



生物进化与球面沿革
古话说:少所见,多所怪。试问多所见的,就不足为怪吗?我们试想想山原河海,草木鸟兽,那一样不是可怪的东西!我们祖先,不懂得这些“怪”,所以用许多神话去解释。对于地面形势,他们说什么真火铸出来的,又说什么挠亚洪水冲出来的;对于生物,他们说什么女娲氏抟土为人,又说什么上帝七日造成的。在十七八世纪之时,虽是有康德拉布拉司说地球是从太阳里射出来的,然而究竟还没有解释球面之变迁;虽有朋奈(Bonnet)说他看见鸡蛋壳里,已经有一个完完全全的小鸡在那里啄米、饮水、抓地皮,然而究竟还没有解释人与动物之发生的关系。一直到了十九世纪科学昌明的时代,才有一个里页耳(Lyell)和达尔文(Darwin)把这一类的神话根本破除,而改用普通的天然力来解释这些怪而不怪的问题。我们且来叙述这些问题之科学的解释。
(一)生物进化
我们试到万生园中看看,有许多花卉果木羽毛虫豸,各种有各种的习惯,各种有各种的形状。但是这些光怪陆离的种,还是自有生以来,就是这么多呢?还是同出于一原呢?从前的人以为:自有生以来,就是这么多种,各种有各种的特性,这些特性,就是亚里士多德所谓“最终法式”。他想把宇宙间事事物物都分成类——在生物界即为种,——每一类有一类的最终法式。虽然每类中分子不齐,不必尽合乎这个法式,然而总有要合乎这个法式之趋向。譬如说:人类之长为五尺半,虽有超过五尺半的长子,和不够五尺半的矮子,然而大家都有要合乎五尺半之趋向;夏天之温度为九十度,虽有时热到一百度,有时只热到八十度,然而所有夏天,都有要合乎九十度的趋向。这样的理论虽是不易受人家的攻击,然而究竟不能餍满实证精神之欲望。中古以后,进化观念逐渐发展,当时大哲学家如康德笛卡儿莱柏尼都有关于进化的学说。但是这些学说不过是哲学的思辨,不是科学的证明;不过是从生物学范围以外的研究,而延及生物学,不是由生物学范围以内的研究,而构成系统的联贯。有系统的科学的生物进化之理论,实在是查里士达尔文一人苦力创造出来的。自他的《原种》(Origin of Species)出世之后,大家才知道这生物界中许多的种,都是同出于一原。生物进化之理论,到了他的手里,才有确定的意义,丰富的证明。现在我们把这个理论,叫做达尔文主义,他实在可以当之而无愧色。所谓达尔文主义,不是专指狭义的竞争,如宗教家之所指摘的,它实在包含全部的进化,换言之,即是说明种类之变迁。
各种不同的生物,既是同出于一原,然而何以自一原而变成许多种呢?这个问题,笼统说起来,是很简单的,——变而已矣。然而详细分析起来,却不是易于解决的。我们先从同的方面看起来,最易看得出的,是同类必生同类。俗话说:龙生龙,凤生凤,老鼠生儿子会打洞。这个道理,就是生物学里所叫做的遗传(heredity)。然而生物界中,如果只有遗传一个因子在那里动作,则始祖所生出的子孙,必定个个都和始祖一样,那就不成其为种了。再从异的方面看来,每个有每个的性质形状习惯之不同,无论如何相近决不能够完全是一样的,俗话说:一娘生九子,九子不像娘;这个道理,就是生物学里所叫做的分衍(variation)。然而生物界中,如果只有分衍一个因子在那里动作,则自始祖起,每代所生出的,都和父母不同,而且彼此都不相同,那么,地球上的生物,纵的横的不过是千万无数的分个,那也就不成其为种了。所以遗传分衍两个因子,必定是相辅而行,才能生出来同之间又有不同,不同之间又有同的种。然而依此说来,生物界中之种,一方面因为遗传而有同,一方面因为分衍而有不同,那么,若把生物界中现在存在的各种排列起来,应该成一个不可辨别的联续;甲与乙相似,甲乙之间,又有丙与甲更相似,甲与丙相似,甲丙之间又有丁与甲更相似,就同无限分数一般。我们试设想在生物族系树(genealogical tree以表牒叙明生物发生之系统,其形式如树者)上,若是枝桠过于繁密,——繁密到了中无空隙的地位,则成混然一片的堆体,还有什么族系之可言?然而依普通事实看来,有些不同的种,其间似乎有判若鸿沟的界限,又是什么缘故呢?对于这个困难,我们必定另外还要有一个理论来解释它。生物由分衍而来的不同的种,有适宜的,有不适宜的,适宜的生存,不适宜的淘汰。古语说:顺天者存,逆天者亡,我们若把这个天字当作天然的“天”解释,这两句话所包含的道理,就是生物学里所叫做的天择(natural selection)。我们所看见的种,都是经过天择而生存的种,还有许多已经淘汰的种,我们现在看不着了。所以我们觉得这些生存的种之间,有判若鸿沟的界限。这个理论,是生物学中最大的发明,也就是达尔文进化理论之脊椎骨。又有分限(isolation)一个因子,也是“使种之区别加大”之一个原因。例如二海之间有土峡,或者两平原之间有高山,或者两大陆之间有沙漠,则两边生物之区别甚大。这一种原因,也是可以成立此种与彼种之间之界限的。我们现在就遗传、分衍、天择、三个理论,各为叙述一番。
(1)遗传
父母的性质,既与子女的性质有同有异,然则以何种性质遗传下去呢?还是只有先天的性质——种的性质(racial character)——遗传下去呢?还是后天的性质——习得的性质(acquired character)——也可以遗传下去呢?自兰马克(Lamarck)和爱拉斯莫司达尔文(Erasmus Darwin)倡后天性质可以遗传之说,十九世纪中的生物学家,大半都没有反对的。依兰马克的理论,一个机关之发展和退缩,全凭用与不用而定。用者发达,而且将此已发展的机关,直接底遗传下去。所以就此部机关而言,是有“干父之蛊”的气象,一代比一代好。不用者退缩,而且将此已经退缩的机关,直接底遗传下去,所以就此部机关而言,是有“五世而斩”的气象,一代不如一代了。
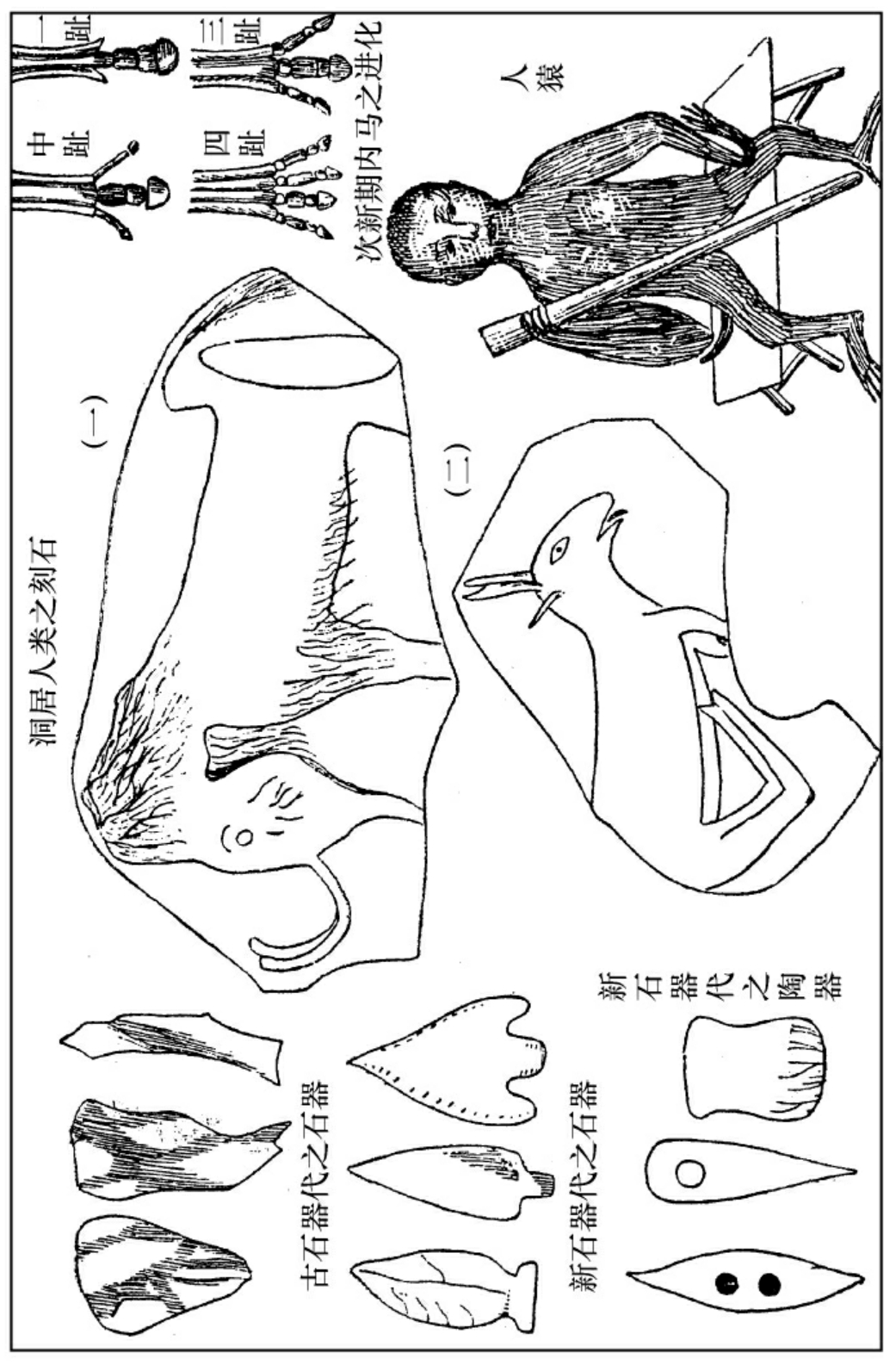
赫胥黎拿家鸭和野鸭的翅骨和腿骨之重量,互相比较,他寻觅出来家鸭的腿骨较重于野鸭的腿骨,野鸭的翅骨较重于家鸭的翅骨。他从此推论:家鸭和野鸭虽出于同一的祖宗,然而野鸭飞的时候多,家鸭走的时候多,所以野鸭的翅骨发展,腿骨退缩,家鸭的腿骨发展,翅骨退缩。起初不过是小有不同,但是这个发展和退缩之结果,是直接遗传下去的,所以经过许多代数之后,发展者愈见其发展,退缩者愈见其退缩,于是就大有不同了。他又在新生地层之中考察河獏变马之历史,他寻出来最古一层有河獏,其足有五趾;较新的一层中,有三趾马,其足只有三趾了;再较新的一层中,有主趾马,其中趾较大,左右各一趾较小;至于最新的一层中,则有我们现在所看见的马,只有中趾发展最盛而成为蹄,其左右二趾竟缩成微细的软骨。足见河獏进化之时,因为奔走甚多,所以中趾特别发展,其他各趾,逐渐退缩(中趾在奔走时最为得力)。一直到了马的阶级,中趾之发展,已达极点,然后可以有一日千里之能力了。
德康多耳(De Candolle)试验植物,使之受饿,其结果乃得较平常更繁丽的花蕊;用此受饿之植物之种子传下去,其生出之子孙,花蕊亦茂盛。淘沃(Tower)以蕃薯甲虫置之非常温度及湿度之下,因其甲壳坚硬,不起变化,但其所生出的子孙,倒有不同的颜色与花纹,而且递传下去,不复回祖宗之状态。这些事实,也似乎是辅助后天遗传之理论的。
不但筋骨之发展与退缩,可以直接遗传,即脑力之发展退缩,也可以直接遗传的。进一层说,下等的本体(instinct)可以进化成为高等的智慧(intellect)。达尔文把针鱼养在玻璃箱里,以海水注之,箱内安一玻璃墙,将此箱隔成甲乙两段,针鱼放在甲段里,他又把一些活的小鱼,放在乙段里。针鱼本是肉食的鱼,所以它们看见着小鱼,就一往直前底射激过去吞噬。但是它们这种举动,不但是没有果腹而归,而且在那玻璃墙上,碰着一个大钉子。到了碰的次数多了,它们的射激的举动,也渐渐减少了。到了一月之后,他们虽看见墙那边的小鱼,似乎是到了身边,也就望望然而去之。以前见小鱼而射激,是它们的本能,以后见小鱼而不射激,已经是智慧之萌芽了。若是遗传的代数多了,就渐变成种的习惯(racial habit)。猫狗见家禽而不咬,就是种的习惯,由遗传而得来的。垩文(Owen)带着一种猎犬去打猎,他打着了两只野鸡,但是不过受了伤,都还不曾死;这种猎犬的天性,是喜欢生擒禽兽来给它们的主人,所以那中射的禽兽,如果不是已经射死了,它决不咬死着衔回来。但是这一次,它却遇着一个困难的题目:它一次不能衔两个回来。如果它只衔一个回来,又恐怕那一个跑脱了。它徘徊了一些时候,于是它咬死一个,而把第二个活的,衔着回来。不咬死是它的本能,这一次咬死,也就是智慧之作用。兰马克派相信这样习得的智慧,也是要影响到生殖细胞而遗传下去的。
