层最为丰富。优势种和特征种Periclaraia circularis占绝对优势,D为95.1%,H为0.247。另一特征种P.chaoxianensis只见于71层,数量少。牙形石Neospathodus homeri最早出现于50层,一直向上延续到77层,但在77层数量极少。表生固着底栖与浮游生物之比为33∶1。双壳动物Periclaraia circularis,P.chaoxianensis的耳及足丝缺口清晰可见,均为表生固着底栖型,化石顺层面分散保存,很少有叠覆现象,每个个体保存完好。P.circularis的个体大小以8~20mm者居多,说明该群落为原地埋藏。
此群落带的围岩为钙质泥岩及灰岩,各类生物化石除牙形石外,均采自钙质泥岩,
图3.3 生态地层柱状图图例
(a)沉积岩岩石花纹图例(b)沉积构造图例
(c)各门类化石代号(d)生态类型图例
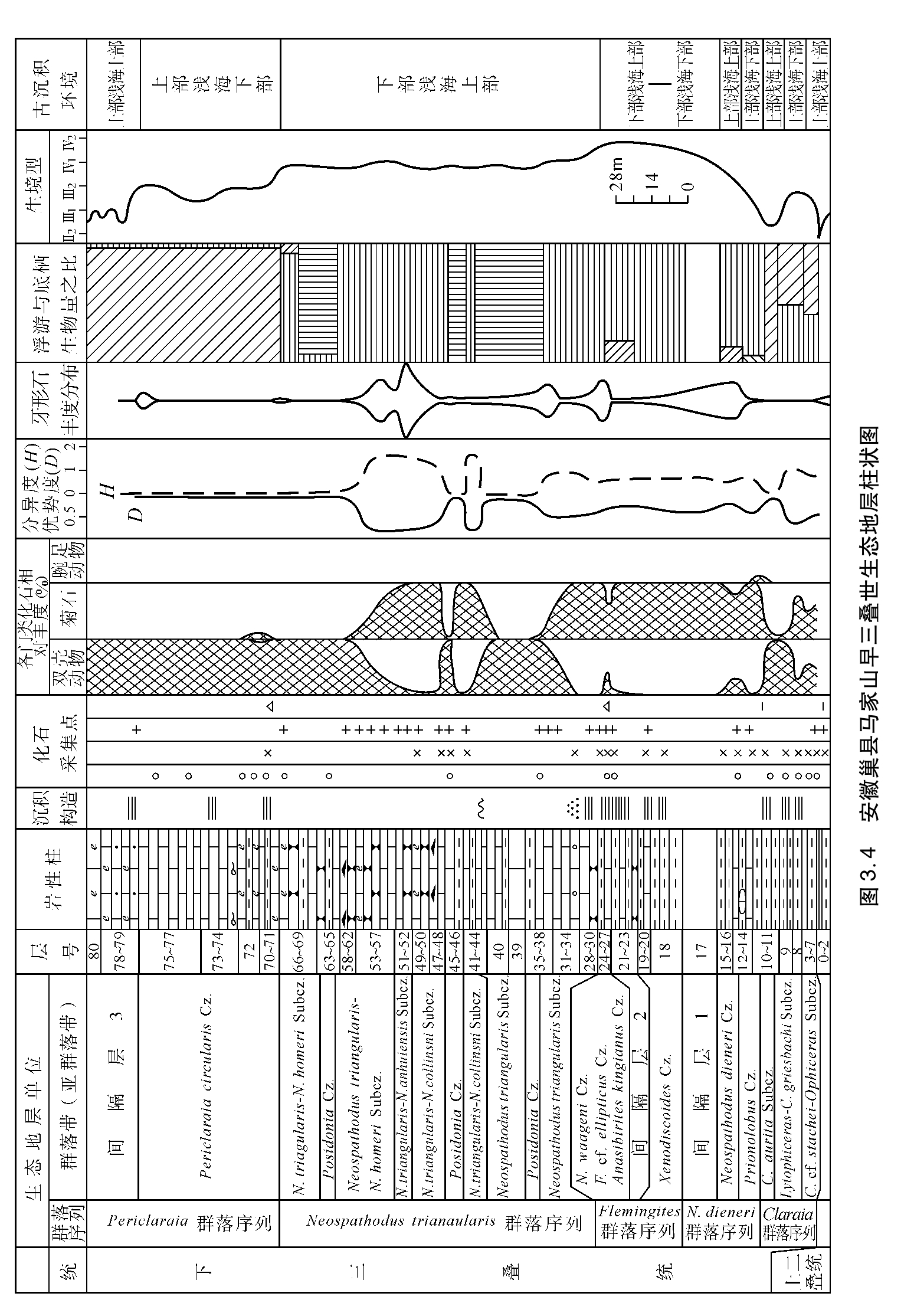
牙形石从灰岩中获得。灰岩具水平纹层,73层中含有一层10cm厚的蠕虫状灰岩,在下部的灰岩中见有介形虫碎片及少量完整的介形虫,这些都表明水体较浅。从生态分析,菊石Subcolumbites含量少,外卷型壳,壳饰粗壮,王义刚(1988)认为这类菊石生活于浅海盆地浅水区;牙形石Neospathodus homeri,Cypridodella conflexa,Ellisonia sp.等尽管数量极少,但这些分子形态纤细,属静水的产物。Clark和Hatleberg(1983)认为N.homeri主要分布于外陆架,并向深水盆地和内陆架延伸。丁梅华等(1990)认为该种主要分布于下部浅海环境。由此得出,该种的存在至少可以说明水体有一定的深度。综合上述各种因素,推测本群落带大致相当于上部浅海下部环境,属Ⅲ2生境型。
本群落序列与Neospathodus triangularis群落序列之间为突变取代,理由有两个:①N.triangularis群落序列的生物以浮游的牙形石、菊石和底游型的双壳动物占绝对优势,而Periclaraia群落序列则以表生固着底栖的双壳动物占优势;②较深水的砾屑灰岩和瘤状灰岩被较浅水的蠕虫状灰岩所取代。
~~~~~突变取代~~~~~
Neospathodus triangularis群落序列
13.Posidonia群落带(37,46,64层)厚度分别为0.60m,4.02m,2.22m
此群落带生物以双壳动物Posidonia sp.为主,D为100%,H为0,无底栖生物。在46层上部的微晶灰岩中含有菊石Pseudoceratites cf.contractus,牙形石Neospathodus triangularis,Cypridodella sp.,Ellisonia sp.,Cornudina anterodentata和双壳动物Posidonia sp.。化石保存完整。个体较小,一般为0.2~0.5cm,化石平行层面分布,都为同一面朝上密集排列。一般认为Posidonia是机会种,营底游生活,大量埋藏于贫氧、静水、水体较深的环境中。本群落带分布于扁担山组泥岩、钙质泥岩中,有时具压实现象。由于本群落带与Neospathodus triangularis群落带中几个亚群落带交替出现,故其生活环境应与N.triangularis群落带的环境相当,属Ⅳ1生境型,相当于下部浅海上部环境。
![]()
本群落带是一个牙形石群落带,根据化石分子的不同,可分为4个亚群落带,按出现的先后顺序自新到老描述如下:
12-4.Neospathodus triangu laris-N.hom eri亚群落带(53~63层,65~69层)
19.57m,16.04m
