



第三节 抗原递呈
T细胞借助其表面TCR识别抗原物质,但一般不能直接识别可溶性蛋白抗原,而仅识别与M HC分子结合成复合物的抗原肽:CD4+T细胞识别APC表面抗原肽-M HC-Ⅱ类分子复合物;CD8+T细胞识别靶细胞表面抗原肽-M HC-I类分子复合物。细胞将胞浆内自身产生或摄入胞内的抗原消化降解为一定大小的抗原肽片段,以适合与胞内M HC分子结合,此过程称为抗原加工(antigen processing)或抗原处理。抗原肽与M HC分子结合成抗原肽-M HC分子复合物,并表达在细胞表面,以供T细胞识别,此过程称为抗原递呈(antigen presenting)。APC或靶细胞对抗原进行加工与递呈,是TD抗原诱导特异性免疫应答的前提。
根据被递呈抗原的来源不同,可将其分为:
1.外源性抗原(exogenous antigen):来源于细胞外的抗原,如被吞噬的细胞、细菌或某些自身成分等。APC加工处理外源性抗原后形成的抗原肽,常由M HC-Ⅱ类分子递呈给CD4+T细胞,此为溶酶体途径或M HC-Ⅱ类途径。
2.内源性抗原(endogenous antigen):是细胞内合成的抗原,如病毒感染细胞所合成的病毒蛋白、肿瘤细胞合成的蛋白以及胞内某些自身正常成分等。内源性抗原在胞内加工后形成的抗原肽则与M HC-I类分子结合,递呈给CD8+T细胞,此为胞质溶胶途径或M HC-I类途径。
一、外源性抗原的加工、处理和递呈
(一)外源性抗原的加工处理
APC通过胞吞作用(endocytosis)或称内化作用(internalization)而摄入外源性抗原,包括吞噬、吞饮或受体介导的内吞作用。所摄入的外源性抗原由胞浆膜包裹,在胞内形成内体(endosome),逐渐向胞浆深处移行,并与溶酶体融合形成内体/溶酶体。内体/溶酶体中含有组织蛋白酶、过氧化氢酶等多种酶,且为酸性环境,可使蛋白抗原降解为含13~18个氨基酸的肽段,适合与M HC-Ⅱ类分子结合(图9-7)。
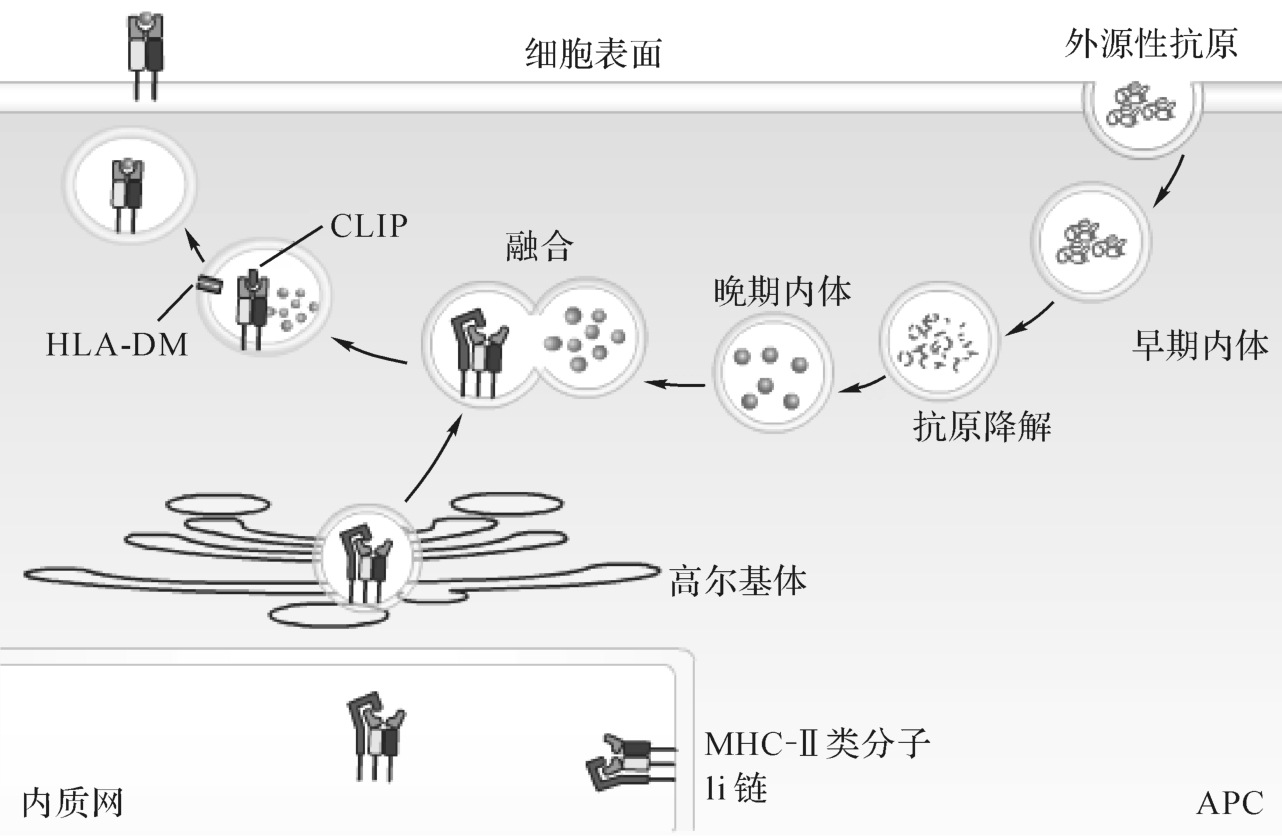
图9-7 外源性抗原的加工、处理和递呈
(二)M HC-Ⅱ类分子的生成和转运
