



第二节 生物多样性
一、生物多样性
世界上目前已经发现的生物种类大约有200万种,但每年还有许多新物种被发现,估计生物的总数可达2000万种以上。其中已命名的有141.3万种,这些形形色色的生物物种就构成了生物物种的多样性。
生物多样性(Biological Diversity)是指一定范围内,多种多样活的有机体(动物、植物、微生物等)有规律地结合所构成的稳定的生态综合体。这种多样性包括物种多样性、物种的遗传与变异的多样性及生态系统的多样性。其中,物种的多样性是生物多样性的关键,它既体现了生物之间及环境之间的复杂关系,又体现了生物资源的丰富性。
我国是生物多样性最丰富的国家之一,其生物多样性居世界第8位,居北半球首位。我国的生物多样性不仅表现在物种极其丰富,生态系统类型多样,而且特有性程度高。我国特有种超过一万种,如银杏、银杉、水杉、金钱松、珙桐、鹅掌楸都属中国特有的珍稀孑遗植物;珍稀动物如大熊猫、金丝猴、褐马鸡、扬子鳄、白唇鹿等。
我们赖以生存的这个世界的重要特征和基础是生物的多样性。生物多样性和人类有着非常密切的关系。可以这样说:没有生物多样性就没有我们人类。总体来说,生物多样性是改良土壤、涵养水源、调节气候、净化环境的基础和保障,同时也是全人类食物、水和健康的保障。
生物多样性是地球生命的基础。它的重要的社会经济伦理和文化价值无时不在宗教、艺术、文学、兴趣爱好以及社会各界对生物多样性保护的理解与支持等方面反映出来。
生物多样性具有很高的价值,它不仅可以为工业提供原料,如胶、油脂、芳香油、纤维等,还可以为人类提供各种特殊的基因,如耐寒抗病基因,使培育动植物新品种成为可能。许多野生动植物还是珍贵的药材,为治疗疑难病症提供了可能。
据统计,生物资源对工农业、医药业的贡献每年价值达数十亿美元,它们在维持气候、保护水源与土壤、维护正常的生态学过程中,对整个人类作出的贡献更加巨大。
但是,随着经济的发展,由于自然资源(包括生物资源)和能源过度消耗,生态环境日益恶化,生态系统遭到破坏,生物多样性日渐丧失,保护生物多样性已是当务之急。
据有关资料指出,生物多样性毁灭所带来的直接损失在每年2万亿到5万亿美元之间。这比目前席卷全球的金融危机的损失还要大得多,而且这个损失每年都会发生。另外这个估计数字可能还显得保守,因为有些价值难以用金钱衡量。
破坏着生物多样性的主要因素有全球气候变化(如臭氧损耗和气候变暖),生境丧失和破碎化,外来入侵物种,对食物、能源和其他自然资源的不断增加的需求导致滥捕乱猎和掠夺式的过度利用等。
联合国大会宣布2010年为生物多样性国际年。这一年适逢生物多样性公约各缔约方和各国国家元首和政府首脑在2002年约翰内斯堡可持续发展世界首脑会议上通过的2010年生物多样性目标实现时期。
设立国际生物多样性年的目的是提高公众对生物多样性的重要性以及生物多样性丧失后果的认识,并对公众和其他行为者参与执行《生物多样性公约》起到极大促进作用。国际生物多样性年还庆祝了在实现到2010年大幅减少生物多样性丧失的目标方面取得的成功,其目的是获得国际社会对加强《公约》执行的承诺。
