



第一节 蓝藻的生物学及生态学特征
一、蓝藻概述
蓝藻在地球上大约出现在距今35亿年前,已知蓝藻约2000种,中国已有记录的约900种。蓝藻分布十分广泛,遍及世界各地,但大多数(约75%)来自淡水。有些蓝藻可生活在60~85 ℃的温泉中;有些种类和菌、苔藓、蕨类和裸子植物共生;有些还可穿入钙质岩石或介壳中(如穿钙藻类)或土壤深层中(如土壤蓝藻)。
蓝藻不具有叶绿体、线粒体、高尔基体、中心体、内质网和液泡等细胞器,唯一的细胞器是核糖体。蓝藻含叶绿素a、叶黄素、胡萝卜素,还含有藻胆素等。一般来说,凡含叶绿素a和藻蓝素量较大的,细胞大多呈蓝绿色。也有少数种类含有较多的藻红素,藻体呈红色,如红海束毛藻呈红色,大量繁殖后使海水也呈红色,红海便由此而得名。蓝藻虽无叶绿体,但在电镜下可见细胞质中有很多光合膜,称为类囊体,各种光合色素附于其上,光合作用过程在此进行。蓝藻细胞壁和细菌细胞壁的化学组成类似,主要为肽聚糖,贮藏的光合产物主要为淀粉粒和蓝藻颗粒体等。细胞壁分内外两层,内层是纤维素,少数是果胶质和半纤维素。外层的胶质衣鞘以果胶质为主,或有少量纤维素。内壁可继续向外分泌胶质。有些种类的胶鞘很致密;有些种类的胶鞘很易水化,相邻细胞的胶鞘可互相融合。
蓝藻有球状、丝状、螺旋状等多种形式,有些蓝藻细胞分裂后子细胞埋在胶化的母细胞壁内而成为群体,如若反复分裂,群体中的细胞可以很多,较大的群体可以破裂成小群体。有些蓝藻细胞由于附着生活,有了基部和顶部的极性分化,丝状体是由于细胞分裂按同一个分裂面反复分裂、子细胞相接而形成的。有些丝状体上的细胞都一样,有些丝状体上有与固氮相关的异形胞分化(图2-1),有的丝状体有伪枝或真分枝,有的丝状体的顶部细胞逐渐细化成为毛体,也叫有极性的分化。丝状体也可以连成群体,包在公共的胶质衣鞘中,这是多细胞个体组成的群体。
蓝藻的繁殖方式有两类,一类为营养繁殖,包括细胞直接分裂(即裂殖)、群体破裂和丝状体产生藻殖段等几种方法;另一类为某些蓝藻可产生内生孢子或外生孢子等进行无性生殖。细胞分裂方式,大体可分为3类:① 三向分裂,形成群体,如微囊藻属;② 两向分裂,成平板状群体,如平列藻属;③ 单向分裂,成单细胞个体或成丝状体,如念珠藻目的种类。丝状蓝藻往往断裂成短的细胞列(5~15个细胞),称为段殖体(或称连锁体、藻殖段),可以继续分裂成新的丝状体。