我想乘叙述后天智慧是否遗传的机会,简单讨论动物是否有智慧之动作。动物之有本能,例如蚕吐丝,蜂酿蜜,而且这种本能,是可以遗传下去的,已经成为无可辩驳的真理。但是对于智慧,却有一些意见之不同。有人以为智慧(intellect)与理性(reason),乃异字而同义。它与语言为同存的性质,是人类所独有的。动物只有本能,如上文所举的针鱼,是小鱼而射激,猎犬不咬死禽兽而衔归,都是本能的作用,即令有遇变迁的环境,而变迁其本性的事情,也不过是属于感觉方面的动作(perceptual process),不是属于概念方面的动作(conceptual process)。概念是由抽象而得来,抽象须从个体的事实,抽出普遍的概念;这是智慧的作用。概念须有一种符号来维系它,语言乃是尽这个维系的责任的。希尔(Hill)教他的狗开箱子,而以饼干为奖赏,它开一次,可得一块饼干。于是当它的主人在旁边的时候,它就不待命令而开之。以后他把一块热肉,放在箱子里,又将这只箱子放在没有人的院子里,他在一个窗子里望着;这只狗虽是闻着肉味,但是不知道立刻开箱子去取出来。开箱子而盼望主人给饼干,不过是一种感觉的作用,开箱子而自取东西,乃是概念的作用。因为要从它的经验之中,把开箱子和主人和饼干三个概念分开,而把开箱子一个概念联合到箱子里边的热肉上去,不是一只狗所能做得到的。所以我们只能说动物有知性(intelligence),不能说动物有智慧(intellect)。知性是包含感觉的动作和概念的动作二者而言,智慧是专指概念的动作的。但是感觉的动作和概念的动作,也难有确定的分界。一个高等动物改变它的习惯以应付新发生的环境之时,我们能断定它没有“用此方法得彼结果”之概念动作吗?若说概念动作,在动物进化程序上边,究竟发生在什么时候,我们无从知道,所以我们不能说动物有概念动作之本领,那也是不充分的理由。我们人类不能记忆四岁以前的历史,试问我们能够断定我们人类概念之动作,究竟发生在什么年龄吗?若说语言为概念之符号,则动物亦可吐出不同的声音,表写不同的心理状况。若说联接语言(联结字而成句,联接句而成篇),是人类所独有的,所以惟有人类有概念的动作,那也不是无可辩驳的定案;因为人类也有用简单的声音表现情感之时,动物也能了解简单联接的语言,既能了解,必能成立语言所代表之概念。我们自己听见了敲钟,就到饭厅上去吃饭,我们承认我们自己的脑子里,有一个“凡敲钟的时候都有饭吃”的普遍概念。我们看见一群鸡子听见管家太太祝祝的呼声,就围着她而待哺,我们为什么一定不承认它们脑子里有“凡被呼的时候都有食吃”的普遍概念呢?况且从行为论方面讲来,有一定的激动,都有一定的反应;我们固然不能证明概念之有,却也不能证明概念之无。人与动物都是如此,并没有什么不同,纵有不同,也不过是阶级之不同,决不是类别之不同。但是说到此处,我们要归到遗传之本题了。从动物进化到人类,从本能进化到智慧,必定有后天的因环境变迁而习得的性质遗传下去,才能一步步的提高。这是兰马克派所最注重的一层。
如果后天性质可以遗传之理论,是十分普遍的真理,那倒是很可庆贺的一桩事情。那么,我们只要拼命读一代书,我们的子孙,个个都是孔夫子;我们只要竭力学一代算学,我们的子孙,个个都是奈端。岂不是好!无如事实一方面,却不是如此如愿。我们详细考察,可以否证此理论的事实极多。例如磨擦而生的粗皮,操练而生的筋肉,都是不能直接底遗传下去的。还有一件事,是我们所最常看见的例子;就是:中国女子缠足,年代总也不算少了,然而女子却不是生来就是小脚。至于由学习得来的知能,不能遗传下去,成为良知良能,那更是不待言了。所以近二三十年来,反对此说者,日见其多,然而推源溯始,当推门德耳(Mendel)为第一人。
门德耳的基础试验,仍为现在动物学中所常引的,就是高矮豌豆之配合。以高豌豆和矮豌豆相配合,则所生的第二代豌豆,并非不高不矮,介乎二者之间,如我们的常识所预期的,但是全是高的。若将这第二代高豌豆自相配合,则所生的第三代豌豆,又不全是高的,如我们的常识所预期的,但是四分之三是高的,四分之一是矮的。若将这第三代的矮豌豆自相配合,则所生的第四代豌豆又全是矮的;足见这第三代矮豌豆,是纯矮种。但是第三代高豌豆之中,有三分之一,若自相配合,则所生的第四代豌豆,全是高的;有三分之二,若自相配合,则所生的第四代豌豆,和第二代高豌豆所生的相同;足见第三代高豌豆之中,有三分之一是纯高种,有三分之二是杂高种。今以「」为杂种之记号,列表于下:
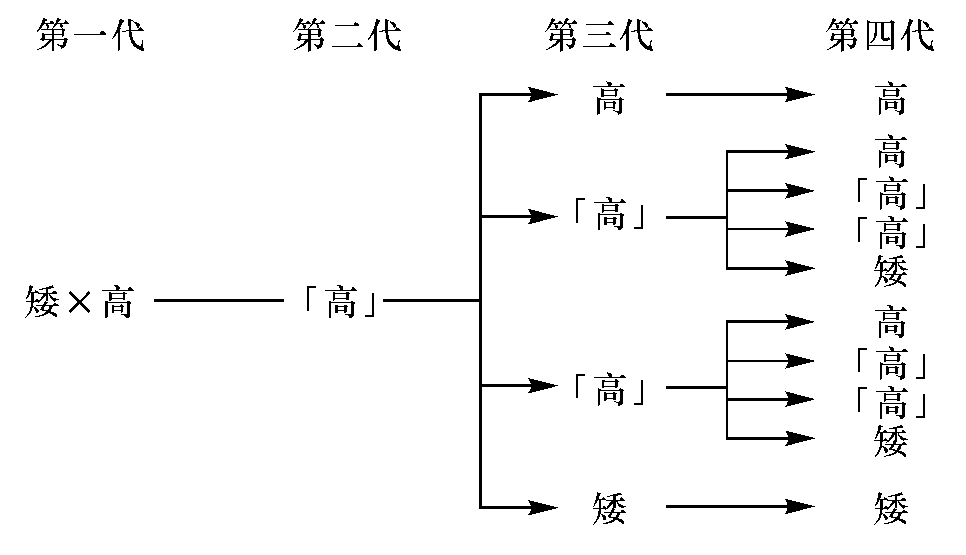
他把第二代必发见的性质(如上例中之高),叫做盛的性质(dominant character),第二代隐藏的性质,叫做衰的性质(recessive character),这盛衰二性质之共总,叫做相配的对偶(allelomorphic pair)。
以上所举的例,不过是一个古老而简单的。以后生物学家对于豌豆之黄(盛)绿(衰)圆(盛)绉(衰),兔鼠之灰(盛)黑(衰),鸡之玫瑰冠(盛)和笔架冠(衰),蜗牛之无花纹(盛)和有花纹(衰),麦之无须(盛)和有须(衰),鼠之平常性质(盛)和旋转之性质(衰),都做过同样的试验,而所得的结果,都和门德耳所考的成分相符。这个成分,就叫做门德耳成分。这些事实,不是后天性质可以遗传之理论可以解释得了的。所以门德耳另外创设一个单位性质之理论(unit character)。此理论之大概,是以一物之所有的性质,分为许多单位,如上段所说的高、矮、青、黄、圆、绉,等等。这些相配的性质,盛者发见,衰者隐藏。但是这个隐藏的单位性质,并不是消灭的,或融化的,它在后代,还要发见出来,就同化学中的原子之不可磨灭一般。有了这个理论,我们才懂得何以子女的性质,有时与父母不同,何以一族系中,有时有一种性质隔一代或两代而发见;推而论之,何以以尧为父而有丹朱,以瞽瞍为父而有舜,都可以藉此而解释了。
近代反对后天遗传之最有名的学说,就是威司蛮(Weismann)的生殖腜津永活(continuity of germplasm)之理论。这个理论,乃是近代生物学中极大的出产品。试为节述如下:
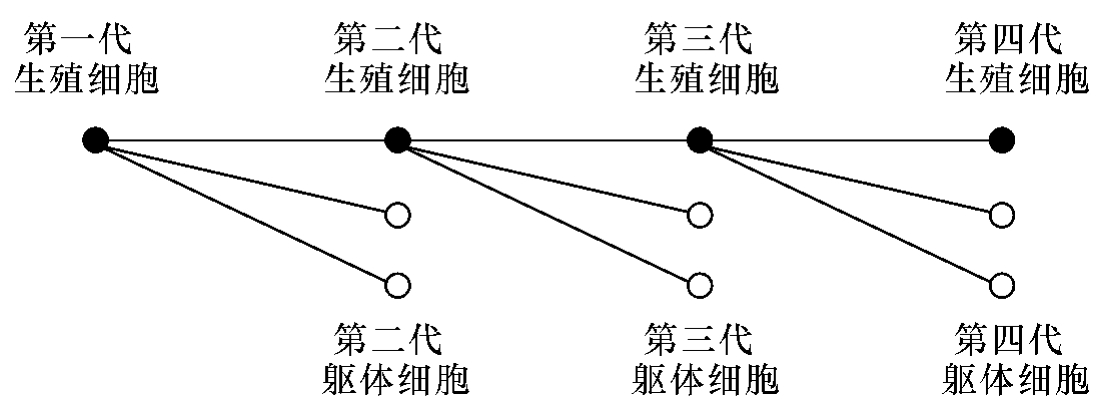
一生物之起首,乃是一个生殖细胞。此细胞乃是由一个精虫与卵结合而成,其中有细胞核,此细胞核中有生殖腜津。当一个生殖细胞由一分判为二,由二分判为四,由四分判为八……而发达为个体生物之时,留着一部分生殖腜津长育为生殖细胞,以供第二代生殖之用,而用其余的部分长育为躯体上的各机关,如手足头尾心肝肺肠之类。后一类的细胞,统称躯体细胞。到了第二代生殖之时,也是“如法炮制”。这样看来,躯体细胞,不过是生殖细胞的附属品、旁产物。人之有死,不过是躯体细胞倒塌罢了。至于生殖细胞,早已在生殖的时候,分判而成为第二代生殖细胞,储藏在子女的生殖机里边,自有生以来,未曾死过;因为一个细胞分判而成两个细胞,不能算做死。生殖细胞,永远在那里分判,所以他说:生殖细胞是长生不死的。而且若依此说,则当一个生殖细胞发达成为生物的时候,已经储留一部分腜津为第二代生殖细胞,那么,儿子乃是父亲的弟兄,女儿乃是母亲的姊妹,父母还未出世之时,子女已经分配出来,父母后天的行为,自然不能影响到子女的性质。所以他说:后天性质是不能遗传的。总之,他以为生殖细胞和躯体细胞是不相搀扰的,躯体细胞,有生有死,生殖细胞,永生不死。生殖细胞是主体,躯体细胞是旁产物,不过用作视听思想消化运动……各种器具,以供给生殖细胞之生活而已。