《生物多样性公约》(Convention on Biological Diversity)是一项保护地球生物资源的国际性公约,于1992年6月1日由联合国环境规划署发起、在内罗毕举行的政府间谈判委员会第七次会议上通过,1992年6月5日,由签约国在巴西里约热内卢举行的联合国环境与发展大会上签署。公约于1993年12月29日正式生效。联合国《生物多样性公约》缔约国大会是全球履行该公约的最高决策机构,一切有关履行《生物多样性公约》的重大决定都要经过缔约国大会的通过。常设秘书处设在加拿大的蒙特利尔。
该公约具有法律约束力,旨在保护濒临灭绝的植物和动物,最大限度地保护地球上的多种多样的生物资源,以造福于当代和子孙后代。公约规定,发达国家将以赠送或转让的方式向发展中国家提供新的补充资金,以补偿它们为保护生物资源而日益增加的费用;应以更实惠的方式向发展中国家转让技术,从而为保护世界上的生物资源提供便利;签约国应为本国境内的植物和野生动物编目造册,制定计划保护濒危的动植物;建立金融机构以帮助发展中国家实施清点和保护动植物的计划;使用另一个国家自然资源的国家要与那个国家分享研究成果、盈利和技术等。
截止到2010年10月,该公约的缔约方有193个。中国于1992年6月11日签署该公约,1992年11月7日批准,1993年1月5日交存加入书。
2010年10月,为期两周的《生物多样性公约》第十次缔约方会议在日本中部城市名古屋举行。会议通过了规定遗传资源利益分配的《获取与惠益分享名古屋议定书》,同时提出2010年至2020年保护生物多样性的数值目标。
一个基因可能关系到一种生物的兴衰,一个物种可能影响一个国家的经济命脉,一个生态系统可能改变一个地区的面貌。全球生物多样性的迅速丧失,不仅意味着我们正失去大量以后可资利用的资源,而且更重要的是,那将最终导致我们人类自己,也像其他生物一样,从这个星球上消失。从这个意义上说,保护生物多样性就是保护人类自己。
二、物种与生物的分界
地球上的生命是丰富多彩的,仅苔癣植物就有13000种之多,且大自然中每一样生命都是独特的,不可替代的。
世界上共有5,000,000~30,000,000种生物,已发现和命名的物种约有1,500,000种,其中植物260,000种,昆虫700,000种,脊椎动物500,000种。
(一)物种的概念
物种即种(Species)。不同专业的生物学家对物种的概念有不同的理解。分类学是依据表型特征识别和区分生物的基本单位。现代遗传学则把物种定义为:物种是一个具有共同基因库、与其他类群有生殖隔离的群体。生态学家则认为,物种是生态系统中的功能单位,不同物种占有不同的生态位。如果两个物种以相似的方式利用同一有限的资源和能源,它们必定会发生竞争和相互排斥,其中必定有一个获得相对的胜利。如果一个物种的种内发生变异,占据了多个生态位,那么从生态学的角度看,就意味着新种的生成。
我国生物学、昆虫学家陈世骧(1905~1988)先生认为:种是由种群所组成的生殖单元,在自然界占有一定的生境,在系谱上代表一定的分支。这个定义包括种的四个标准:种群组成、生殖隔离、生境地位和系谱分支,是一个广泛接受的较为完善的定义。
(二)种的命名与分类