本亚群落带优势种为Neospathodus triangu laris,特征种为N.hom eri。D为28.89%,H为1.566。53~63层中共生的牙形石分子有N.collinsoni,N.bransoni,N.hungaricus,Neogondolella sp.,Cypridodella sp.,Ellisonia sp.,Cornudina anterodentata和Aduncodina unicosta;而65~69层只有Neospathodus homeri,缺乏N.triangularis。另外还有Claraia sp.。
12-3.Neospathodus triangularis-N.anhuiensis亚群落带(51~52层) 6.11m
优势种Neospathodus triangularis,特征种N.anhuiensis。D为20%,H为1.666。共生分子有N.bransoni,N.homeri,N.spathi,Cypridodella sp.,Ellisonia sp.,Cornudina anterodentata,C.oezdemirae,Aduncodina unicosta。
12-2.Neospathodus triangularis-N.collinsoni亚群落带(44~45层,47~50层)
8.28m,12.70m
优势种Neospathodus triangularis,特征种N.collinsoni。D为37.93%,H为1.597。共生的牙形石N.homeri,N.spathi,Cypridodella sp.,Ellisonia sp.;菊石Columbites parisianus,Beneckeia sp.。
12-1.Neospathodus triangularis亚群落带(28~36层,38~43层) 22.45m,21.97m
优势种和特征种均为Neospathodus triangularis。D为57.14%,H为1.418。共生的牙形石有N.spathi,Cypridodella sp.,Ellisonia sp.,Cornudina anterodentata,N.oezdemirae和菊石Columbites parisianus。
综上所述,本群落带以Neospathodus triangularis为优势分子,以N.triangularis,N.homeri,N.anhuiensis,N.collinsoni以及N.bransoni,N.spathi和N.hungaricus为特征种,遍布种有Cypridodella sp.,Ellisonia sp.等,此外还有菊石Columbites parisianus,Beneckeia sp.,菊石只见于34层、44层、47层、50层,且数量少。牙形石多产于微晶灰岩、含生物碎屑微晶灰岩和瘤状灰岩中,这些岩性在纵向上与砂砾屑灰岩呈韵律分布,砂砾屑微晶灰岩分布于本剖面的34层、40层、48层、52层、62层、66层,呈中厚层至巨厚层状,砂砾屑在岩层中一般平行层面分布,呈正粒序、逆粒序或逆—正—逆粒序,砾屑大小悬殊,一般为3~7mm,砂屑小于2mm。本区砂砾屑基本以内碎屑为主,砂砾屑成分和基质成分一致,应为风暴成因,属大陆架环境。瘤状灰岩的成因有多种解释,我们认为,此区瘤状灰岩在沉积时泥质和灰质呈一定比例,是成岩期和成岩后期压溶作用的产物,故不能用瘤状灰岩本身的特性来确定沉积环境,只能依据瘤状灰岩中所含牙形石、菊石及其纵向上风暴岩、微晶灰岩呈韵律出现来确定,因此它所形成的环境应与风暴岩的形成环境相一致。
本群落带牙形石分子的基腔比较小,细齿纤细,一般认为属平静较深水环境。Clark 和Hatleberg(1983)认为Neospathodus triangularis在Spathian早期分布于外陆架至盆地边缘,而在Spathian晚期可以从内陆架延伸到盆地边缘;N.collinsoni只分布于外陆架至盆地边缘;N.homeri可从外陆架向盆地和内陆架延伸。本群落带中的菊石Columbites以横肋粗壮的外卷型为特征,王义刚(1988)认为该属分布于浅海盆地浅水区。综合有机和无机两方面的资料,本群落带为下部浅海上部环境,相当于Ⅳ1生境型。