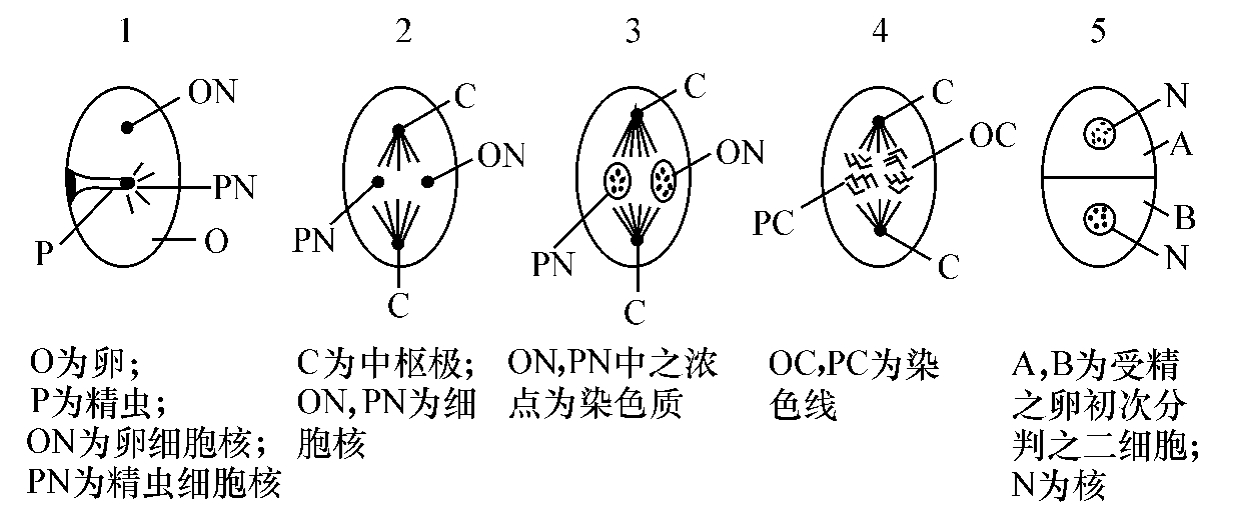
单位性质,丽附于生殖细胞之细胞核之中。细胞核当分判之时,有染色线(chromosomes)。染色线之多寡,以各种而不同,有一种高甲(虾类),其染色线有一百六十八条之多。但是:无论如何底多,总不能以一个染色线代表一个单位性质,因为一个生物之性质太多,不是染色线所能代表得了的。所以:我们还要在染色线之下层,去寻单位性质之居停,就同在原子之下层去寻电子一般。一个细胞核在生殖细胞里边,和独立的生物相同,可以藉食料而生长。它里边储藏生殖腜津,就是细胞学(cytology)中所叫做的染色质(chromatin)。(但是生殖腜津有时要包括核中之仁(nucleolus)与核边之中枢围(centrosphere)而言。)这些染色质,是由若干壹旦(idants)集合起来的,每个壹旦,是由若干壹德(ids)集合起来的。这些东西,经过一定染液浸润之后,都可以用显微镜看得出来(所以叫染色质)。这些染色质,在分判的时候,排列成为上节所说的染色线。每个壹德是一个小宇宙,把各种性质发展之可能,都包藏在里边。它的组织,是依过去的历史而规订。每个壹德,又由许多定子(determinants)集合起来。每一个定子,将来即长成一个可以独立底随环境而变衍的机关,例如手足心肝之类。这些定子之集合之形式与成分,都是由历史规订的,换一句话说,性质之遗传,都是靠着他们去担当这一份责任。每个定子,又由许多生质(biophores或progen或gammules)集合起来。这些生质,或者可以离细胞核,入细胞液而支配其行动。但是这些极小的分子,也出于闻见的范围,近于玄学的存体,无怪行为论家笑称它们为显微镜里面的上帝。
雄的生殖细胞为精虫,雌的生殖细胞为卵。当精虫入卵之时,精虫核与卵核相接近,两核中之染色质,和在细胞分判的时候一样,发生为染色线。今以鼠为例而言,精虫核与卵核所发生之染色线,其数各为十二,共有二十四。这些染色线复各自剖析,成为四十八。其时核墙消灭,而有两个中枢极(centrosomes)成于两端。此四十八条染色线,有二十四条归于此极,有二十四条归于彼极。每极所得之二十四条之中,有十二条是原于精虫核的,有十二条是原于卵核的。于是这个受精之卵,遂分判而成两个细胞。每个细胞,各得一半原于精虫核的,一半原于卵核的染色线。这是很重要的一层。因为如此,所以受精之卵分判为两个细胞之时,每个细胞核中所有的染色线之数,仍然与未分判时之染色线之数相等。而且一半是原于雄者,一半原于雌者。因为性质之遗传,是由于染色线之分配,已成确不可移之义,所以我们可以论定:凡子女所得自遗传的性质,一半是原于父的,一半是原于母的。以后细胞分判,染色线都是照样的剖析。所以无论分判若干次数,每个细胞里之染色线之数是不变的。总之,精虫核与卵核中之染色质,发生为染色线,及染色线分归两极,这两层行为,必是父母性质遗传子女之枢纽;而且父母性质之遗传,各有二分之一之机会,也似乎是显而易见的。
照单位性质之理论讲来,生物性质之单位,是有一定的,不能创造,也不能消灭。那么,后天的性质,自然不能加到原有性质里边,一道遗传下去。照腜津永活之理论讲来,生殖细胞与躯体细胞是不相搀扰的,那么,躯体细胞方面的后天行为,自然不能影响于生殖细胞,而遗传于后世。所以这两种理论,是同属于后天性质不能遗传之一派的。
但是威司蛮的理论,也不能博取全部的生物学家的信从。其持异议最盛的,是为赫特维(Hertwig)。他反对的理由是:(一)生殖腜津永活理论中之所假定的细胞核之别择的分判,——即是一部分成为躯体细胞之核,一部分成为生殖细胞之核,使原来一种细胞成为两种细胞,——并没有切实的证明。而且增倍的分判,——即一个细胞核分成两个细胞核,与原来细胞核完全相同,而且彼此完全相同,——反来有较多的实据。例如单细胞的生物——如酵母——并无躯体细胞与生殖细胞之分,它的分判总是增倍的分判。(二)低等动物——如马蝗——之重生,无论割切那一部分,都可以长成一个单位的动物,即高等动物亦有修补损伤之能力;足见不同样的细胞,可以通融使用。(三)植物可以萌生,以一枝番薯藤插于土中,即成一个独自生活的番薯,足见其躯体细胞之中,有生殖细胞之分子。(四)高等植物可以移接,梨树上可接桃子,蒿子上可以接菊花,足见不同种的细胞,都可以交相为用。(五)人可以换血,以强壮者的血液贯入患血亏的血管,可以收得与自己血液相同的效用,足见在不同个体中之细胞,——血轮——也可以代行供养之职务。(六)用人工分判海猬之卵,各部皆可长一个生物,足见此卵之各部,皆有生殖腜津。他根据这些事实,反驳威司蛮的理论。他以为一个生物,各机关(如呼吸机关消化机关)的细胞,有各自特别的性质,然而都有一种潜隐的性质,为此一种生物所共有,而此潜隐性质,即依据于可供生殖的物质。任何躯体细胞之中,都有此种可供生殖的物质,不一定要以生殖细胞为限。
所以:后天性质不能遗传之说,虽是博得多数之赞同,然而:倘若我们说绝对不能,也未免失之急切。新兰马克派极力想设法转圜,改变兰马克原来的简单方式,即威司蛮本人,晚年也承认后天的行为,可以影响生殖腜津之内容。现在讲遗传的,大约都以为后天性质,是不遗传的,但是“可以发展后天性质”之可能性,是遗传的。例如一个白人居于热带,可成棕色,然而他的儿子,决不是生来就是棕色。这就是说:他的棕色的——后天的——性质,没有直接底遗传下去,但是他的可变棕色之可能性,是遗传下去的。总之,一个性质之因子有二:(一)细胞之组织,(二)环境之刺激,二者缺一,则此性质不能发生。但是环境之刺激,有常存的,如空气、日光、食料、仇敌;有不常存的,如社会上情境之变迁。我们往往把由于常存的刺激而发生的性质,叫做先天的性质,或种的性质,把由于不常存的刺激而发生的性质,叫做后天的性质,或习得的性质。其实里边必定有一定的组织,外边必定有一定的刺激,才能发生一定的性质,无分于先天后天啊。以上所说的,都不过是把已有的性质保存下去,所说的变迁,都不过是退步的变迁,这就是说:将原有的性质减少若干而已。然而自单细胞进化而到我们人类,其中必定有进步的变迁,这就是说,有些时候,必定加入一些新性质。这个进步的变迁,由何而来,现在生物学家还没有一定的解释。倘若他们能够给予我们一定的解释,则我们对于各种生物,——包括我们自己人类而言——要加入什么性质,就加入什么性质,其有益于人类,或者比蒸汽机还要大得多。我们说及这一层,就不能不讨论分衍了。
(2)分衍
生物之性质不同,原于其细胞组织之不同,然而细胞组织,何以渐到不同的地位呢?实在要归功于分衍。我们可以把分衍分为二类:(A)寻常的分衍,或简称修改;(B)出轨的分衍,或简称剧变。
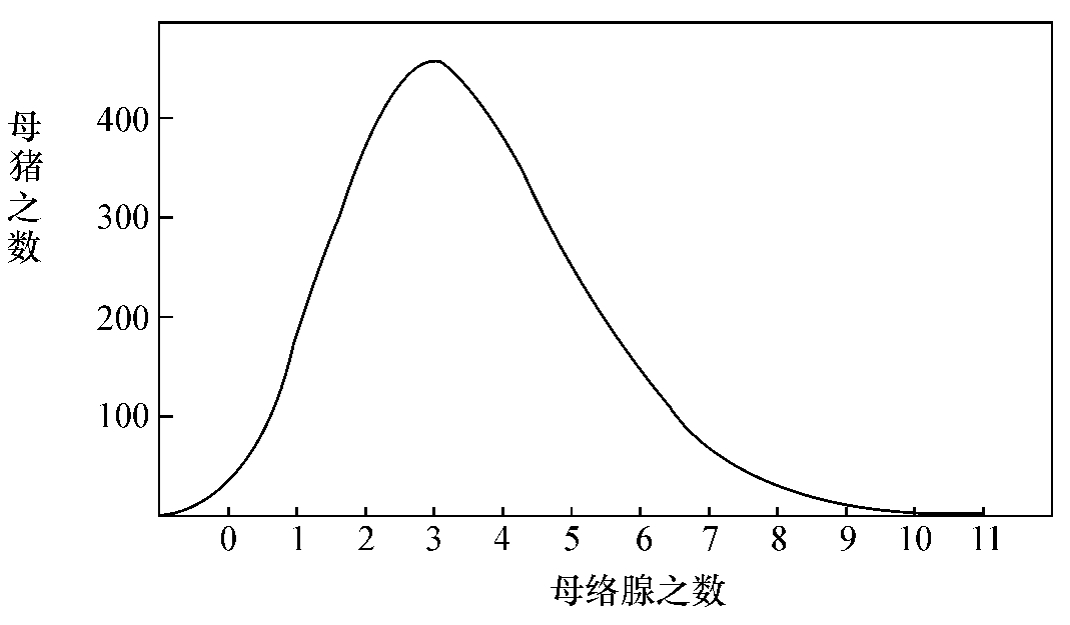
(A)寻常的分衍 我们试到一个会场,考察人之高度,最高的或有六尺多高,最低的或有四尺高,然而普通多数总在五尺半左右。若用方格纸统计之,以竖标记人数,以横标记高度,则必得一抛物线状的曲线(如图)。加耳敦(Galton)皮耳孙(Pearson)等用这个方法,去量各种的性质,如豌豆之大小,树叶中之水脉之数,牝猪右前腿之母络腺(Müller's glands)之数,所得的结果,都是这样的。这样的曲线,叫做错误发见率之曲线(curve of frequency of errors)。依此曲线观之,一类中之一种性质,必有一个平均代表。多数分子之此种性质,皆在平均代表之左右。离平均代表愈远,则分子之数目愈少。这个平均代表,又谓之种的平均(race average)。这样看来,一类生物之性质,各分个俱有微有不同之处,所谓物之不齐,物之情也。但是:倘若只就一种生物之一种性质而言,其不齐之程度,或相倍蓰则有之,然而总不能远离于种的平均,不至于或相什伯,或相千万的地步。