蓝藻除细胞分裂增殖外,也进行无性生殖,形成内生孢子,如皮果藻属;有些形成外生孢子,如管孢藻属,有些丝状蓝藻则形成厚壁孢子,如念珠藻目。厚壁孢子有各种形状,它一般比营养细胞大,细胞壁厚,孢子内贮存大量的蓝藻体、多肽和糖原,内含物丰富,有较强的抵抗外界不良环境的能力,很多蓝藻以厚壁孢子的形式越冬。还有些蓝藻在黑暗中以其内源性可溶态磷酸盐作为磷源,利用储存的糖原作为能量合成聚磷。聚磷作为磷和能量的存储物,调节细胞中总磷的溶出率,减少不平衡渗透的破坏,同时也为合成核酸和磷酯提供所需的磷和能量。
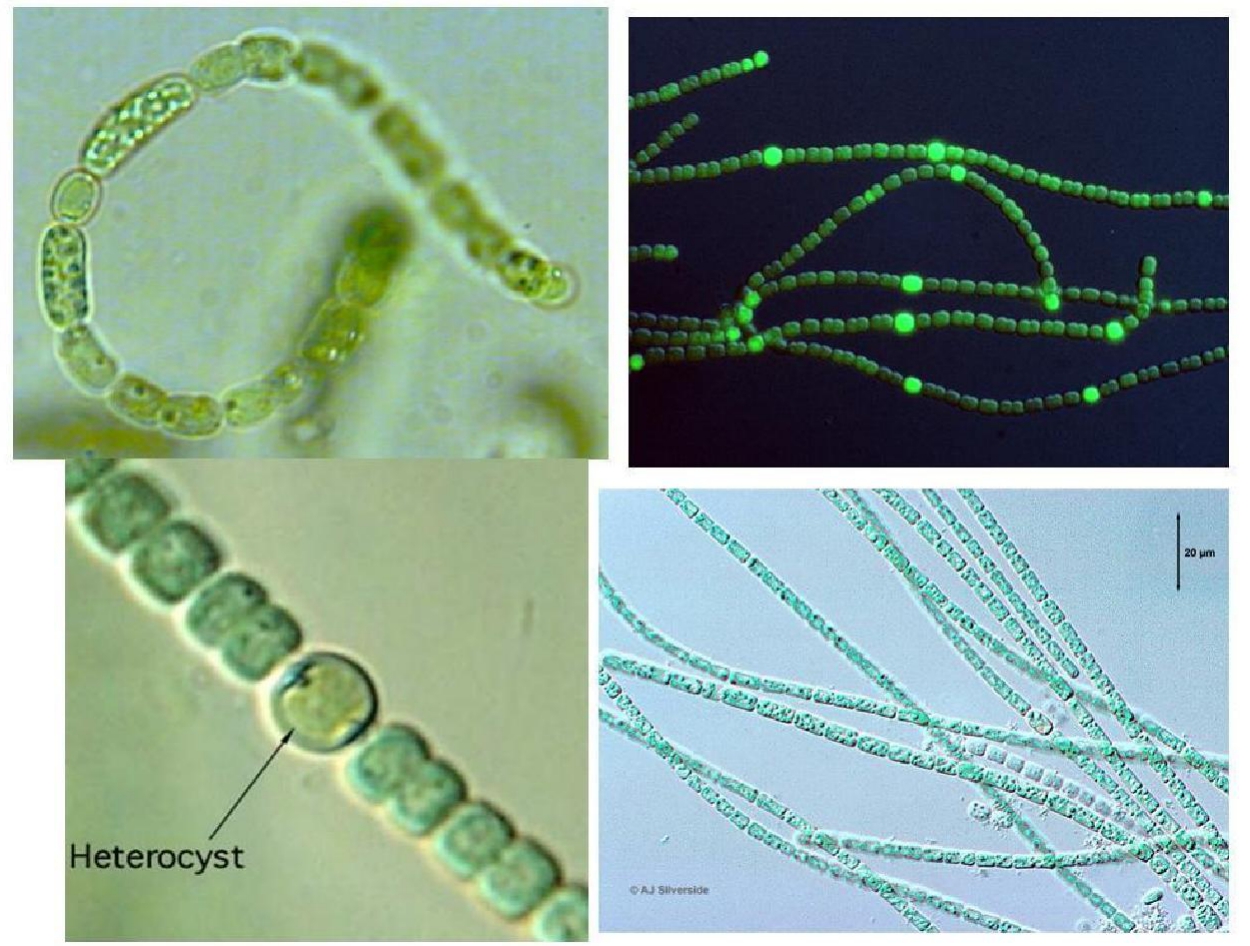
图2-1 蓝藻异型胞(Heterocyst)
二、微囊藻的生物学特征
微囊藻是太湖、巢湖、滇池蓝藻水华的优势种。群体微囊藻呈球形团块状或不规则的网状团块。细胞呈球形或长圆形,互相贴靠,一般不易见到两两成对的情况。常见的有铜绿微囊藻、水华微囊藻、具缘微囊藻和不定微囊藻等。微囊藻多生长在湖泊、池塘等有机质丰富的水体中,营浮游生活。pH以8~9.5为宜。温暖季节水温在28~32 ℃时繁殖快,生长旺盛,使水体呈灰绿色,形成水华,肉眼可见。它是一种原核生物,细胞内没有膜的细胞器,细胞外有一层组份为肽聚糖的细胞壁。其幼体为球形或长圆形的实心群体,直径3~7 μm,蓝绿色。群体胶被透明无色,一般具有伪空胞,多生长于静水中,在春季和夏季生长茂盛,常形成水华。
微囊藻属蓝藻门色球藻目,是光能自养型浮游生物。生长在水体中的微囊藻有单细胞和囊状不定形群体两种形态,群体常由几十个、几百个甚至上千个单体细胞组成,微囊藻细胞壁外被胶鞘(图2-2)。
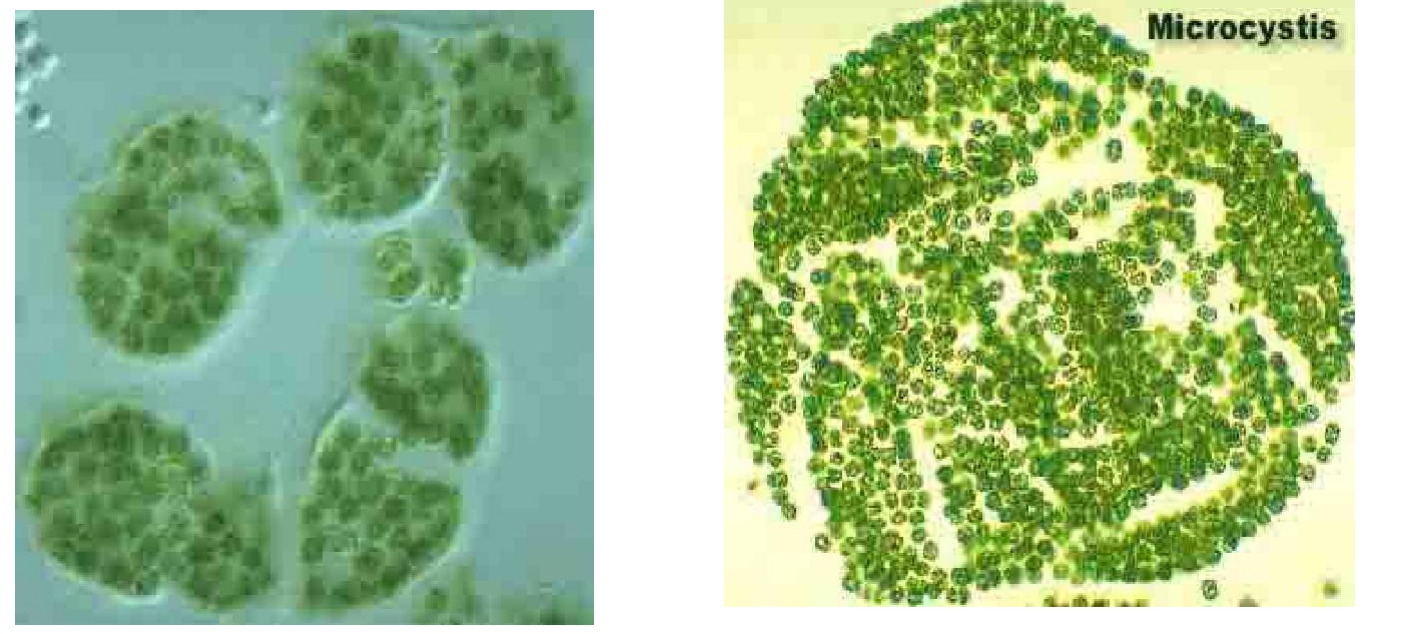
图2-2 群体微囊藻照片
1. 微囊藻群体形成的生态意义
水华暴发时微囊藻细胞聚集成群体上浮至湖面,而在室内培养条件下,野外群体会大量破裂解散,很难以大群体方式上浮。因此,形成群体是微囊藻水华暴发的必要过程,自然界存在驱动微囊藻群体形成的关键因子。综合国内外相关文献,以下阐述微囊藻群体的形成机制及其影响因素,分析群体化对水华暴发的重要意义。
微囊藻细胞在生长过程中会向胞外分泌糖类、蛋白质、脂类等有机物,一部分分泌物会在胞外积累包裹形成胶鞘(Mucilaginous sheath),这为微囊藻细胞聚集成群体提供了可能。国内的研究者通过光谱分析和色谱分析发现胶鞘主要是酸性杂多糖类物质,微囊藻分泌的胞外多糖少部分溶解到水中,大部分与藻细胞、细菌、碎屑等黏附在一起形成有机聚集体(Organic aggregates)。虽然微囊藻体内蛋白质高达49.7%。但是胶鞘使其生物可利用性受到极大的限制。我国的四大家鱼均不能消化这层胶鞘。很多微囊藻细胞内有伪空胞结构,可调节浮力进行垂直移动,具有一定的昼夜节律性和季节节律性。
