



专题三:生命也是数学设计的
1946年,爱丁顿著《基本理论》中明确地断言:我们的头脑可以从先验知识中建立一门关于自然的纯科学。这门科学是唯一确定的,任何其他的都会有逻辑上的矛盾。因而从我们的头脑中可以获知光速是有限的,甚至自然中的常数——例如质子质量与电子质量之比——也可以先验地确定,这种知识独立于宇宙的实际观察并且比经验知识更加确定[1]。
爱丁顿毕竟是著名的天文学家,断言宇宙常数独立于宇宙的实际观察。
2000年8月15日[爱]《纽约时报》刊登困扰世界的10大物理学难题之二的文章:“为什么宇宙常数有它自身的数值?它是否为零?是否真正恒定?”恐怕到现在也未取得共识。
但从这段文章虽然不长却有一句掷地有声的话——宇宙常数有它自身的数值来讲,这也就是爱丁顿所深信的“自然是数学设计的”。
本文加题所示的要旨,就是认为生命也是数学设计的,如图2‐3‐1(a)。
这样的夸大其词,要请读者让我从头说起。
1984年11月,联合国教科文组织(UNESCO)《信使》杂志《宇宙的形成》专辑,美国宇宙科学家卡尔·萨根(Carl Sagan)《生命的奥秘》一文,竟然采用我国2000年前的一幅古画——伏羲—女娲人身蛇尾画,并和英国爱丁堡天文台摄制的DNA双螺线结构展示在文章的中心位置上,如图2‐3‐1(a)所示。

图2‐3‐1
这就好像在说,生命起源于一张画或者说伏羲—女娲是人类的祖先了(据报道:100万年前,地球上的人口大约为5万人)。
那么到底有什么样的数学设计,可以用来说明生命也是数学设计的呢?有没有这样的数学设计呢?它在哪里呢?这个问题,我花了五年时间不得其门而入,也就是说,生命起源的难,表现在这两张图像上无法用某种数理机制来解答它们两者的数学上的自洽关系,好像著名的51号照片在尚未被罗莎琳德·富兰克林(1920-1958)发现之前,全世界都无从得到哪怕是很微小的线索以便进而探索生命起源问题。
而我好在幼小读过《三字经》,上有这样的语句:
曰水火,木金土,此五行,本乎数
然后作这样的数学分析,并立即显示双螺线如图2‐3‐2所示。
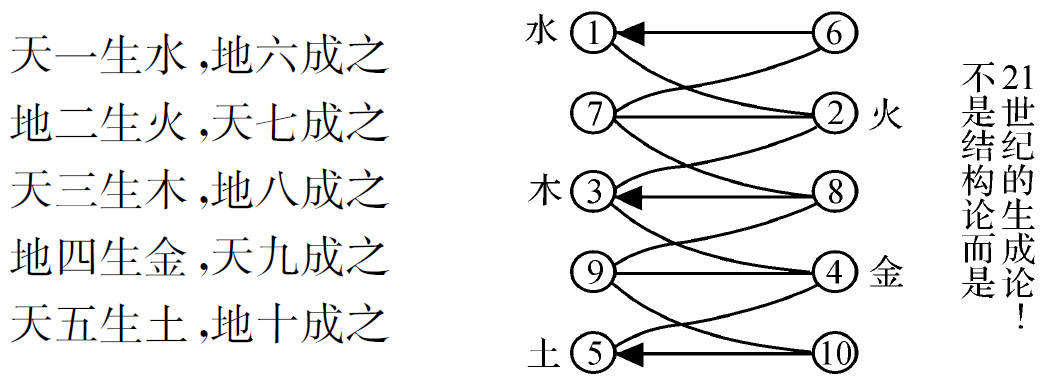