国际上目前采用的种和亚种的命名方法是瑞典植物学家林奈在他的《自然系统》(Systema Naturae,1786年)中制定的生物学名双命名法(binomial nomenclature),即属名加种名的方法。生物学名是用拉丁文书写的,属名在前,种名在后。属名是单数名词,第一个字母需大写;种名是形容词,是限制属名的,故小写。一般还会在种名之后还应加上定名者的姓氏或其缩写。例如狼的学名应是Canis lupus Linne;人的学名:Homo sapiens,L。对于亚种一般采用三名法(trinomial nomenclature),即在种名之后再加上一个亚种名,如尖音库蚊淡色亚种(淡色库蚊)为Culex pipieus pallens Coquillet(1898)。
根据生物之间相同、相异的程度和亲缘关系的远近,使用不同等级特征,将生物逐级分类。在自然分类系统中,分类学家将生物划分为自高而低的7个阶元,它们的顺序是:界(kingdom)、门(phylum)、纲(class)、目(order)、科(family)、属(genus)和种(species)。上述7个阶元是最基本的,有时,为了更精确地表达种的分类地位,还可加入另外一些阶元。如必要时还可以在某一等级之前增设一个“超级”(super-)或在之后增加一个“亚级”(sub-),如总目、亚目、总纲、亚纲和超界、超科等。
(三)生物的分界
古希腊Aristotle首次把生物分为动物和植物两大界。林奈也把生物分为两界。两界系统比较简便,并不能反映生物界的复杂性和进化关系。但由于当时的条件限制,只能根据肉眼所能观察到的能否运动对生物进行分界。随着显微镜的发现和广泛使用后,许多单细胞生物兼有动物和植物的特性,这种中间类型的生物是进化的证据,却是分类的难题。故而,德国生物学家海克尔(E.Haeckel)于1886年提出三界学说:植物界(Plantae)、动物界(Animalia)和原生生物界(Protista)。后来的电子显微镜技术的发展,揭示了细菌、蓝藻细胞的细微结构,并发现与其他生物有显著的不同,于是提出原核生物和真核生物的概念。1969年魏特克(Whittaker R.H.)提出了五界分系统。他首先根据核膜结构有无,将生物分为原核生物和真核生物两大类。原核生物为一界。真核生物根据细胞多少进一步划分,由单细胞或多细胞组成的某些生物归入原生生物界(Kingdom Protista)。余下的多细胞真核生物又根据它们的营养类型分为植物界(Kingdom Plantae),光合自养;真菌界(Kingdom Fungi),腐生异养;动物界(Kingdom Animalia),异养。五界系统虽然能反映出生物间的亲缘关系和进化历程,但仍不够完善。
Cavalier Smith T于1989年提出了八界系统,将原核生物分为古细菌界(如嗜热细菌和嗜盐细菌等)、真细菌界,真核生物分为古真核生物界、原生动物界、藻界、植物界、真菌界和动物界六个真核生物界(图8-4)。很多学者认为这一分界系统比较合理,且可把八个界归纳分属于原核生物和真核生物两个超界。

图8-4 八界系统学说
随着分子生物学和生物化学等学科技术的发展,在分类学研究中,常用不同生物的同源蛋白质分子(如细胞色素c等)一级结构进行比较,然后根据这些分子中氨基酸差异状况再建立起生物进化系谱。但是由于这些蛋白质分子往往只存在于较高等生物中,而且实验操作也较麻烦。因此从20世纪70年代以来多用保守的核糖体小亚单位(small subunit rRNA)即ssr-RNA(原核为16S,真核为18S)的序列进行比较,并将所得到的数据再用计算机分析,由此再建立起分子系统树。然而,研究过程中发现许多生活在极端环境中的古细菌,在生理、生化及分子机制方面与真细菌(如E. coli)之间却存在着巨大的差异,为此许多学者对把生物分为原核和真核两大类的合理性提出了疑问。