本群落序列与其下的Flemingites群落序列呈突变取代,突变标志有两个,一是Flemingites群落序列的Anasibirites,Flemingites cf.ellipticus和Neospathoduswaagei均先后于此群落序列末消失,取代这些生物的是Neospathodus triangularis群落序列的Columbites和Neospathodus triangularis;二是在Neospathodus triangularis群落序列中,泥岩百分含量小,灰岩多呈中、厚层状,偶夹薄层和巨厚层灰岩,见有砾屑灰岩,属下部浅海上部环境,而Flemingites群落序列中,以泥页岩为主夹瘤状灰岩和薄层泥灰岩,含黄铁矿晶体,反映一种平静、缺氧的较深水浅海盆地环境,反映水体较深。
~~~~~突变取代~~~~~
Flemingites群落序列
![]()
本群落带为牙形石群落带,由Neospathoduswaageni,N.dieneri,Cypridodella sp.和Ellisonia sp.组成。优势种和特征种均为N.waageni,遍布种N.dieneri,Cypridodella sp.和Ellisonia sp.。D为63.4%,H为1.003。在26层与牙形石共生的有菊石Pseudosageceras sp.,但数量极少。N.waageni从24层出现,一直延续到26层,在24层与菊石Anasibirites kingianus共生,在25-2层与Flemingites cf.ellipticus共生,且数量丰富。本群落带的岩性为含生物碎屑微晶灰岩、微晶至粉晶灰岩夹薄层泥岩,牙形石从含生物碎屑微晶灰岩中获得,菊石从泥岩中找到。Clark和Hatleberg(1983)认为N.waageni分布在外陆架至盆地边缘,从本区资料证实了这种结论。此群落带的围岩及生物均反映较平静的深水环境,为下部浅海下部环境,相当于Ⅳ2生境型。
![]()
该群落带的生物有菊石、双壳、牙形石和鱼类动物,主要以浮游生物为主。优势分子Flemingites cf.ellipticus,特征分子F.cf.ellipticus和Neospathoduswaageni,遍布种Claraia sp.,Entolium sp.,同时含有亚全骨鱼类。D为69.44%,H为0.92。除微体化石外,浮游生物与表生固着底栖生物之比为7∶1。菊石F.cf.ellipticus的大小个体一起保存,最小者为2mm,最大者40mm,但以中等大小为主,化石保存完整,属原地埋藏。本群落带的围岩为泥岩和灰岩,呈韵律分布。F.cf.ellipticus,Claraia sp.,Entolium sp.分布在25-1和25-2两层泥岩中,Neospathoduswaageni分布在灰岩中。Claraia sp.个体较小,最大者为6mm,小的为1~2mm,为表生固着底栖型;Entolium sp.营底游生活,可生活在各种不同深度的环境中;Flemingites为半外卷,有横纹但未成肋,该类菊石多分布在下部浅海(王义刚,1988);N.waageni数量丰富。从岩性和生物面貌分析,水体深度与11群落带相当,为下部浅海下部环境,属Ⅳ2生境型。
![]()
本群落带的生物有菊石Anasibirites kingianus,A.onio;双壳动物Claraia sp.,Entolium sp.和少量牙形石Neospathodus waageni。优势种和特征种为Anasibirites kingianus。遍布种Claraia sp.。D为83.35%,H为0.61。浮游生物与固着底栖生物之比为10.8∶1。Anasibirites kingianus个体保存完整,纹饰清晰,大小个体均有分布,以中等者居多,故该群落为原地埋藏。本群落带的岩性为泥岩与具水平纹层的微晶灰岩呈互层,菊石和双壳动物化石保存在泥岩中,牙形石在灰岩中获得。菊石Anasibirites kingianus为半内卷,纹饰较弱,未成强肋状,王义刚认为,此菊石生活在正常浅海的深水环境中。由此推测,该群落带是在水体平静的较深水环境中形成,与10,11群落带相似,均为下部浅海下部,属Ⅳ2生境型。