这些事实,和加耳敦祖先定律(ancestral law)极有关系。依祖先的定律而言,一个生物之性质,由遗传而得自每个祖先的部分,等于此代祖先之数之反数(reciprocal,或译交换数)之平方。例如最近的一代祖宗,只有父母二人,则子女得自父母各人的部分,为1/2之平方,即四分之一。那就是说一个生物之先天的性质,有四分之一,是从父遗传下来的,有四分之一,是从母遗传下来的,共总有二分之一,是从最近一代的祖宗遗传下来的。其次最近的一代祖宗,有祖父、祖母、外祖父、外祖母、四人,则孙子孙女,得自祖父、祖母、外祖父、外祖母、各人的部分,为l/4之平方,即十六分之一。那就是说:一个生物之先天的性质,有十六分之一,是从祖父遗传下来的,有十六分之一,是从祖母遗传下来的,有十六分之一,是从外祖父遗传下来的,有十六分之一,是从外祖母遗传下来的,共总有四分之一,是从其次最近的一代祖宗遗传下来的。再往上溯,仿此类推,距离代数更远的祖宗,所遗传下来的部分更少。一直推到安麦帕,我们由它遗传下来的部分,真正是微乎其微了。所以同种的生物,其性质必定大致相同,因为它们多数的祖宗,都是相同的。它们不是“同出于一祖”,它们是同出于多祖。所以寻常的分衍,总是在种的平均所能代表的范围以内,不能发生新种出来。
(B)出轨的分衍 依旧兰马克派说,生物变迁,本无所谓剧变。生物之变迁,是逐渐的、缓行的,不是骤尔而来的。以分个的生物而言,一机关之发展与其退缩,全凭用与不用为转移。用与不用,全凭环境之需要与不需要为转移。需要则用,不需要则不用,固然是人之常情;用则发展,不用便则退缩,却也是天之常理。在分个的方面(ontogenetically),既是如此,在种族的方面(phyllogenetically),也是如此,而且化石学里,还有许多的例子,可以证明。这些发展或退缩的机关,都直接的遗传下去,阅时既久,由小不同而集成大不同。这就是缓变之说。此说颇似近理,而且和常识所预期的,也甚相符,所以在生物学界中,占据势力八九十年,即达尔文自己,也以为生物之分衍,是逐渐的,是偶然的,其所以能成种的缘故,全由于天择之淘汰与保存。
以分个的进化,推较到种族的进化,也有可信的理由。高等动物在胚胎阶级之时,常有一些机关,与低级动物在长成分个之时相同。即以人类而言,其在胚胎阶级之时,心只二房,与鱼(已经长成分个的)相同,脊骨为脆骨,与一种低级鱼相同,后体有尾,与一班哺乳动物相同。又如肱足(蛤蜊)类中之各种,最低级者其壳平滑,较高级者其壳有横埂,再较高级者,其壳上之横埂更显,再较高级者,其壳既有横埂,又有直埂,最高级者有刺。这些刺,就是直埂横埂之交点之发大。但是有刺的肱足在个体发育之时,仍然经过以前几层的阶级。其余与此相类之例尚多。此种事实须有理论方面的解释。负解释之责者,是惟温习之理论(recapitulation theory)。高等动物在胚胎里,由一个细胞——受精的卵——发育成为分个的动物,要经过“自单细胞进化而为人类”所有的历程,就同温习旧课一般;换一句话说,分个方面的胚胎之发育,就是种族方面的进化之缩影,把亿万年的路途,用几个月的工夫走完了。分个方面之发展与退缩,既原于用与不用,则种族方面之发展与退缩,也是根据同样的理由。于是分衍的问题,就可以得到相当的解释了。
但是推较终是推较,不能算稳固的证明。依温习理论而言,种族相近的动物,在胚胎里,其相近的程度,应当更高,且可以高到几乎不可辨别的地步。然而有些相近的动物,在胚胎时的区别,比长成分个时的区别还多,例如节足动物类中之环行蚰(Peripatus)。所以分个进化和种族进化,也不能算作同一的现象。而且在种族进化方面,要发生后天性质遗传的问题;由于用与不用而发展或退缩的部分,如何移注而入于生殖细胞,以遗传于后世,是不易获有明显的证明的。我们只能说:这些发展与退缩的影响,总能修改生殖细胞之内容,到了第二代的时候,如果遇着与第一代发展或退缩时所受的同样环境之刺激,又发生同样的组织而已(参观上章)。
德斐里(De Vries)试验番石榴(CEnothera),而得与缓变之说相冲突的结果。此项植物传种若干代,与原种无殊,及至数代之后,骤尔发生许多不同的新种。所以德斐里创一理论,说生物进化有两个时期;(a)固定的时期,在此时期内,父子沿传不变;(b)骤变的时期,在此时期内,生物向各方面骤尔变迁,而发生不同的种这个变迁,是骤来的,似乎是无因而至的。然而我们与其说是无因而至,宁可说我们不知道它的因罢了。由此看来,生物的进化,不是联续的,但是跳跃的。
这样的分衍,总是因为细胞组织有变迁哪。但是细胞组织之变迁,是从何而来呢?我们取其简便,且把各家理论对于此点可以有相当的解释者,分为二类:(a)由于外的,(b)由于内的。
(a)由于外的 由于外的理论,又可以分为二类:(甲)直接的,如爱莫耳(Eimer)之说。生物组织之变迁,全由外界的刺激而来,如热、光、潮湿、食料、仇敌之类。试举一例而言,植物之有绿色,似乎是常存的性质。然而只是在日光中生长的时候,是如此的,倘若生长在暗室里,则成为无色的。这是因为外面没有日光之刺激,里面就没有叶绿质之发生。总而言之,凡细胞组织之变迁,都是物理的与化学的反应。(乙)间接的,如达尔文之说。外面的环境,并不能促使内部发生变迁;但是许多生物之内部,若有若干不同的变迁,则外面的环境,可以有选择的权柄。其变迁与环境相适应的,即保存之,其变迁与环境不相适应的,即淘汰之。外面的环境,和考试人一样,组织之变迁,和赴考的学生一样。考试人不能直接叫学生去读什么书,但是:若是学生所读的书,与考试人所定的资格相符,就可以中选了。
(b)由于内的 由于内的,也可以分为二类:(甲)生力说(bathmism),如哥布(Cope)所主张的。外界的环境,有一定的需要,生物不能答复这些需要,就不能生存。但是生物都有生存的欲望,生物的生命,是有目的的。因为要达到生存的目的,所以时时刻刻底试验各种方法,来适应这些需要,以满足其生存之欲望。这就是努力(effort),也就是生力(growth force)。因为有这个生力,所以发生组织之变迁,去适应外界的环境。这样看来,组织变迁之中,有心理之分子。(乙)创化说,如柏格森(Bergson)之创化论。每个细胞都是时时进化不已的,时时变迁的,就是时时创造的,而且所有的变迁,无论如何写远,都保存于现存的细胞之中。那就是说:所有过去的历史,都是现在发现的现象之原因。推而论之,这些历史的影响,不一定发见为可觉察的变迁,但是潜伏而不灭,到了一定的时候,可以都发见出来,成一个很猛烈的变迁。所谓剧变者,就是这一种的变迁而已。
除以上数说之外,我们还可依门德耳的原理,而有下列两种解释:
(a)婚配(amphixis)和杂婚(hybridization) 当生物生殖之时,父母的性质,各有一半的机会遗传下去。若父母性质不同,即所生的子女,或与父同,或与母同,例如前段所叙述的豌豆;或与父母俱不同,而另具一新性质,例如白玫瑰与红玫瑰配合,则生绛玫瑰,尖长叶柳与扁长叶柳配合,则生梭形叶柳(在尖长扁长之间),是成一新种。此新种与他种相配合,又可以发生别的变迁。威司蛮以婚配为分衍之唯一的因子。但是如此说法,也有极大的困难,因为由婚配而发生的变迁,不能出于种的平均性质之范围;由杂婚而生的子女,往往不能传种,例如骡,或传种一二代而即绝,例如山羊与绵羊杂婚而生的子孙,若自相配合,不能有绵延之世胤。不过此项传种之限制,似在动物界较大,而在植物界较小而已。