从20世纪70年代中期开始沃尔斯(C.R.Woese)和福克斯(G.E.Fox)在分析了古细菌ssr-RNA序列之后,并结合古细菌的一些表型特征:如细胞壁不含胞壁酸(muramic acid);质膜中富含四醚酯;rRNA的TΨc环没有胸苷;RNA聚合酶亚基特殊等,认为古细菌应当从原核生物中独立出来。并于1980年提出了生物进化系的三主干学说。在三主干中,除了真核生物主群外,原本视为一个整体的原核生物被分为古细菌主群(Archaebacteria)和真细菌主群(Eubacetria)。这个研究结果和根据表型比较而建立的系统大部分是相符的,现在已被多数人所接受。
现在常用的生物分界系统是三主干六界的分界学说。三主干是真核生物、真细菌和古细菌。古细菌主群为一界,是三主群中最小的,大约有数十种到数百种。真细菌主群也算一界,它包括古细菌以外的所有的原核生物。真核生物主群最庞大,包括原生生物、真菌、植物和动物四界。
三、微生物及其多样性
微生物(microorganism,microbe)是一类个体微小、结构简单,肉眼看不见或者看不清楚的微小生物的统称。这些微小生物根据有无细胞结构可以分为无细胞结构微生物和细胞微生物两大类,无细胞结构微生物包括病毒和亚病毒,细胞微生物包括原核微生物和真核微生物。微生物分布广泛,种类众多,在生物六界分类系统中,微生物包括四界(图8-5)。
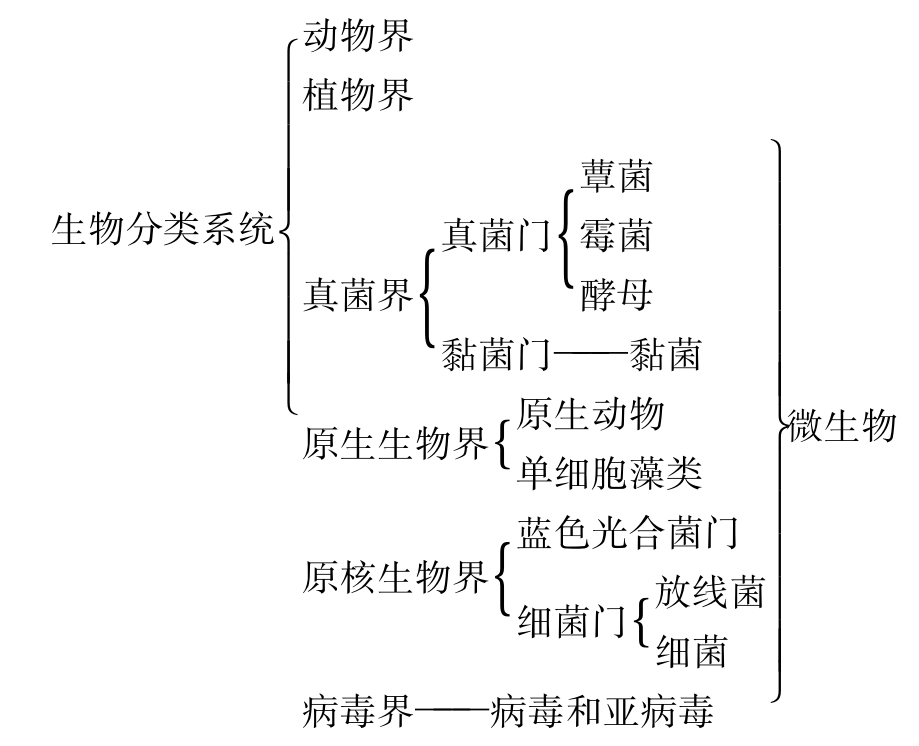
图8-5 生物六界分类系统
1.原核微生物
原核微生物是指一大类细胞核无核膜包裹,只有称作核区的裸露DNA的原始的单细胞生物,包括细菌(也有称真细菌)和古菌(也有称古细菌)两大群。真细菌的细胞膜含有由酯键连接的脂类,细胞壁中含特有肽聚糖(无壁的支原体除外)。细菌、放线菌、蓝细菌、支原体、立克次氏体和衣原体等都属于真细菌。古细菌的细胞膜含有不可皂化的脂类,细胞壁为蛋白质或假肽聚糖,但有类似于肽聚糖的杂多糖。产甲烷菌、嗜热或超嗜热菌、嗜热嗜酸还原硫酸盐菌、极端嗜盐菌、无壁热原体等属于古细菌。
(1)细菌
细菌的形态极其简单,主要有球状、杆状和螺旋状三大类,还有少量其他形状,如丝状、三角形、方形和圆盘形等。细菌大小一般以微米(μm)为单位,多数细菌细胞的直径约为0.5μm,长度约为0.5~5μm,有的细菌与最大的病毒粒子相近,显微镜下仅勉强可见,而有的细菌长度可达几百微米,肉眼可见。
细菌的结构包括基本结构和特殊结构。基本结构包括细胞壁、细胞膜、细胞质、核质体、核糖体、异染粒等,为多数原核细胞所共有;特殊结构如鞭毛、菌毛、芽孢、糖被(包括荚膜和粘液层)等,特殊结构为部分细菌或某些细菌在特殊环境下才有。
根据细菌细胞壁的化学组成成分的差别,通过革兰氏染色法染色,可将细菌明显分成两类,一类是革兰氏阳性细菌(G+细菌);另一类是革兰氏阴性细菌(G-细菌)。革兰氏阳性细菌的细胞壁厚度大,化学组分简单,一般含有90%肽聚糖和10%磷壁酸,而革兰氏阴性细菌细胞壁厚度较革兰氏阳性细菌薄,层次多,成分较复杂,肽聚糖层很薄,故机械强度较革兰氏阳性细菌弱。在革兰氏阴性细菌的肽聚糖层外部有一层主要为脂多糖、磷脂和若干外膜蛋白组成的外膜结构,脂多糖由类脂A、核心多糖和O-特异侧链三部分组成,其中类脂A是革兰氏阴性细菌致病物质。