![]()
此间隔层的岩性为微晶灰岩、瘤状微晶灰岩与泥岩互层。瘤状灰岩是成岩期和成岩后期压溶作用的产物,它在深水、浅水中都可存在。根据微晶灰岩多为薄层状,且与泥岩呈互层,推测水体较深,为下部浅海上部至下部浅海下部环境,相当于Ⅳ1、Ⅳ2生境型。
![]()
该群落带的生物有菊石Xenodiscoides sp.,Flemingites sp.和牙形石Neospathodus pa-kistanensis。优势种Xenodiscoides sp.,特征种Xenodiscoides sp.和Neospathoduspakistanensis。D为72.22%,H为0.855。Xenodiscoides sp.分布于18层和20层中,Flemingites sp.只在20层中找到,N.pakistanensis在19层的灰岩中获得。此群落带与9,10群落带一样,均为原地埋藏。本群落带之岩性为微晶灰岩与泥岩互层。菊石保存在泥岩中,牙形石从微晶灰岩中得到。N.pakistanensis分布于外陆架至盆地边缘(Clark和Hatleberg,1983),尽管此种在本区含量少,但从整个群落面貌及围岩特征看,代表了一种较深水平静环境,推测为下部浅海上部环境,属Ⅳ1生境型。
![]()
浮土覆盖
Neospathodus dieneri群落序列
![]()
该群落带的生物有牙形石Neospathodus dieneri,N.cristagalli,Platyvillosus costatus,Cypridodella sp.,Ellisonia sp.;菊石Prionolobus sp.,Pseudosageceras sp.和双壳动物Claraia aurita。优势种Neospathodusdieneri。D为68.42%,H为1.014。特征种有N.dieneri,N.cristagalli,Platyvillosus costatus和菊石Pseudosageceras sp.,遍布种有Cypridodella sp.,Ellisonia sp.。菊石Prionolobus sp.和双壳动物Claraia aurita均为上延分子,且数量少。本群落带生物以浮游型牙形石为主。此群落带的岩性为泥岩夹微晶灰岩、含生物碎屑微晶灰岩,生物碎屑为钙质海绵骨针,少量双壳动物碎片和钙球等。灰岩中获得大量的牙形石,菊石和双壳动物保存在泥岩中。Neospathodus dieneri分布于外陆架和盆地内(Clark和Hatleberg,1983)。本剖面的N.dieneri分布于灰岩中,其上下均为泥岩,应属水体较深的环境,推测为下部浅海上部环境,相当于Ⅳ1生境型。
![]()
本群落带的生物有菊石和牙形石,优势种和特征种为Prionolobus sp.。共生分子有Prophychites sp.和牙形石Neospathodus cristagalli。D为73.68%,H为0.71。菊石保存完好,大小均有,为原地埋藏。此群落带的岩性为泥岩与泥灰岩互层,顶部为薄层微-泥晶灰岩,菊石保存在下部泥岩中,牙形石产在顶部灰岩中。Prionolobus sp.为内卷至半内卷,壳面光滑。属下部浅海上部环境,生境型为Ⅳ1。
Neospathodus dieneri群落序列与Claraia群落序列在岩性上差异不明显,但从生物群落面貌则有着显著的区别。Neospathodus dieneri群落序列以浮游生物菊石和牙形石占绝对优势,偶见表生固着底栖双壳动物;但Claraia群落序列则以Claraia各种占绝对优势,共生生物有Ophiceras,Lytophiceras和Hindeodus sp.等。因此,两个群落序列为突变取代。