(b)单位性质之互相影响 一个生物之性质甚多,我们可以设想:每个性质,有一个单位的东西,在里边为之主持。这些单位的东西,互相搀搅,其所得的结果,不期定是物理的结果,——彼此相加的总数,它们可以得化学的结果——彼此化合而另生一种新东西。只要这个新东西,以后永不析解开来,也就等于一个单位性质了。
(3)天择
说到天择,我们须将竞争和适应合在一道来讲,因为这三项是分不开的。我们首先且谈生物之生殖的能力;生物生殖之能力,实在是大到可惊的地步。蕨草之一个栭囊(spore sac)中之栭,有一万之多。一对青蝇,可生二千蝇卵,孵出之后,至两星期,又长成生殖期成熟的青蝇;这二千青蝇,又能生二百万蝇卵,每个蝇卵,又孵出一个青蝇。足见生物之所能生殖,远超过于地面之所能供养,僧多粥少,势不能不起争端了。这就是达尔文的生存竞争。近来相信克鲁泡特金学说者,推崇互助主义,以为达氏生存竞争之说,已经完全打破,这实在是戴着伦理的蓝眼镜,去观察天然现象。两个狗为着一块骨头而打架,不能说不是同类的竞争;我们文明人类拼命底防虎疫霉,不能说不是异类的竞争。不过人类有道德的情感,不应该抹煞同情的本能,有推测的智慧,不应该不计算最终的利害;而且这两层之重要,远超出于兽类的简单竞争之上,我们当然是承认的。
生物界中,有同类互助的现象,是为动物之群性(social character)。例如昆虫类之蚁与蜂,其分工合作之组织,可比于法治的国家;飞禽类中之雁与骏鸟(paradise birds),哺乳类中之羚羊,都是出则同阵,处则同居,共同工作,互相守望;他们都是富于群性的动物。又有异类互助的现象,是为同生(symbiosis)。例如豌豆与其根上所附丽的细菌,豌豆以细胞液供给细菌为食料,细菌使空气中之淡固定而为淡合物,以充豌豆之肥料。石南与一种细菌,亦是如此。又有同棲(commensalism)之现象,亦为异类互助之一种。例如寄居蟹寄居于海葵硬管之下端,寄居蟹以海葵为保护之武器,海葵藉寄居蟹而移动,又藉以获取食料。又如一种海鸠与鳄鱼同居,海鸠啄食鳄鱼身上之水蛭,鳄鱼以海鸠为灵敏的警察。又如水母与目虾,虾食水母之分泌液,水母因虾之逃走,而知避免仇敌之袭击。由此言之,狼狈相需而行——如果此故事是真的,——实亦互助之一种,特用之以喻朋比为奸,于是就失去良好的意义了。这是生物互助之一班举例。达尔文也曾经注意到同类互助的事实;他以为这是一种竞争生存的方法,因为有些弱小的动物,不如是不足以生存。但是到了克鲁泡特金的手里,互助之重要之程度因之而加高,互助之责任之意义,也因之而更加神圣了。
达尔文之竞争,并不是专指“夺人之食而食之”,他是注重在如何可以得食的本领。设有一班人去应考,若是有人排挤同僚而获选,是不道德的;若是有人竭力用功“揣摩风气”而获选,并不是不道德的。生物竞争,也是像揣摩风气的。它们变迁其本身的组织,以适应环境之要求,以便生存,并没有若何不道德的意义,而且生物之变迁,不期定要走同一的途径,——不期定都是一样的——有变迁得好的,与环境相适合,就为天然所录取而保存;有变迁得不好的,或是不曾变迁的,与环境不相适合,就为天然所淘汰而灭亡。凡淘汰的,都是不善变迁的,那更不能怪人家变迁得好了。所以达尔文的竞争之理论,与克鲁泡特金的互助之理论,不是根本的冲突,乃是延续的发挥,后者较前者更为详切而已。
为天择所保存的生物,不但可以保存自身,并且可以多占传种的机会,而且可以使他们的后裔之生存的能力逐渐增加。试取一个例来说明这个道理。设若有一处发生一种瘟疫,必定有若干人遭瘟疫而死。假使这些人都没有防疫的知识,则遗剩下来的经过瘟疫而获愈的人,必定是身体强健的,必是细胞可以战胜疫霉的。在此次瘟疫之后,只有这些遗剩下来的人,在那里传种了,而且他们受过瘟疫而获愈,他们对于这个瘟疫的抵抗力,格外加大,这和防疫血清注射,是同样的道理。我们并且可以相信:这个抵抗力,有可以遗传下去的,所以他们的后裔,对于瘟疫的抵抗力,也加大了。例如一种瘟疫对于甲民族是致命的,而对于乙民族没有同等的危险性,这是因为乙民族的祖先,已经受过这种瘟疫多次,而抵抗力加大了。据说:中国人染着白喉症,往往可以获愈,西洋人在中国染着白喉症,往往是致命的。西洋人遇着流行感冒(influenza),有可以获愈的,但是自白人到了美洲,红印度人为白人所带来的流行感冒所杀害的,比他们为白人殖民政策所残伤的,还要多得多。
达尔文的天择——天然的选择——之理论,是从人为的选择比较而来。他看见农夫以及畜牧家之选择种子,而设想天然界中也有一个主体在那里选择,不过这个主体,不是人道的农夫,也不是神道的上帝,它就是天然的环境。自从这个理论发展完备之后,又渐渐回头推到应用的人为的选择上来。农艺、园艺、畜牧各种艺术之中,应用此项原理而收良美的效果者,已是不可胜数。美国有一次有一地方苦旱,有一班人去考查旱地的状况,其中有一少年学生,忽然注意到枯槁的麦田之中,有少数发育如常的麦穗。他想到这几株麦,必定是能抵抗旱灾的。于是用此种麦所结的种子,为普遍的麦种,果然可以避免旱灾。加耳敦竟直成立所谓优种学,要凭藉这个原理,来选择人种,使优秀分子多传继胤于第二世;这也算是进化原理之应用,到了无远弗届的地步了。
与天择的理论同时发生,而其重要之程度较低,而且其后又有渐趋衰落之势的,是为性择之理论。当达尔文创设此项理论之时,华烈司已经表示过不满的意见;因为:就此项理论而言,配偶选择之主权,皆操之于雌性;雄蜘蛛之跳舞而表现其腹部之花纹,雄孔雀之大张其彩屏之尾,以及其嗈喈的鸣声,都是为着要引起雌性之选择;华烈司对于雌性动物审美的本领,颇觉怀疑。达尔文以后也以为“雄性之美丽与强壮者,得传种之优先”一层原理,只能适用于多妻的动物,至于一夫一妻的动物,即美丽强壮之程度较低者,亦有传种之机会。其淘汰与生存,则皆决定于寻常生活之中,而不是决定于生殖的生活之中。假使达尔文之性择之原理,可以推到人类,则世人之劳碌而求富贵利达者,在消极的方面,是要免除室人交谪之困难,在积极的方面,也不过要可以施施从外来,骄其妻妾而已矣。
(4)总论
生物进化,就是遗传、分衍、天择三个因子,在那里动作。这几项动作,都是和物理的现象一般,依照天然定律而进行。所以兰马克说:进化乃是一个天然的现象,所谓天然的者,就是有一定的因果的关系。德斐里说:进化乃是可以用试验方法去研究的题目。达尔文的最大的功劳,也就是把生物现象的神秘性打消,而置之于试验科学管辖权之下。生物进化,就是随环境而变迁,用不着加上一层自不善而进于善,自不美而进于美的意义。
近代人类思想所受于进化理论的影响,大而且广,是不待言的。我们只要看进化二字应用之普遍,就可以发见它在人类思想上所占据的势力。社会也说进化,文艺也说进化,工业也说进化,甚至宗教也说进化。但是在这些所谓进化里边,都有自不善而进于善,自不美而进于美的意思,不是生物进化之原义。我并不是说:我们人类各种动作之进化,不应该趋向于美善之目标;我是说:人类进化,和生物进化一样,只要以生存与进步的欲望为基础,就可以解释一切的动作。所谓生存者,就是获得维持生活之资料,以供给体魄之营养(此指个体生存非指种族生存而言)。所谓进步者,就是不以现在的生存为满足,而求取较好于现在的生存。若是我们要把这种理想的较好于现在的生存,叫做善的美的,却也未为不可。至于实际上还是较好与否,还是美善与否,那只能在结果上去下批评。就这样进步的欲望而言,人类无异于生物,但是人类有发展最高的智慧,既可以预定将来的目的,又可以计划趋向此种目的的方法。从此点说来,即是人类比动物高的不可以道里计的地步。我们以智慧为工具,可以寻出来:有如何的组织可以适合如何的环境,有如何的环境,又可以发展如何的潜能。所以人类进化,不但是讲求变迁里边的组织,以适应于天然,并且要讲求改造外边的环境,使原有的天才不至于淹没下去,这都要凭藉首出庶物的智慧去做工夫。我们不要侈谈尽善尽美的究竟,而抹煞如何得此善美的工作。庄子说:得鱼忘筌,但是:没有筌,如何能够得着鱼呢?