细菌的繁殖方式较简单,无性繁殖,主要为裂殖,只有少数种类进行芽殖方式。
(2)放线菌
放线菌是一类呈菌丝状生长、陆生性强、主要以孢子繁殖的革兰氏阳性细菌。放线菌与人类的关系极为密切。绝大多数属有益菌,至今已报道的近万种抗生素中,近70%为放线菌所产生,其中链霉菌属(Streptomyces)又占放线菌中的首位(占放线菌产生的抗生素中的90%以上)。一些放线菌具有固氮和分解复杂有机物的能力,因此它们在自然界物质循环和提高土壤肥力等方面有着重要的作用。
放线菌菌体是由分枝状菌丝组成,菌丝(hypha)大多无隔膜,是单细胞微生物。菌丝粗细约为1μm。它没有明显结构的细胞核。其菌丝由于形态和功能的不同,分为基内菌丝、气生菌丝和孢子丝三种(图8-6)。
放线菌以无性方式繁殖,主要是形成孢子繁殖,也可通过菌丝断片繁殖。放线菌生长到一定阶段,一部分气生菌丝分化为孢子丝,孢子丝成熟便形成许多孢子。另外有些放线菌可在菌丝上形成孢子囊,在孢囊内形成孢囊孢子,孢囊成熟后,释放出大量孢囊孢子。
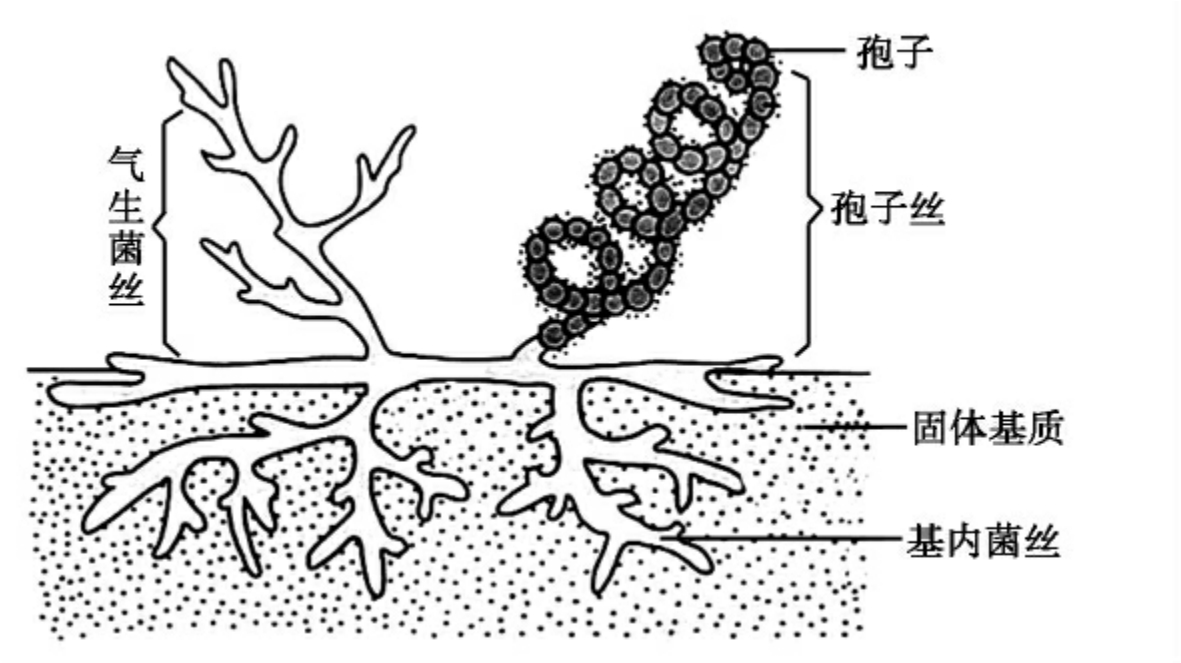
图8-6 放线菌的形态结构图
(3)蓝细菌
蓝细菌(cyanobacteria)也称蓝藻或蓝绿藻(blu-greenalgae),是一类含有叶绿素a、以水为供氢体和电子供体,通过光合作用将光能转化为化学能,并同化二氧化碳为有机物的光合细菌。蓝细菌可能是地球上第一个产氧的光合生物。蓝细菌曾作为藻类的一群,现在已知它们的细胞核为原核,所以归属于原核微生物。
蓝细菌的基本形态有球状、杆状和长丝状等(图8-7)。蓝细菌的最小直径为0.5~1μm,最大直径可达60μm。蓝细菌没有鞭毛,但能借助于黏液在固体基质表面滑行。蓝细菌的运动有趋光性和趋化性。
蓝细菌的主要繁殖方式是裂殖。有些丝状蓝细菌还能通过静息孢子(akinete)度过不良环境进行繁殖。
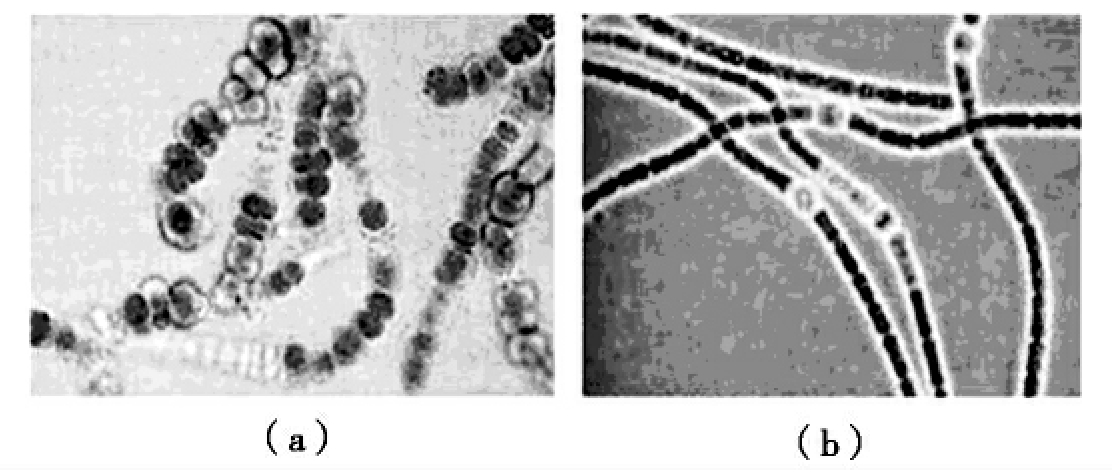

图8-7 几种蓝细菌的形态
(a)费氏蓝细菌属;(b)鱼腥蓝细菌属;(c)粘杆蓝细菌属;(d)皮果蓝细菌属;(e)颤蓝细菌属。
(4)支原体、立克次氏体和衣原体
支原体(Mycoplasma)、立克次氏体(Rickettsia)和衣原体(Chlamydia)是三类革兰氏阴性细菌,代谢能力差,主要营细胞内寄生的小型原核生物。其大小和特性均介于细菌与病毒之间。
支原体是一类无细胞壁、介于独立生活和细胞内寄生生活之间的最小型原核生物。其突出的特点是无细胞壁,形态易变,对渗透压敏感,对抑制细胞壁合成的抗生素不敏感,对能抑制蛋白质生物合成的抗生素(如四环素、红霉素等)和破坏含甾体的细胞膜结构的抗生素(两性霉素、制霉菌素等)都很敏感。支原体分布广泛,腐生和寄生都有,有一些是动植物的病原菌。肺炎支原体(Mycoplasma pneumoniae)则是人的病原菌,能引起非典型肺炎,与由病毒引起的非典型肺炎不同。
立克次氏体是一类只能寄生在真核细胞内的革兰氏阴性原核微生物,它有细胞壁,不能进行独立生活,其细胞较大,无滤过性,合成能力较强,且不形成包涵体。立克次氏体可使人患斑疹伤寒、恙虫热或Q热等传染病,病原往往由节肢动物(如虱、蚤、螨等)为媒介。立克次氏体的致病机制主要是在宿主血液中大量繁殖,并产生内毒素,使人致病。