对于微囊藻群体的形成过程,有研究者提出两种途径:一种是分裂形成的子细胞与母细胞未分离,导致微囊藻群体以指数方式扩大;另一种是已经存在的单细胞相互聚集形成群体。杨州等认为微囊藻群体细胞数和形态不规则,更可能是业已存在的单细胞相互黏附聚集形成的,而规则性的群体更可能是细胞分裂而未分离所致,但无论哪种方式形成群体都与胶鞘的黏附和包裹作用密切相关。Zhang等发现微囊藻群体在解散后,光合色素、蛋白、胞外多糖等含量明显降低,Yang等在浮游动物诱导实验中发现微囊藻群体形成过程伴随着胞外多糖含量增加、胶鞘增厚的现象。因此,胶鞘直接影响微囊藻表型可塑性(phenotypic plasticity),但是微囊藻胶鞘厚度的内在调节机制还需要深入研究。
2. 微囊藻群体形成的影响因素
(1)温度、光照、营养盐对微囊藻群体形成的影响
温度、光照、营养盐对微囊藻群体形成影响的报道很少,这里引用其他藻类群体形成的研究作为补充说明。Lazaroff发现在营养充足的黑暗条件下,藓类念珠藻(Nostoc muscorum)以单细胞存在,而将其转移到弱光环境则可以观察到丝状群体的形成。在低温和弱光下棕囊藻以群体形态存在,而在暖温和高光照下则表现为单细胞形态。Veldhuis等发现普氏棕囊藻(Phaeocystis pouchetii)仅仅在磷浓度低于1 μmol/L的条件下大量形成群体。Lancelot等研究发现在氮、磷、硅限制条件下棕囊藻能够大量形成群体,并且氮限制是主导因子。
通过以上研究可以看出低温、弱光、低营养盐的不利条件下藻类更容易形成群体,本研究小组通过稀释培养基,降低生长速度的方式也常常能诱导出微囊藻群体,形成群体可能是藻类适应不利环境的一种方式。一方面,群体形成后细胞间隙可以富集更多的营养元素。Grossart发现蓝藻群体内细胞间隙的磷浓度可以达到水体磷浓度的1000倍以上。耿兰芳等通过电感耦合等离子光谱(ICP)发现微囊藻胞外多糖中富含金属离子,这有利于微囊藻群体在弱碱性的富营养化水体中富集低浓度的钙、铁、镁等金属离子。另一方面,群体内部的环境相对稳定,有利于微囊藻细胞以休眠方式渡过恶劣环境。
(2)水动力过程对微囊藻群体形成的影响。
水动力过程影响微囊藻生长和生物量的空间分布,已经有研究者探明蓝藻生长与水流的阈值关系,不同强度水动力扰动的影响方式有差异。高强度的扰动不利于微囊藻生长,微囊藻群体解散,水华难以上浮,微囊藻大多以悬浮状态分布,竞争优势大打折扣。中等强度的扰动可以使微囊藻做定向运动,在湖泊中表现为水平漂移。低强度的扰动有利于气体和营养交换,增加微囊藻细胞的接触黏附机会,有利于微囊藻细胞聚集和上浮。Logan等采用浆式搅拌机和滚筒模拟实验发现微粒在合适的水动力条件下可以形成微聚体,但是提高扰动强度后微聚体分散,这与微囊藻群体的聚散过程类似。