图2‐3‐2
这种双螺线结构怎么样,用51号照片比比看,并用当代双螺线拼板图比比看(见图2‐3‐3)。
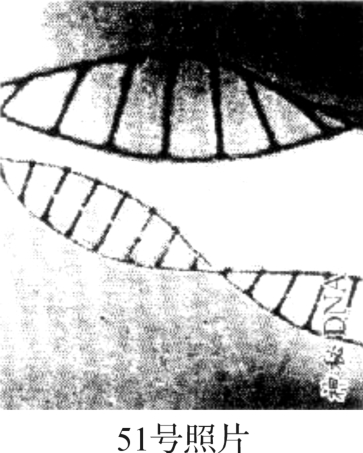
图2‐3‐3
①生命是从照片开始设计的(见图2‐3‐4)。

图2‐3‐4
②生命是用插头插座拼板设计的(见图2‐3‐5)。
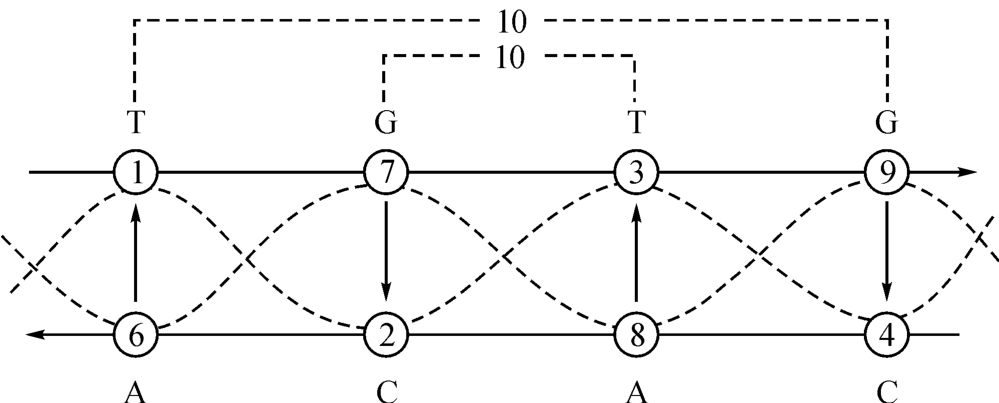
图2‐3‐5
③生命是数学设计的,这种数学是什么货色?草根。但是,西方学者无法数字化,而只好用拼版式(见图2‐3‐6)[2]。
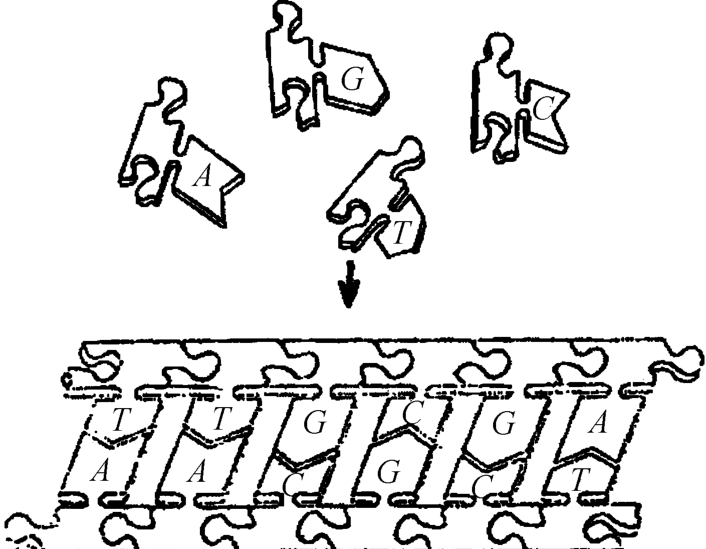
图2‐3‐6
从画面看,这里有一个组装过程:先是A和T、G和C相组装,好像“插头”和“插座”相结合面为一块“拼板”如图2‐3‐6。
这是一种“构成论”,在描述生命起源的自然现象上,认为变化是不变要素的结合,而不是主张变化是“产生”和“消灭”或者“转化”的“生成论”。爱因斯坦的质能关系,就是一种质能转化的当量关系。
相比较而言,图2‐3‐2就是一种“生成”模式。它的古老术语为:“天一生水,地六成之;地二生火,天七成之,天三生木,地八成之;地四生金,天九成之。”其间,生数成数的互相变换,显示出一种“以奇生者成而偶;以偶生者成而奇”(王安石)的规律性,而与古希腊毕达哥拉期学派“奇偶相生而成数”的理念和强调对立面的“协和”和“和解”的观点相呼应。同时,“1-7-3-9”和“6-2-8-4”这样两组不相连缀的离散数,却因奇—偶相生原理达成“1→2→3→4”和“6→2→8→4”相映成趣的双螺线结构和一个有着共同耦合常数“5”的奇—偶成对并协变量系统,假如海森堡(W.K.Heisenberg)还健全而且被他看到,他一定会这样说:“这些纵横关系……以一种简单得不能再简单的方式呈现出来……我们只能谦恭接受下来的礼物。即使是柏拉图也可能不相信它们是如此之美,因为这些纵横关系不可能被发明。他们开天辟地以来就一直存在。”[3]