衣原体是一类在真核细胞中营专性寄生的小型革兰氏阴性原核生物。衣原体的特点是有细胞构造,在细胞内同时含有DNA和RNA两种遗传物质,有细胞壁(但缺肽聚糖),革兰氏阴性,缺乏产生能量的酶系统,须严格细胞内寄生,以二分裂方式繁殖,对抑制细菌的抗生素和药物敏感。能引起的疾病主要衣原体有:引起鹦鹉热等人兽共患病的鹦鹉热衣原体(C.psittaci)、引起人沙眼的沙眼衣原体(C. trachomatis)和引起肺炎的肺炎衣原体(C.pneumoniae)。
(5)古细菌
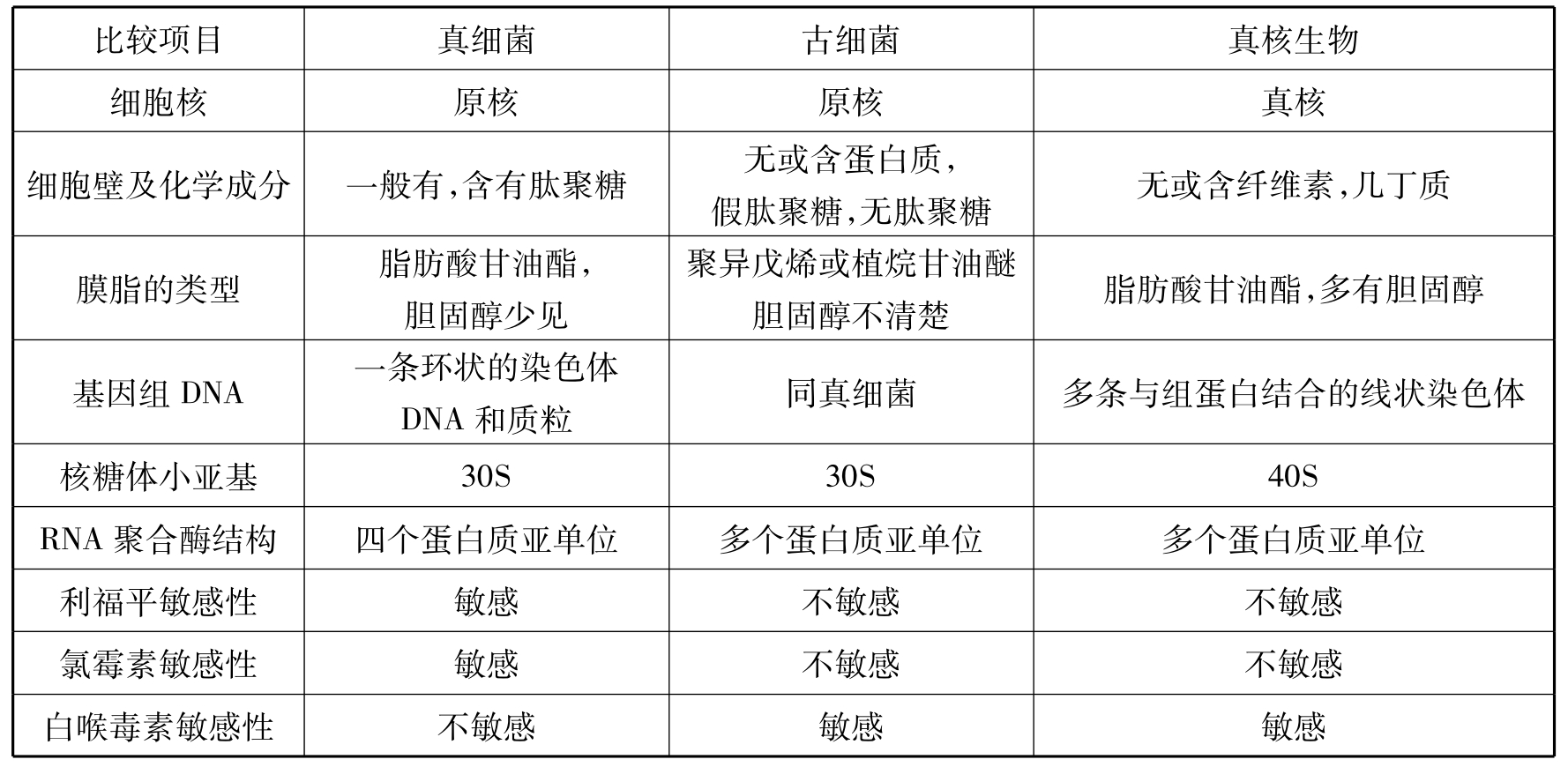
古细菌(archaeobacteria)(又称古生菌、古菌)是一类很特殊的细菌。在细胞形态、细胞核类型、染色体形状等方面与真细菌非常相似。但是在蛋白质合成、DNA复制、核糖体对氯霉素不敏感等方面,古细菌更像真核生物。此外,还具有既不同于原核细胞也不同于真核细胞的特征,如:细胞膜中的脂类是不可皂化的;细胞壁不含肽聚糖,有的以蛋白质为主,有的含杂多糖,有的类似于肽聚糖,但都不含胞壁酸、D型氨基酸和二氨基庚二酸。真细菌、古细菌和真核生物的主要性状对比如表8-1所示。
古细菌多数生活在极端的生态环境中,包括极端厌氧的产甲烷菌、极端嗜盐菌、低酸和高温环境中生活的嗜热嗜酸菌以及无细胞壁的热原体属。
表8-1 真细菌、古细菌和真核生物的对比
2.真核微生物
真核微生物是一大类细胞核具有核膜,能进行有丝分裂,细胞质中存在线粒体等多种细胞器的微生物。真核微生物的主要类群包括真菌(fungi)、单细胞藻类(algae)、原生动物(protozoa)、黏菌(myxomycota)等,真核微生物种类约占微生物总数的95%以上。
(1)真菌
真菌(包括单细胞酵母菌类、丝状霉菌类和大型子实体的蕈菌类)是真核微生物中最重要的一类群体,其种类达10万多种。相互间的形态差异极大,有菌体小至显微镜下才能看见的单细胞酵母菌,也有大至肉眼可见的分化程度较高的灵芝等蕈菌的子实体。生殖方式以无性或有性进行,产生各类无性或有性孢子为主。生活史简单或复杂并存。生活习性多样,腐生、寄生、共生,兼性或专性等都具有。
酵母菌(yeast)是非分类学术语,泛指能发酵糖类的各种单细胞真菌,形状因种而异,通常呈球形、卵圆形、圆柱形或香肠形。酵母菌在自然界分布很广,主要分布于偏酸性含糖环境中,如水果、蔬菜和果园土壤中,以及油田、炼油厂附近的土层里(烃类物质利用)。酵母菌与人类的关系极为密切,几千年来人们利用酵母制作出许多营养丰富、美味可口的食品和饮料,如酒类生产、面点制作等。酵母菌的繁殖分有性繁殖和无性繁殖两种方式(图8-8),以无性繁殖为主,无性繁殖以芽殖为主,也进行裂殖,产生相关无性孢子,酵母的有性繁殖是以形成子囊(ascus)和子囊孢子(ascospore)的方式进行,两个性别不同的酵母细胞,进行融合,经质配、核配、减数分裂后,产生子囊,子囊内形成4个或8个子核,每一个子核和其周围的原生质形成孢子。含有孢子的细胞称为子囊,子囊内的孢子称为子囊孢子,子囊孢子萌发长成新的酵母细胞。

图8-8 酵母的繁殖方式