可见,水动力过程对微囊藻群体形成乃至水华暴发有明显影响,但是在湖泊中水动力过程与风速风向、湖岸地形、水位和流量等密切相关。白晓华等的野外观测发现2.0~2.8 m/s的风速对席状、条带状水华的推动作用最明显,重度水华易于漂移到下风口的封闭湖湾堆积成灾,而此时,颗粒状微囊藻群体仅仅是随风生流缓慢起伏,这为微囊藻相互接触黏附提供了可能,当风速超过3 m/s后水华斑块迅速分散,表层水华明显减弱。
(3)浮游动物对微囊藻群体形成的影响
国内外已经有多位研究者开展了枝角类、桡足类、鞭毛虫、轮虫等浮游动物对单细胞微囊藻形成群体的诱导实验,研究者采用了微囊藻和浮游动物混合培养、半透膜隔离培养、添加浮游动物滤液等实验方式,获得了一些重要的结果。
一方面,不同种类的浮游动物对微囊藻群体的诱导效应有差异,Yang等通过浮游动物和铜绿微囊藻的混合培养实验发现桡足类、枝角类和萼花臂尾轮虫(Brachionus calyciflorus)的处理中都没有发现微囊藻群体,而在棕鞭毛轮虫的处理中发现了微囊藻群体。另一方面,同一种浮游动物对不同种微囊藻的诱导能力有差异,杨桂军等研究了角突网纹溞(Ceriodaphnia cornuta)对铜绿微囊藻(Microcystis aeruginosa)、惠氏微囊藻(Microcystis wesenbergii)、水华微囊藻(Microcystis flos-aquae)的群体诱导效果,结果只在水华微囊藻处理中发现了群体。可见浮游动物对微囊藻群体形成具有针对性,或许与专一性的信息化学物质(inforchemical)有关。Lampert等通过实验证实信息化学物质的存在,并称之为利他素(Kairomone)。Lürling推测这种信息物质可能是藻类的组成物质,经过浮游动物消化后被激活,又反作用于该种藻类诱导形成群体。
但是,浮游动物对藻细胞的消化能力需要引起重视,是否存在未消化的藻细胞团排泄后萌发成群体的可能?这需要深入研究。
(4)群体微囊藻与附生菌的相互作用
群体微囊藻上附生了大量的细菌,这些细菌与微囊藻互利共生,组成复杂的微型生态系统。一方面,群体微囊藻为附生菌提供了栖身场所和有机营养来源;另一方面,附生菌分解有机物,为微囊藻提供可利用的营养物和生长因子。尤其在水华暴发期,附生菌对有机物的分解转化有利于高生物量的微囊藻持续生长,分解衰亡藻体实现物质循环利用。邹迪等研究发现水华附生假单胞菌(Pseudomonas spp.)