海森堡“开天辟地以来就一直存在”的说法,中国话叫“天地神物”。再者,这里说的中国古已有之的生成思维又竟与今天的天然科学思想方法由构成论走向生成论的趋势相合,这真以古为新了。
由此出发,我们不妨推想几十亿年以前,生命起源的自组织过程如图2‐3‐7所示。
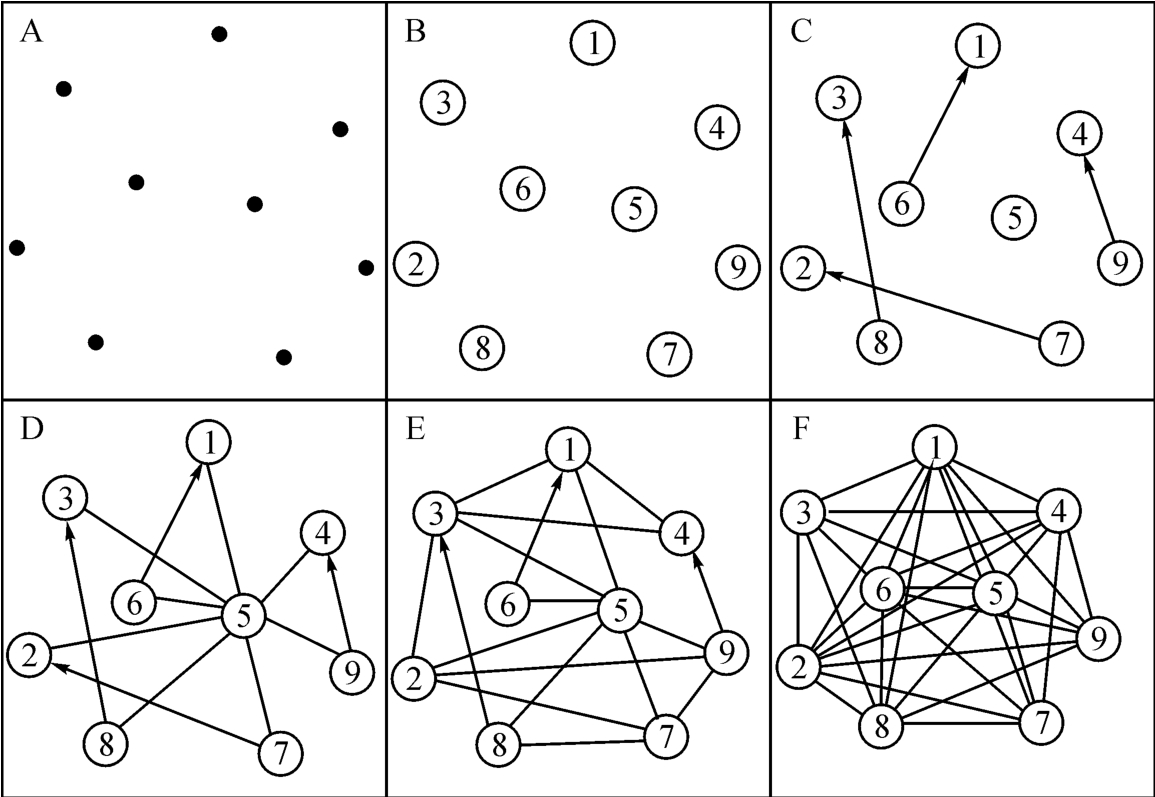
图2‐3‐7
似可认为:远古时期,闪电和太阳光使原始大气中的富氧分子分裂而落到海水中,成为一种原始分子核苷酸。图2‐3‐7(A)的九个点代表这种原始分子。在引入《洛书》结构原理后,如图2‐3‐7(B)所示,这种原始分子有了分奇(阳)和分偶(阴)的区别。在一定的能量条件下,这种分子在混沌的热骚动中产生某种自然尺寸而结对存在。作为一种量,是向量。不但表示一种自组织的方向,而且表示力的大小,如图2‐3‐7(C)所示,分子聚合成为一种有机体,从点到线,到面进而为体,不断复制的结果。这种有机体也不断发展而复杂起来。图2‐3‐7(E)和图2‐3‐7(F)说明这一点,表现在矢量加法三角形的个数上,从图2‐3‐7(D)所示的“△1+5=6”,“△2+5=7”,“△3+5=8”和“△4+5=9”以至于逐渐增多,多到不胜枚举的图2‐3‐7(F)。
原来,我们的《易经》就是讲究变易的学问。它的基础结构是“简易”的,如图2‐3‐4所示。但它的表现形式却是无穷的。
有道是:“易之为道也屡迁,变运不居,周流六虚,上下无常,刚柔相易,不可为典要,唯变所适。”(《系辞·下传》)它的《洛书》式描述如图2‐3‐8所示。
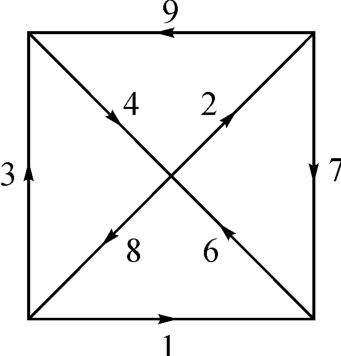
图2‐3‐8
但有一点必须明确,这种“变易”又有其守恒“不易”的一面,那就是必须奇(阳)偶(阴)互补以与宇宙的二重性相应。讲到这种“不易”的“变易”方式,在图2‐3‐7中,是以一种三角形形式出现的,而且是从△1+5=6的矢量加法三角形带的头。而狄德罗(Denis Disetot)曾经认为“物质一定是有感觉的。组成一块石头的分子在积极地寻求某种结合而拒绝某种结合。因而它们的喜好和厌恶是受支配的……于是整个有机体的感觉就是其各部分的感觉的总和,正像一群蜜蜂,出于这只蜜蜂和那只蜜蜂间相互作用的结果,从整体上看,有着团结一致的行为。人所具有的灵魂并不比蜜蜂具有的多”。[4]