(二)球面沿革
现在我们所居住的地球,有高出海平面约三万英尺的高山,有深入海平面四千英尺的深洋,有广袤十余万方里的高原,有流经三千英里的江河;舟车络绎,树艺繁妩,使我们可以藉此为舞台,以尽人文进化之能事。然而这样可赞美的——或者是可诅咒的——地球上面的形势,所谓山原河海,也不是自地球初生以来就是如此的。它自太阳射出的时候,不过是一团赤炽的气体,因为失热于天空,于是球面凝结而为地壳。地壳成立之后,又经过无数的变迁,方成为现在的形势,而且现在的形势,也不是固定不移的,它又要经受同样的变迁,而发现其他的标样。换一句话说,球面上的各种形势,都是由“现在还在球面上发生变迁”的各种天然主司(natural agency)制造出来,并没有什么神秘的地方。
这些天然主司,可以分为二大类:(1)地外主司;(2)地内主司。地外主司分为空气、水、生物三种。地内主司,分为火山、地震、壳绉三种。大风括土,自高处搬至低处,就是空气可以变迁球面形势之举例。黄河流域之黄土石,都是百万年前由风停积而成的。水能变迁球面之形势,更是显而易见的。大水冲倒堤坝,就是水之破坏的例子;湖底停积泥淤,就是水之建设之例子。水之流动速率大者,破坏多而建设少;水之流动速率小者,破坏少而建设多。生物变迁球面之形势,似乎是不常见的;然而我们试作详细的考察,泽薮之间,有水草蒿莱腐朽成泥,蚯蚓可以化石成土,珊瑚可以长成大岛,植物可以长成煤层,都不是稀奇罕见的事实。至于火山喷发,在我国内部,无从看见;然而现在球面上还是很多的;例如日本及意大利,而太平洋沿岸,都有蜿蜒不绝的火山山脉,所以地理学家说:太平洋有一火带围之。地心之中,储有熔石,如半流体,流动奔荡,若是遇着石层之空隙或弱点,即行溃堤而出,是为火山;若是没有寻着出路,它也可以把上面的石层凸起而成山,这或者也是壳绉之原因之一种;或者可以把上面的石层逼迫而破裂了,这或者也是地震之原因之一种。而且球面之石层,似乎有一定的处所,为固定的大块(blocks)。例如非洲大陆、欧洲大陆、印度半岛、中央亚细亚、太平洋海底,皆是这些大块,它们自有地质历史以来,不常随壳绉而升降,但是前后左右互相移动。凡夹在大块中间的处所,名曰地渎(geosynclines)。凡地渎所在的地方,受两边大块之逼迫或延张,最易于发生山陵起伏之壳绉,以及火山地震各种现象。至于大块移动之原因,或者是因为地球旋转,地壳之各部失了力之平衡,或者是因为地心镭质之发热,下面之石层逐渐熔化,而上面漂浮的地壳,渐入不固定的状况。总之,地内主司之原因,较地外主司之原因,较难解释,因为它们是较难为我们所考察的。
地外主司之动作,是使球面自不平而渐趋于平。地内主司主之动作,是使球面自平而趋于不平。现在球面之形势,就是这两类主司同时动作之结果。以上所说的一段,就是地质变迁论之大意。我们在下面再说球面变迁及生物进化之历史大纲。
(三)地质历史
生物变迁和形势变迁之理论之大致,既已如上文所陈述了。把这些变迁之事实记录下来,成一个有系统的历史,就是地质历史。地质历史和人类历史一样,也分有期代之不同。地质历史之分期,即以石层中化石所代表的生物等级之高下为标准。成立较早的石层,其中所含的化石,必定原于等级较低的生物;成立较迟的石层,其中所含的化石,必定原于等级较高的生物。依化石所代表的生物之等级,地质学家把地质历史分为四期:(1)无生期;(2)古生期;(3)新生期;(4)人生期。古生期又分为初古中古二期,新生期又分为中新次新二期。
(1)无生期
在无生期时,球面温度甚高,或者水中溶解物质过多,尚不适宜于生物之生活,所以没有生物,因此没有化石遗留下来。此时球面之大半,皆为海水所遮蔽。凡此期内成立的石层,因为阅时过久,所经历的地壳变迁过多,都改变了它们原来的组织,而成为变形石。
(2)初古期
此期中之动物,都是海居的无脊椎类。此期中之植物都是海藻。所以此期又叫做无脊椎期。然而当此期之终局,动物已有鱼类,植物中已略有蕨类发现。此期中最重要的生物,在前半为三叶(Trilobita),在后半为笔形(Graptophita)。三叶归节足门,与近代生存之虾略同,笔形归腔肠门,与近代生存之珊瑚略同。二者属种繁多,各具形态不同;然取其共相言之:三叶身分十二节,每节有附支一双,但是附支发达甚大,和它的正体成川字形,所以叫做三叶。头部有数节熔化而成的头盔,左右有二眼。因为在深海里要看得清楚,所以它们的眼睛有甚大者。头旁有二长刺,是保护身体的。笔形为一个联合同生的动物,有干若干支,每干上有水孔若干,每一水孔,乃是一个独立的生物,其内有肌花可以伸缩,干内有公管,水自孔流入,在公管内联合生活,与现今生存的珊瑚无异。