的吸磷能力约为铜绿微囊藻的3倍,附生菌可以将微囊藻不易直接吸收的磷酸钙和卵磷脂转化为可溶性磷酸盐,使微囊藻在有限的磷浓度中获得源源不断的供应。在微囊藻胶鞘中还存在可以降解胞外多糖的附生菌,蔡元锋等通过PCR-DGGE和测序分析发现这些附生菌主要为鞘氨醇单胞菌属(Sphingomonas)、褐螺菌属(Phaeospirillum)、假单胞菌属(Pseudomonas)、红环菌科(Rhodocyclaceae)、分支杆菌属(Mycobacterium)等种类,它们的丰度和多样性与微囊藻生物量密切相关,这在湖泊有机碳循环中具有重要意义。微囊藻的胞外多糖含量和胶鞘厚度在自身合成和附生菌的降解间获得平衡,附生菌的降解作用是否可以刺激微囊藻分泌更多的多糖并且加速群体的形成和扩大,还需要实验进一步探明。
3. 形成群体对水华暴发的影响
Reynolds和Walsby指出水华形成有3个必要条件:
① 已存在较多的生物量;
② 充足的伪空胞提供足够的浮力;
③ 水面平静。而形成群体是微囊藻积累生物量上浮的前提条件。
一方面群体化能够使微囊藻抵御浮游动物捕食维持较高的生物量,另一方面,微囊藻群体比单细胞能获得更快的上浮和下沉速率,确立竞争优势。
(1)防御浮游动物捕食
通过长期的自然选择,除了快速生长、分泌毒素以外,形成群体是微囊藻应对浮游动物捕食的有效防御策略。Burns通过实验室培养统计分析得到捕食者与被捕食者间的体型大小关系:y=22x+4.87,其中y是浮游动物可捕食颗粒的最大直径(μm),x是浮游动物的体长(mm)。由此经验公式可知,体长1 mm的浮游动物只能捕食直径小于26.87 μm的颗粒。以此推导十个细胞以上的微囊藻群体即可有效防御小型浮游动物捕食。
(2)提高垂直迁移能力
微囊藻可以通过细胞内伪空胞数量以及糖类、蛋白质等组分变化调节浮力,需要注意的是群体大小也直接影响垂向迁移速率,下面对群体大小与迁移速率的定量关系进行分析。在斯托克斯公式v=[2 (ρ-ρ0) r2/9η]•g中,v为颗粒的迁移速率,ρ和ρ0分别为颗粒与介质的密度,r为颗粒半径,η为介质的黏度系数,g为重力加速度,水华颗粒的迁移速率与颗粒半径的平方成正比。因此,微囊藻群体半径增大10倍,其迁移速度即可增大100倍,微囊藻形成群体后比单细胞藻类更快地垂直迁移。一方面,在弱光条件下微囊藻群体可以加速上浮获取有限的光能;另一方面,在强光条件下微囊藻群体加速下沉逃避光损伤,快速的垂直迁移可以更迅速地占据最佳的生态位(niche)。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