霉菌(mold)是那些菌丝体发达,而又不产生大型肉质子实体的丝状真菌(filamentous fungi)的俗称。霉菌在固体营养基质上形成绒毛状、棉絮状或蜘蛛网形丝状菌丝体。霉菌喜潮湿气候,多为分解有机物的腐生营养型。在地球生态中,霉菌作为分解者起着极端重要的作用。霉菌与人类生活密切相关:在工业上,很多精细化工及其中间产品是由霉菌生产出来的,如柠檬酸、L-乳酸等有机酸和淀粉酶、蛋白酶等酶制剂;在医药工业领域,多种抗生素(青霉素,头孢霉素,灰黄霉素等)是利用霉菌进行发酵生产的;在食品制造领域,很多食品如酱油的酿造和干酪的制作等,都是在雷菌参与下生产加工出来的;很多霉菌也会导致大量工农业产品的霉变,有些霉菌是动物和植物的病原菌;少数霉菌甚至会产生毒性很强的真菌毒素,如黄曲霉产生的黄曲霉毒素(aflatoxin)毒害肝脏,易引发肝癌等。霉菌主要依靠产生形形色色的无性或有性孢子进行繁殖。

图8-9 蘑菇的子实体和担孢子
蕈菌(mushroom)又称伞菌,通常指那些能形成大型肉质子实体的真菌,包括大多数担子菌类和极少数的子囊菌类。典型的蕈菌,如蘑菇,其子实体是由顶部的菌盖(包括表皮、菌肉和菌褶)、中部的菌柄(常有菌柄和菌托)和基部的菌丝体三部分组成(图8-9)。蕈菌广泛分布于地球各处,主要生长在含有有机物丰富的森林土壤中或树干上,许多蕈菌的子实体营养丰富、味美可以食用,目前已利用的食用菌约有400种,已人工栽培的约有50多种,如常见的双孢蘑菇、木耳、平菇等,还有许多种可供药用,如灵芝、云芝、猴头等。蕈菌的有性繁殖是通过担子上着生出外生孢子-担孢子来进行,担孢子成熟后,以弹射的方式从担子上散发出来,经空气传播,单倍体的担孢子在适宜的基质上萌发长出菌丝,但不能形成子实体,单倍体的菌丝经融合形成双核菌丝,生长到一定时期形成小的类似菌蕾的结构,这个结构是蕈菌子实体的雏形。菌蕾在地下可以保留很长时间,直到适宜的条件到来,通常在暴雨后,菌蕾吸收大量水分而膨胀,形成子实体。这个过程需要的时间很短,几个小时或一天,许多子实体可同时形成。
(2)真核藻类
真核藻类是指除苔藓植物和维管束植物以外,含有叶绿素,可进行光合作用,并伴随释放氧气的一大类真核生物。其形态多种多样,有单细胞或多细胞藻类。多细胞藻类多呈丝状。藻类的细胞核为真核,具有真核细胞的一般特征。不同的光合色素使藻类呈现不同的颜色。藻类主要为水生生物,在自然界水生生态系统中,藻类是重要的初级生产者。在特定条件下,藻体异常增殖可造成水体污染,给人类生产与生活带来危害。藻类的繁殖方式有营养繁殖、无性繁殖和有性繁殖。
(3)粘菌
粘菌(Slimemolds,又称粘质霉菌)是非光合营养的真核微生物。其特点是不含叶绿素,吞噬方式摄取食物,能产孢子和子实体,因此它们是介于真菌和原生动物之间的一种生物。生活在腐烂的枯枝落叶、木头和土壤中,它们的食物以细菌为主的微生物,靠吞噬方式摄食。
(4)原生动物
原生动物是一类无色、无细胞壁、能进行运动的单细胞真核微生物。属于原生生物界,是最原始、最低等、结构最简单的单细胞动物,广泛分布在海水、淡水和潮湿的土壤中。
原生动物主要是以吞噬作用捕食其他生物或有机颗粒来获得营养,也通过胞饮作用获得溶液中的大分子物质。有些原生动物,如鞭毛纲种类具有色素体,可以进行植物性营养(光合作用),还有很多种类是进行腐生性营养。
原生动物的繁殖可分为无性繁殖和有性繁殖,主要进行无性繁殖,类似于原核生物的二分裂方式。