M HC-Ⅱ类分子α链和β链在粗面ER中生成,并在钙联蛋白参与下折叠成异二聚体,插入粗面ER膜中。粗面ER膜上存在Ia相关的恒定链(Ia-associated invariant chain,Ii链),与M HC-Ⅱ类分子结合,形成九聚体(abIi)3复合物。Ii链的作用是:①参与α链和β链折叠和组装,促进M HC-Ⅱ类分子二聚体形成;②阻止粗面ER中内源性肽与M HC-Ⅱ类分子结合;③促进M HC-Ⅱ类分子从ER移行,经高尔基体进入M IIC。
胞内合成的M HC-Ⅱ分子被高尔基体转运至一囊泡样腔室,后者称为M HC-Ⅱ类分子腔室(M HC classIIcompartment,M IIC)。含外来抗原多肽的内体/溶酶体可与M IIC融合。随后,在酸性蛋白酶作用下,使与M HC-Ⅱ类分子结合的Ii链被部分降解,仅在M HC-Ⅱ类分子抗原肽结合槽中残留一小段,称为II类分子相关的恒定链多肽(classII-associated in-variant chainpeptide,CL IP)。
(三)M HC-Ⅱ类分子组装和递呈抗原肽
M HC-Ⅱ类分子的α1和b1功能区折叠为2个α螺旋和1个β片层,形成抗原肽结合沟槽,其两端为开放结构,使与之结合的多肽在N端及C端可适当延伸,最适的多肽长度在13~18个氨基酸之间。
存在于M IIC中的M HC-Ⅱ类分子,其抗原肽结合槽由CL IP占据,故不能与抗原肽结合。H LA-DM分子(属非经典M HC-Ⅱ类分子)可使CL IP与抗原肽结合沟槽离解,此时抗原肽才可与M HC-Ⅱ类分子结合为复合物。抗原肽-M HC-Ⅱ类分子复合物随M IIC向细胞表面移行,通过胞吐作用(exocytosis)而表达于细胞表面,供CD4+T细胞识别,完成外源性抗原肽递呈过程。
二、内源性抗原的加工、处理和递呈
(一)内源性抗原的加工处理和转运
胞内合成的内源性抗原在胞浆内被处理和转运。内源性抗原在多种酶和A TP的作用下与泛素结合,泛素化的内源性抗原被解除折叠,以线形进入蛋白酶体(proteosome)。蛋白酶体(20S)是存在于细胞内的一种大分子量蛋白质水解酶复合体,具有广泛的蛋白水解活性。蛋白酶体为中空(孔径约1~2nm)的圆柱体结构,内源性蛋白通过蛋白酶体的孔道,可被降解为含6~30个氨基酸的多肽片段。蛋白酶体由4个各含7个球形亚单位的圆环串接而成,其具有酶活性的组分主要是两种低分子量多肽(low molecular weight peptide,LM P),包括LM P2和LM P7(属于非经典M HC-Ⅱ类基因产物)(图9-8)。