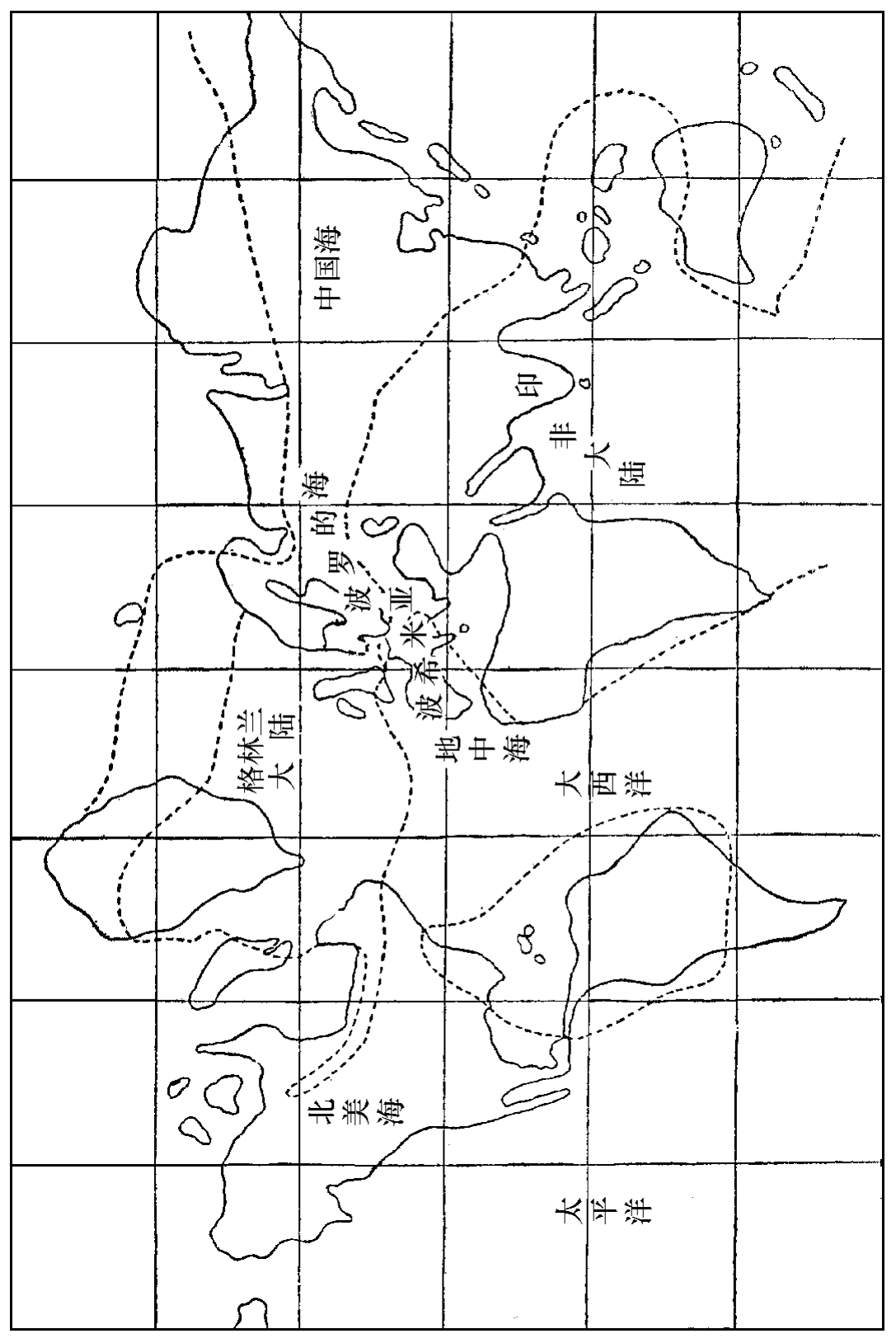
初古期中(奥岛代)球面形势之大概
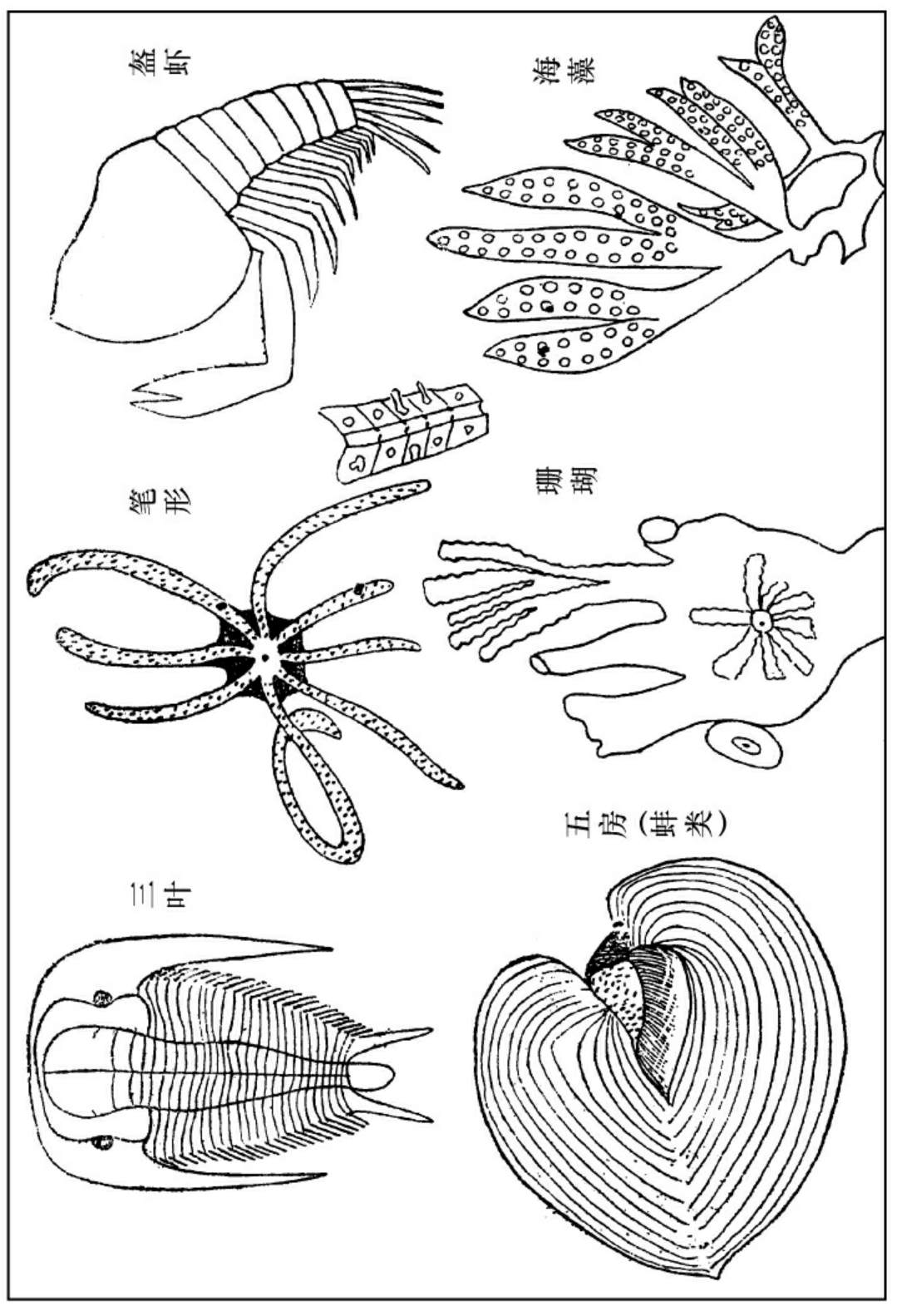
初古期中生物之一班
此二类生物,可以为初古期之全权代表。因为此二类生物世系甚短,笔形在此期未完之前完全灭绝,三叶也几乎完全灭绝,而且此二类生物分布之区域极广,球面各洲,几乎无处无之,所以它们是此期中化石之标准。
当此期之初叶,欧洲与北美之间,有一个北水大陆,即格林兰大陆;非洲与南美之间,有一个非巴大陆。今日长狭之大西洋,在此时期,乃是中间有海而南北皆有大陆。我国北部,此时似亦为陆地。至中叶时,海之面积逐渐推广,东有中国海。非洲与南美之间,亦低降成海,而非洲与印度联成大陆,为印非大陆。欧东有波罗的海,欧西南有波西米亚地中海。这些海都是相通的,因为各处的笔形都是一样的。至此期之末叶,有些处所,受了地壳引起之影响,海之面积又退让,如来因河流域、波罗的海、乌拉山、格林兰及北美西境都是的。
(3)中古期
中古期之生物,以鱼和隐花植物为最盛,所以此期又叫做鱼期。世界煤层之大半都成立于此时,所以又可以叫做煤期。其余无脊椎类,如肱足、珊瑚,也是很多的。此期中之鱼虽多,然而都是属于硬鳞目的,而非高等鱼类。头上多有甲,身上有硬刺,尾为无叉,骨多为脆骨,这都是刚自无脊椎类进化而来的证据。此期中之植物,皆为隐花植物之蕨类。然而因为此时二养化炭充足,温度和暖,有高至十余丈者,不像今日生存蕨类之凤尾草那样矮小。此期中陆地之上,草木繁盛,所以有两栖动物发生。至此期之末叶,两栖动物已经代鱼而为主人翁了。爬行动物也在此期之末叶刚才发见。又此期末叶,居于海内碱湖的生物,都是现退缩不昌的状态。
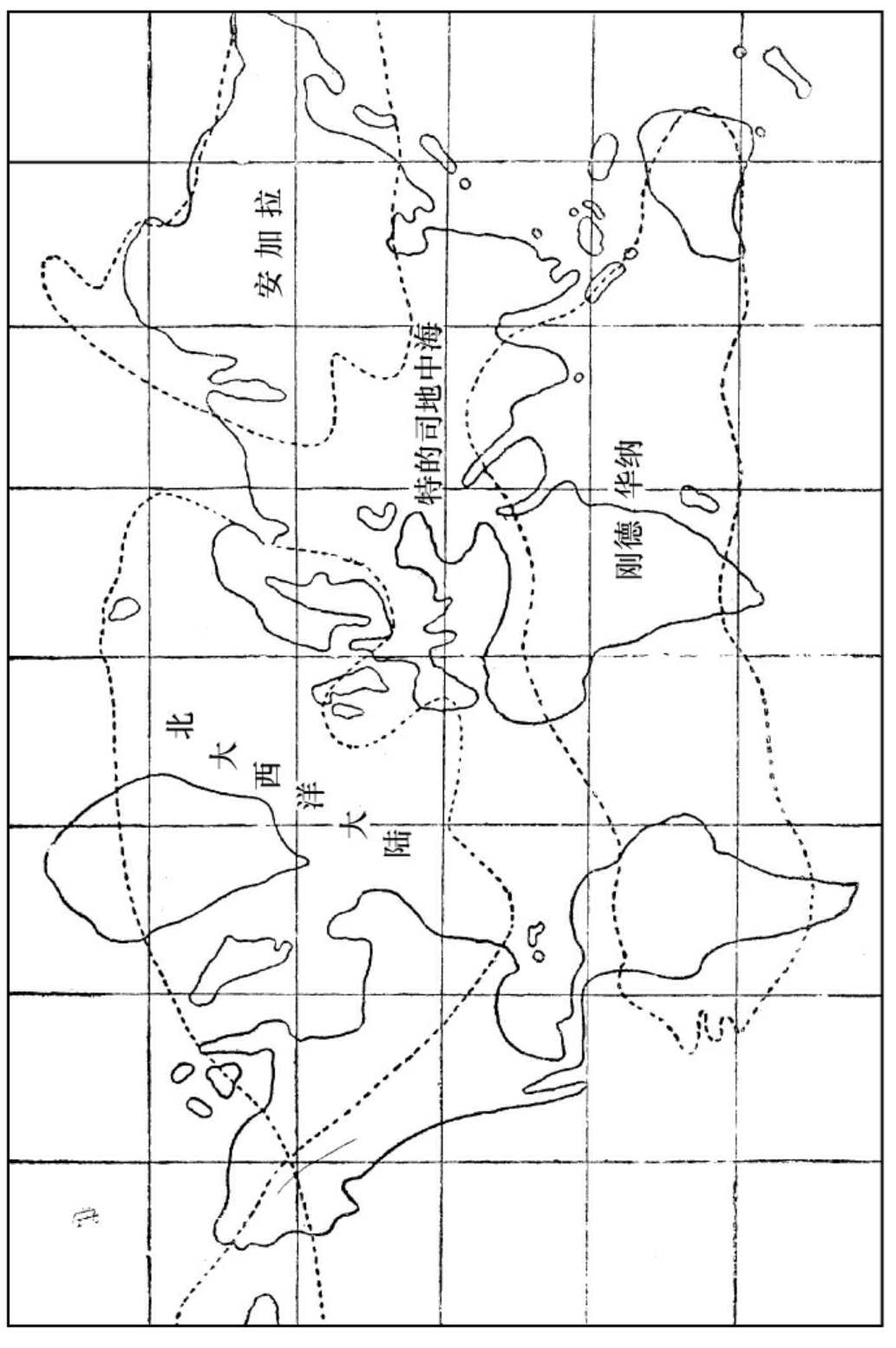
中古期中(石球代)球面形势之大概

中古期中生物之一班
当此期之初叶,各处的海洋都逐渐侵占陆地。欧亚之间,或有一欧亚海。北美洲亦或有一北美海。北大西洋中有一大陆,叫做北大西洋大陆,即初古期之格林兰大陆所引起而加大者。沿此大陆之旁,为旧红沙岩停积之处。南半球亦有一大陆。至中叶时火山发现甚多,故地面变迁亦极多。然而要其总结果而言,北半球有一大陆,联接欧亚北美,叫做安加拉(Angora);南半球亦有一大陆,联接南美非澳,叫做刚德华纳(Gondwana)。此二大陆之间,有一个地中海,叫做特的司(Tethys)。其最要的特点,是:此时地面都高出海面不多,故易于侵没,又易于升起,所以长成极厚的煤层。煤层上之灰石岩极厚。由此可推当时空气中之二养化炭极多。因为这些灰石岩,都是直接底或间接底由海水吸收空气中之二养化炭而成的。直接的吸收,是化学的,由于碱性钙镁之结合;间接的吸收,是生物的,由于多孔动物之甲壳之生长。煤层中之植物,和现在热带植物相似,由此可推当时气候温和。至此期之末叶,北半球火山发现甚多;北美之东因此而成阿巴拉其山脉(Appalachian chain),欧洲因此而成许多的内海。其所以易成内海者,也是因为陆平面与海平面相差不多的缘故。
(4)中新期
中新期中之生物,爬行极多,所以此期又叫做爬行期。飞禽哺乳,俱发见于此期之末叶。凡古生期中之生物,如植物中之古蕨类,动物中之盾首鱼,至此期中都淘汰干净了。所以中古中新二期之交,乃是生物进化之大关键。
此期中之生物,自然以爬行为主。此期中爬行种类极多,有居海的,有居陆的,有能飞的,有肉食的,有素食的,有犬齿的,有平齿的,有乱齿的,有有喙的。它们的躯体都很壮伟,多半有四五十尺长,甚至有长至百尺的。有鱼蝎,其爪轻薄如翅,可以泅水。又有邻蜴,其爪亦如棹,如现在之海狮,亦能泅水。这都是居海的。又有半陆半海的,如凶蜴,可以人立而行。凶蜴有素食的,有肉食的,其素食者更大。有翼蜴,其前肢之拇指极长,拇指及身之间有皮联之,可供飞行之用,略如现今之蝙蝠。若教现在生存之龟蛇数典而念其祖先,应该有不胜今昔之感慨了。
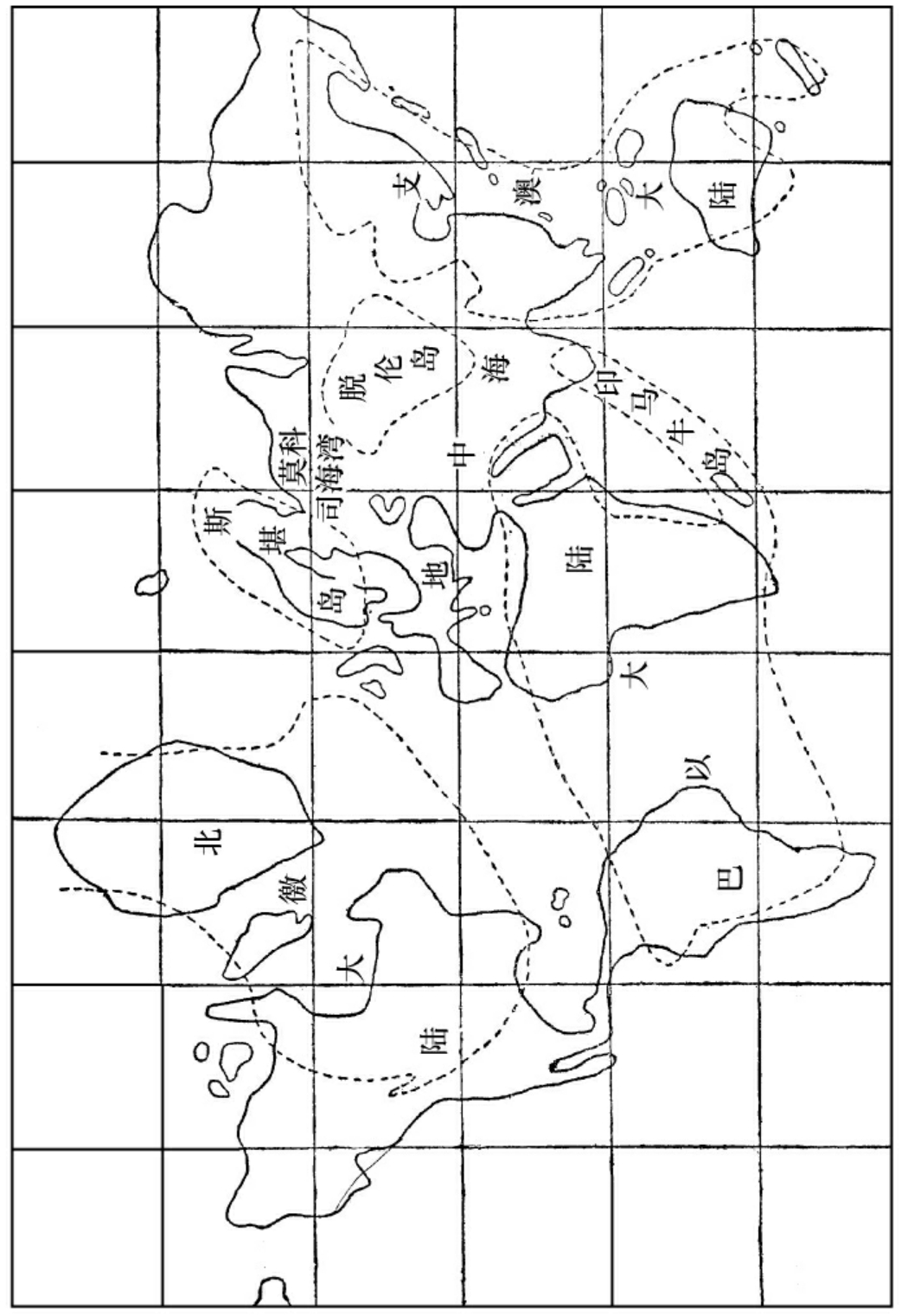
中古期中(侏罗代)球面形势之大概
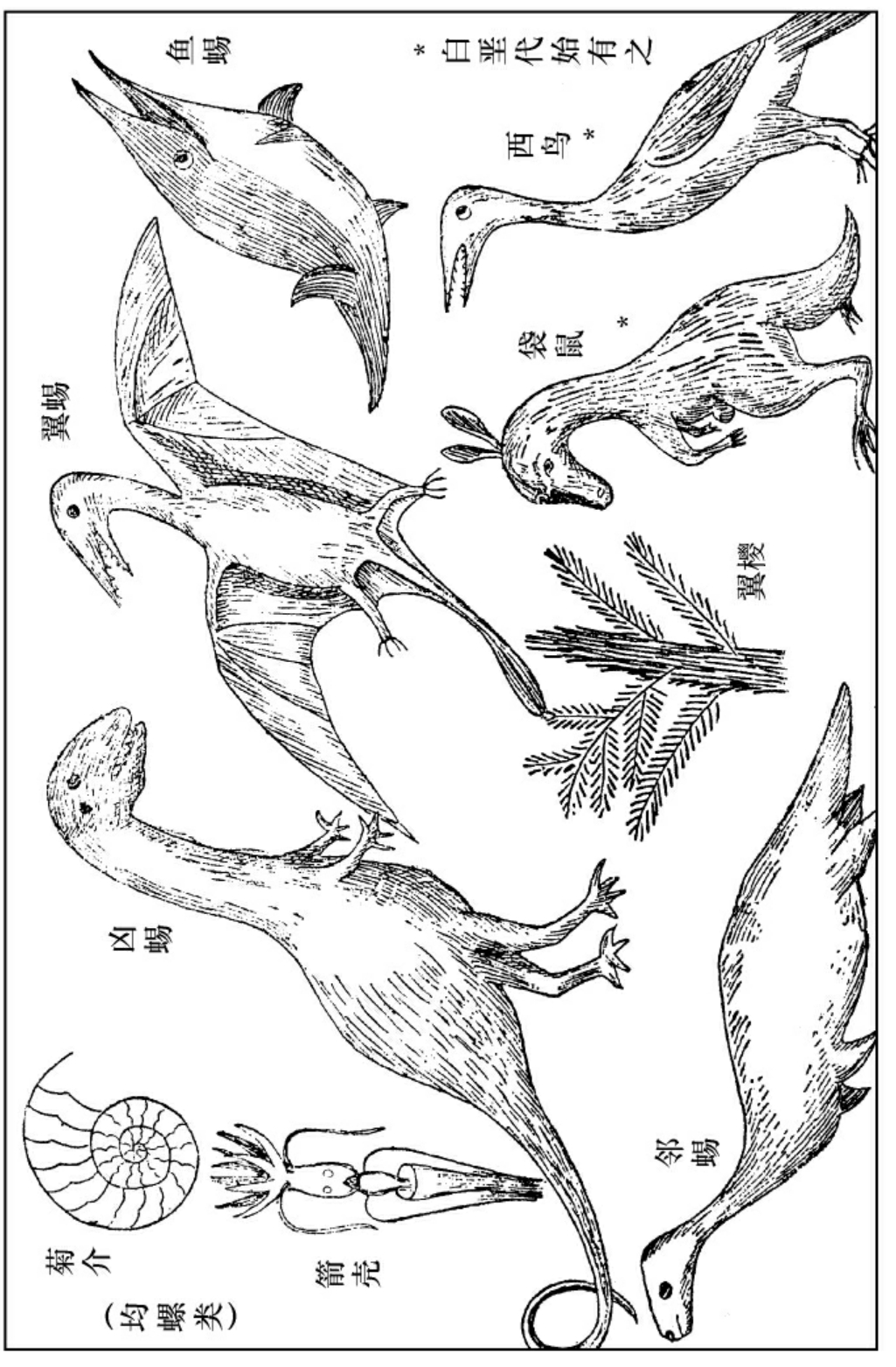
中古期中生物之一班