根据运动方式的不同以及在自然界中的分布,原生动物可分为鞭毛纲、肉足纲、纤毛虫类、孢子虫纲、眼虫类(也被认为是藻类)五个类群。各类群的特征如表8-2所示。
表8-2 原生动物主要类群的特征

3.病毒
病毒为非细胞生物,无细胞结构。病毒的特点非常突出:(1)病毒无细胞结构,个体极其微小;(2)只含一种核酸,病毒的遗传物质只含有DNA或RNA中的一种;(3)专性寄生,病毒一般不含有酶系或酶系极不完全,所以不能独立地进行代谢活动,只能在特定的活的宿主细胞里,利用宿主细胞内的酶系进行复制和增殖;(4)对抗生素不敏感,对干扰素敏感。
病毒基本构成单位是病毒粒(virion)。病毒颗粒由核酸和蛋白质外壳(即衣壳(capsid))所组成。核酸位于内部,外部为蛋白质包裹,衣壳由许多亚基构成,每一亚基称衣壳粒(capsomere)。病毒的蛋白质衣壳是由衣壳粒按照一定的规律排列而成,衣壳粒亚单位有规律的排列使各病毒具有不同的形态,一般分为三类:螺旋对称、多面体对称、复合对称。
病毒是一类严格的专性活细胞内寄生生物,缺乏活细胞所具备的细胞器(如核糖体、线粒体等)以及代谢必需的酶系统和能量。繁殖所需的原料、能量和生物合成的场所均由宿主细胞提供,在病毒核酸的控制下合成病毒的核酸(DNA或RNA)与蛋白质等成分,然后在宿主细胞的细胞质或细胞核内装配为成熟的、具有感染性的病毒粒子,再以各种方式释放至细胞外,再次感染其他细胞。这种繁殖方式称为复制(replication),整个过程称为复制周期(replication cycle)。无论是动物病毒、植物病毒或微生物病毒,其繁殖过程虽不完全相同,但基本相似。概括起来可分为吸附、侵入与脱壳、复制、装配以及释放5个连续步骤。每一步骤的结果和时间长短都随病毒种类、病毒的核酸类型、培养温度及宿主细胞种类不同而异。
亚病毒包括类病毒(viroid)、卫星病毒(satellite virus)和卫星RNA(satRNA)、朊病毒(prion virino)。在亚病毒中,仅有类病毒和朊病毒能独立复制,朊病毒颗粒是不具有核酸的小分子蛋白质。卫星病毒和卫星RNA都具有核酸,但是必须依赖辅助病毒进行复制。
动、植物的很多疾病是由病毒引起的,最早发现的滤过性病毒就是能引起烟草花叶病的病原物。人和动物的病毒病更不胜枚举,人的艾滋病、天花、流感、麻疹、肝炎、脊髓灰质炎,家畜的口蹄疫、禽类的劳氏肉瘤、牛痘等等,都是由病毒引起的疾病。
干扰素(interferons,IFNs)是高等动物细胞在病毒、灭活病毒或dsRNA等诱生剂的刺激下,所诱导产生的一种具有高活性、广谱抗病毒等功能的特异糖蛋白。它的功能除了能抑制病毒在细胞中的增殖外,还具有免疫调节作用和对癌细胞的杀伤作用,因此可用于病毒病和癌症的治疗。
干扰素虽有广谱抗病毒的特性,但受宿主种属特异性的限制。如人细胞产生的干扰素只能保护人细胞免受各种病毒的感染;由鸡产生的抵抗流感病毒的干扰素,可抑制鸡身上包括流感病毒在内的多种病毒的感染,却不能用于抑制人或者其他动物的流感病毒感染。早期用于临床应用的干扰素都是通过细胞工程方法得到,从20世纪80年代初,开始采用基因工程菌大规模发酵生产各种干扰素,在治疗流行性感冒、乙型肝炎、黑色素瘤和若干癌症中具有较好的疗效。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