在古希腊的理论中,认为“1”是数的产生者,2,3和4则产生线、面和体。认为这是神圣之物,由此并提出一个神圣三角,如图2‐3‐9所示。

图2‐3‐9
而柏拉图则把三角形看成最简单的平面图形。并把它看作基本元素构建五种凸多面体。在我们这里,如图2‐3‐7(E)和图2‐3‐7(F)所示,构成的空间结构就越来越复杂了,表现在矢量加法三角形的个数上,除了图2‐3‐7(D)所示的四个外,又有了图2‐3‐7(E)所示的△1+3=4,△2+3=5,△2+7=9以至于更多的不胜枚举的如图2‐3‐7(F)所示的三角形的聚合了。说明生命的起源有一个自组织过程;也是一种自复制过程,从而表现出一种累加现象。这就应验了禅宗的一句偈语:“万物源自比特(the it from bit)。”说明我们所探索的系统越小,所能发现的原理越多。早在2500年以前的老子就曾说过:“道生一,一生三,二生三,三生万物。”笔者就曾用1、2、3三个数,构成一个“△1+2=3”的矢量加法形,对夸克束缚态以至引力子和光子等作出定量、定性的描述,但在这里无法展开。
至于自组织原理在生物学上的理论意义。美国宾夕法尼亚大学的生物化学家考夫曼(Stuart Kauffman)曾说:“自达尔文以后,有机体一直被看作由随机突变经自然选择而产生的一种特定新玩意儿,然后达尔文并不知道什么是自组织。”[5]。
回头再看看图3‐2-1的意义。除了已如上述的生成论说法已经由图中的数量表示作出印证之外,我们还可看到一种催化关系。比如图中第一个A→T的配对关系的意思是核苷酸T(胸腺嘧啶)要由倍于它的能量的核苷酸A(腺嘌呤)才能激发它使之产生催化效应。而同样的A→T的配对关系,但是处在9→4的档位上,意思是这里的T要由9倍于T的能量去激发它进而产生催化效应。参照资料如图2‐3‐10所示。
图2‐3‐10