~~~~~突变取代~~~~~
Claraia群落序列
3.Claraia aurita群落带
本群落带分布于巢县和江宁湖山、无锡嵩山三个地区,根据优势种的分布特点和数量的多寡,分为二个亚群落带,即Claraia aurita亚群落带和C.concentrica-C.aurita亚群落带,前者分布于巢县马家山,后者见于江宁湖山和无锡嵩山,但在无锡嵩山为异地埋藏,应称作C.concentrica-C.aurita生物组合层。
![]()
亚群落带的生物有Claraia aurita;菊石Ophiceras sp.和腕足动物Lingula。其中11-1层的化石D为100%,H为0。据野外统计,在30cm×30cm的面积中计有40个个体,最大个体长×高为45mm×40mm,一般为30mm×25mm,最小为20mm×20mm,小个体稍多一些,可能是Claraia aurita的幼年个体营浮游生活,而未能与成年个体一起保存下来;化石保存完好。此亚群落带的岩性为泥岩,Claraia aurita分布于下部,而菊石Ophiceras sp.和腕足动物Lingula sp.见于上部。Claraia aurita及Lingula均为浅水生物。在Claraia与Ophiceras之比远大于1时,一般视为上部浅海上部。为此推测该亚群落带为上部浅海上部环境,属Ⅲ1生境型。
2.Claraia griesbachi群落带
本群落带分布于巢县、江宁湖山和长兴煤山,根据优势分子分布特点和数量的不同,可分为二个亚群落带,即Lytophiceras-Claraia griesbachi亚群落带和C.wangi-C.griesbachi亚群落带。前者见于巢县马家山,后者分布于江宁湖山和长兴煤山。
![]()
此亚群落带生物有双壳动物Claraia griesbachi,C.zhenanica,C.sp.;菊石Lytophiceras
sp.,Ophiceras sp.。优势分子Lytophiceras sp.和Claraia griesbachi。D为34.52%,H为1.12。固着底栖生物与浮游生物之比为1.1∶1。Lytophiceras sp.的大小个体都有,为原地埋藏。C.griesbachi无小个体保存,可能是由于幼年个体多营浮游生活而未保存为化石所致。菊石和双壳动物化石保存完整的事实,说明它们为原地埋藏。本群落亚带的岩性为黄绿色泥岩夹薄层灰岩,化石保存在泥岩中。此亚群落带浮游生物含量比3-1亚群落带中浮游生物含量大大增加,尽管两个亚群落带中双壳动物均为表生固着底栖类型,但由于优势度和分异度的改变,推测本亚群落带的水体深度比3-1亚群落带要深,为上部浅海下部环境,相当于Ⅲ2生境型。
1.Claraia cf.stachei群落带
本群落带见于巢县和江宁湖山。根据优势分子的数量和分布特点的不同,分为二个亚群落带,即Claraia cf.stachei-Ophiceras亚群落带和Ophiceras-Claraia cf.stachei亚群落带。前者见于巢县马家山,后者分布于江宁湖山。
![]()
双壳动物Claraia cf.stachei,菊石Lytophiceras sp.,Ophiceras sp.,鱼碎片和牙形石Hindeodus sp.为本群落带的生物分子。优势种Claraia cf.stachei。D为56.36%,H为0.88。特征种为Ophiceras和Lytophiceras,但以Ophiceras为主。浮游生物与表生固着底栖生物之比为1∶1.3。岩性为钙质泥岩夹一层含生物碎屑白云岩。化石保存完整,纹饰清晰,大小均有。菊石和双壳动物化石共生,属原地埋藏。推测沉积环境为上部浅海上部环境,相当于Ⅲ1生境型。
~~~~~突变取代~~~~~
上二叠统(略)
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