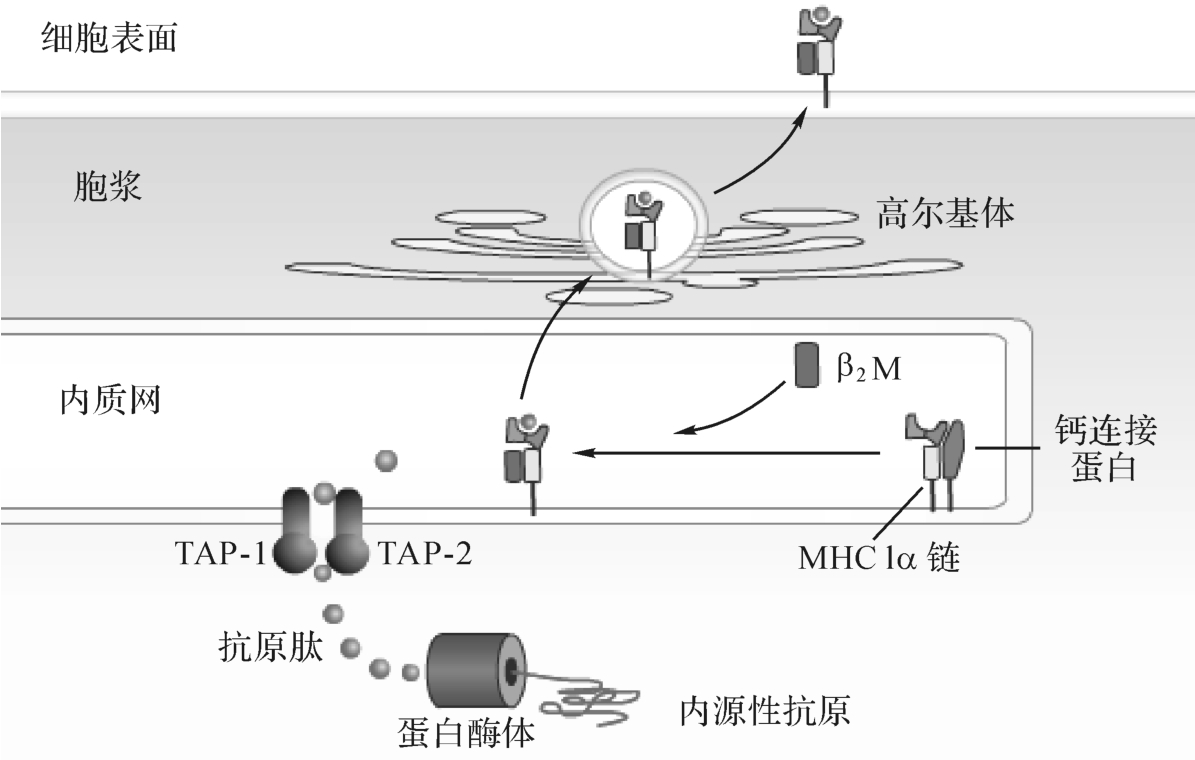
图9-8 内源性抗原的加工、处理和递呈
经蛋白酶体降解的抗原肽片段须进入内质网(ER)才能与M HC-I类分子结合,该过程依赖于ER的抗原加工相关转运体(transporter associated w ith antigen processing,TAP)。TAP由TAP1和TAP2两个亚单位组成,是ER膜上的跨膜蛋白,各跨越ER膜6次,共同在ER膜上形成孔道。
胞浆中的抗原肽先与TAP的胞浆区结合,在TAP分子的A TP结合结构域作用下,使A TP降解,导致TAP异二聚体结构改变,孔道开放,抗原肽通过孔道进入ER腔。
TAP可选择性转运适合与M HC-I类分子结合的肽段,其机制为:①TAP能选择性转运含8~12个氨基酸、适合与M HC-I类分子结合的抗原肽;②TAP优先选择C端为碱性或疏水性残基的多肽片段,这些残基乃抗原肽与M HC-I类分子结合的锚着残基。
(二)M HC-I类分子的生成和组装
M HC-I类分子的重链(α链)和轻链(β2m)在粗面ER中合成后,被转运至光面ER。在ER中,M HC-I类分子须立即与某些伴随蛋白(chaperone)[如钙联蛋白(calnexin)、钙网蛋白(calreliculin)和tapasin]结合。此类蛋白的作用是:参与α链的折叠及与β2m组装成完整的M HC-I类分子;保护α链不被降解;帮助M HC-I类分子与TAP结合。
(三)M HC-I类分子组装和递呈抗原肽
在伴随蛋白参与下,M HC-I类分子组装为二聚体,其α链的α1及α2功能区构成抗原肽结合沟槽,沟槽的两个侧面为α螺旋,底面为β片层结构。M HC-I类分子沟槽纵向的两端是封闭的,能结合含8~12个氨基酸的多肽。
M HC-I类分子与ER上的TAP相连,再与经TAP转运的抗原肽结合,形成抗原肽-M HC-I类分子复合物,然后与TAP、伴随蛋白解离,移行至高尔基体,通过分泌囊泡再移行至细胞表面,递呈给CD8+T细胞。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