飞禽之见于此期者,皆有齿而平胸,因为它们尚未脱除爬行之性质。但是飞禽或者是哺乳发见未久之时,从哺乳发生出来的横出一枝的支派,不是从爬行直接而变演出来的。然而此期之飞禽,总是等级甚低,与爬行还是很接近的。
哺乳之见于此期者,皆为有袋类(如袋鼠),或单管类(如鸦嘴兽),其肢长而尾甚大,与此期之爬行略同。
植物中蕨已渐衰, (叶干类)松(球实类)等裸子植物起而代之。包子类之植物,亦已见于此期之末叶。
(叶干类)松(球实类)等裸子植物起而代之。包子类之植物,亦已见于此期之末叶。
当此期之初叶,欧亚之间,极多内海,还是上期遗留下来的。然而内海之面积逐渐扩张,自阿耳皮斯至中国,皆为大海。至中叶时,有火山发现,东方成一大陆,叫做支澳大陆。北美洲成一大陆,叫做北徼大陆。南大西洋成一大陆,叫做巴以大陆。欧亚之间,有二大岛,叫做斯堪底岛、脱伦岛。海与岛之间,海陆交错甚多,所以有许多三角洲停积。至此期之末叶,海面又加广。自西欧至西亚,皆为海,亚东澳西亦为海。北美之西,和南美巴西亦为海。至中新次新之交,始有壳绉行动,造成现今之大陆。
(5)次新期
次新期中之生物,以哺乳为主了。中新次新之交,也是生物变迁形势变迁中之关键。中新期中霸盛一时的爬行动物都淘汰了,即令有存在的,也把它们的原来的组织变掉了。无脊椎中之菊介、箭壳也淘汰完了,又有他种较新的生物出来代替它们。质言之,现今生存的生物,都发迹于此期之中。赫胥黎研究出来的马趾之进化,也是在此期发见的。至于此期中之软体动物,和现在生存的种类相同的极多;因为从此期到现在,没有什么剧烈的形势变迁了。植物中之包子类,也逐渐滋盛于此期之中。总之,此期是开现代的先声的时代。
当此期中,火山发现甚多,造山壳绉之运动亦甚显明。现今球面之最高山,如喜马拉亚、阿耳皮司、安第斯、落机诸山,都是此期中之壳绉运动所规定的。我国辽东山东之玄武岩,似乎也是此期中之火山所喷出的。德国著名的褐煤,亦成立于此期。
(6)人生期
人生期中之生物,自然以人为主了。当此期之初叶,有冰川广被欧洲北美,其流及之区域甚广,有如现今之北冰洋、南冰洋。故有谓地球旋转之轴迁移地位者。他处亦有片段的冰川。人类始祖生活于这种冰天雪地之中。他们都是洞居的,有毨象与之为伍。此种毨象多绒毛,能御寒,不像现在热带的象。但是近来有人找出来在次新期之末叶,已经发见人类之遗迹,则人类之产生,实在人生期之前,不过经过很长久的野蛮时代,与木石居,与鹿豕游,方才到了有文字记载的历史时期。所以布郎说:夜比日长得多,考古学家的责任,就是拿烛光照明此夜之黑暗。他们把人类进化分为三代:一曰石器代,二曰铜器代,三曰铁器代。铜器之发明所以较铁器为先的缘故,是因为铜矿有鲜丽之颜色,较铁为易寻,而铜之炼冶艺术又较铁为简易得多。石器代又分为二代,一曰古石器代,二曰新石器代;古者粗率不文,新者形式美备。古石器代又分为二代,一曰毨象代,此代人类与毨象同居,此种毨象,现今已无生存者;二曰雪鹿代,此代人类与雪鹿同居,与现今哀司基莫人略同。但是以上所分之代,自然也不是普遍全球的。现今各民族之文明程度不同,古代各民族之进化,也可以有先后不同的。
考古学家掘取此时人类之遗迹,已有雕刻图像的石壁,有殊生动可观者。足见初民也有审美的情感,因为这些雕刻对于他们,决无实用的价值。
以上所说的一切,可以归到一个哲学原理之下,曰进化,曰变迁。把这个观念应用到一切世界上去,或者还有讨论的余地,但是物质的世界——包有机世界而言——是进化的,变迁的,已经是无可辩驳了。
从生物学里边,我们知道许多生物进化的理论;从地质学里边,我们看见许多生物进化的证明。进化就是变迁。生物变迁不已,并没有一定不移的种。所谓种者,就是有一定的形态、机能、习惯,与他种有不同的地方。生物的种,既不是一定不移的,则其形态、机能、习惯,自然都有随时流动的性质。人类为生物之一,当然也逃不出这个普遍的公例。推而论之,人类的语言、文字、典章、制度、风俗、礼教、行为、思想,无时不可经受适应的变迁,以期达到生存的目的。不过人为万物之灵,其生存之意义,较之生物生存更为深远而已。古话也曾说:穷则变,变则通,变迁并不是什么稀罕的事情。所以我们看见“自古未有”的东西,用不着瞠目而咋舌。从十九世纪的人看来,自古未有出没云霄的飞艇;但是从中新期中之伟大的爬行动物看来,从来没有能用口舌说话的人。
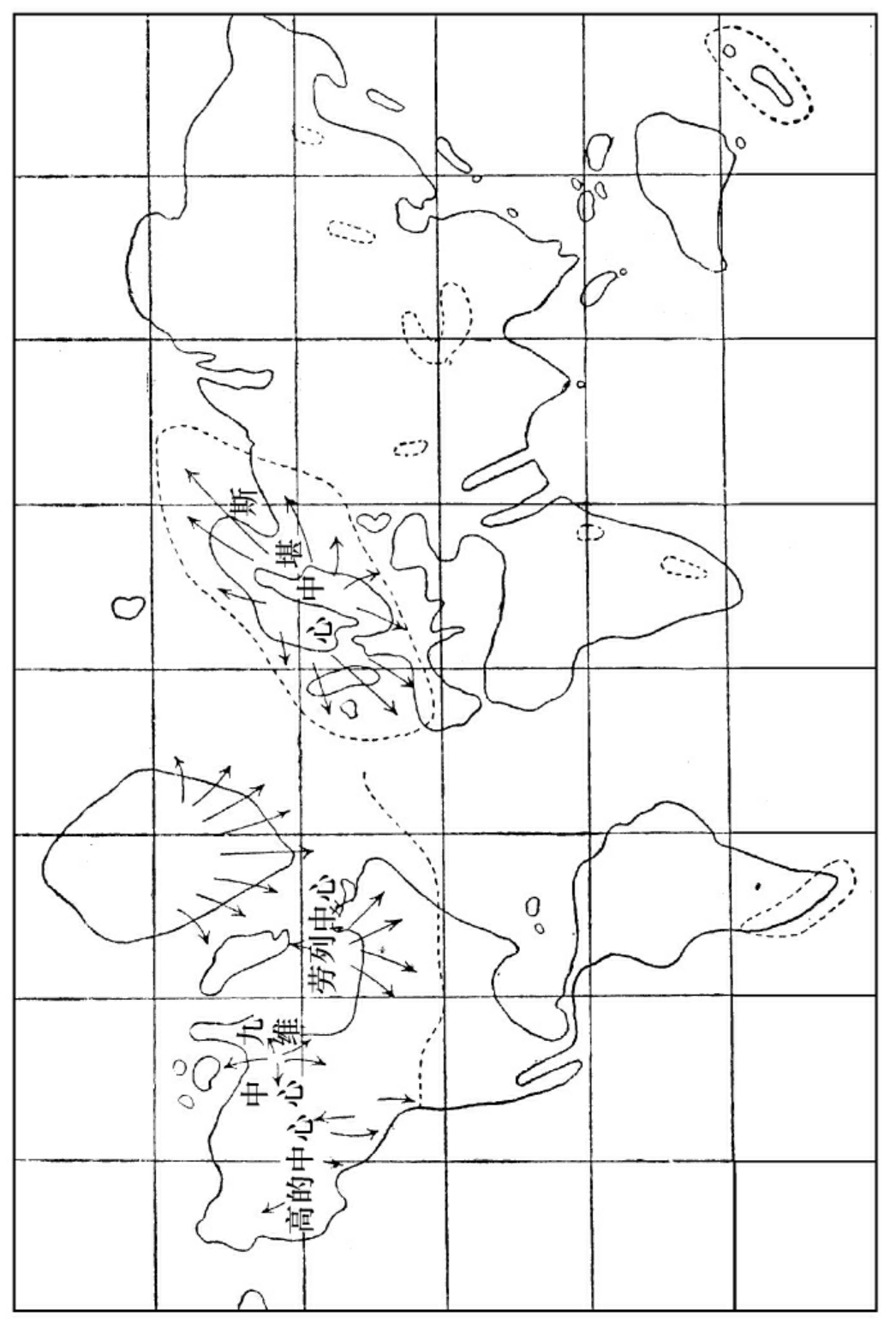
人生期中冰川世中兵之分布
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