这也就是说,在图2‐3‐2中,除了用几何对称的形式对DNA双螺线结构作出恰如的描述以外,还有对催化关系,甚至当量关系也作出相应的表示了。而当量关系的主要单义在于把化学、热学、电学、磁学和生命学之间的联系看作是一种“转换”而总能量保持不变,这种不变的数字表示在我们这里就是一个恒定的结构常数“5”。它在说明生命起源的图2‐3‐2中是“5”;用以描述宇宙大统一的(见图2‐3‐10)也是“5”:
|6-1|=|7-2|=|8-3|=|9-4|=5。
至此,我们发现图3‐2‐2所示的碱基配对关系并非如上图3‐2‐1所描述的那样划一不二的以大促小。这就值得用《洛书》的结构原理来作出某种说明,也就是说,《洛书》机制观照下的图2‐3‐6,究竟存在什么样的花样,详见图2‐3‐11。
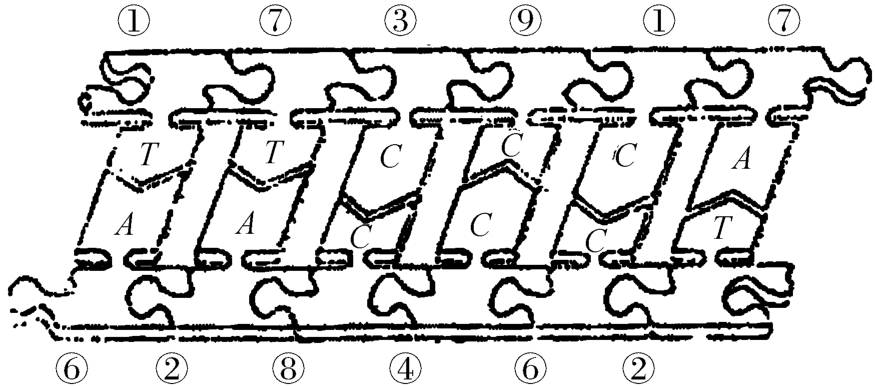
图2‐3‐11
这也就是说:图2‐3‐6所示的纯粹表示为“插头”、“插座”的拼版图,一经加上《洛书》式的结构原理,那么这里的A-T、G-C的关系就获得了一种定性、定量的分析了,比如本图左起的A与T的意义,就可解读为胸腺嘧啶T,要有6倍于T的浓度的腺嘌呤A才能使之催化,这种催化是一种自催化,接着的第二项:7→2相对于A→T的关系,却相反了,好像是说T的存在,在这里不接受A的催化,而为一种自阻化。但也正因如此,成就了一种“交叉催化。”意思是说,属于两个不同反应链的两种产物各自促进对方的合成。中国的语言叫“相反相成”。
这似乎就是《洛书》矩阵的结构原理,既是守恒的,又是变易的,所以是能动的、发展的,不然,生命现象就不可能如此五彩缤纷。
由此,我们不妨对很长的DNA双螺线试作《洛书》式的统计学处理,即以本图2‐3‐13而言,它的数码表示如图2‐3‐12所示。
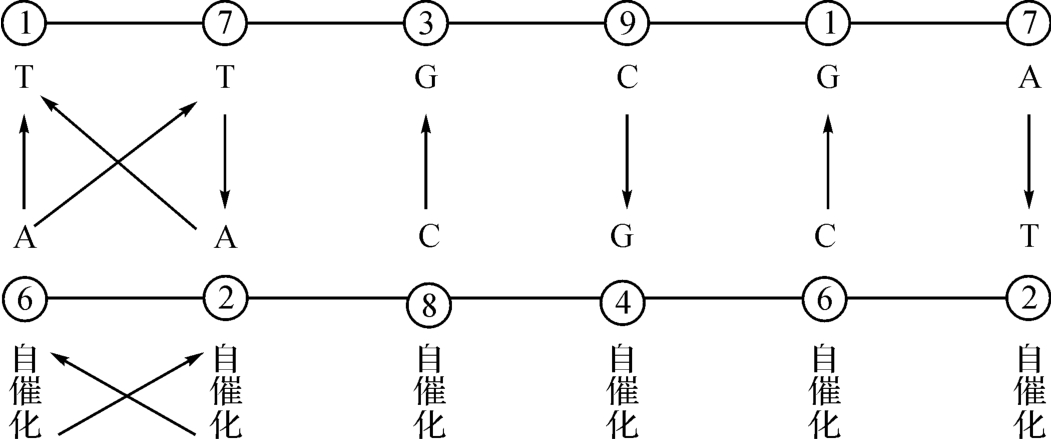
图2‐3‐12
一、《洛书》式的统计学处理
这就是说,面对很长的催化链的字母表示,对于其中的催化关系究竟是自催化还是自阻化,以至是否存在交叉催化,是很难一眼看出的。但若借助自然自组织的《洛书》式的规范对称,即可审定它们的催化性质了。而同时,还可把很长的催化链分成若干个组成单位,如图2‐3‐13所示。
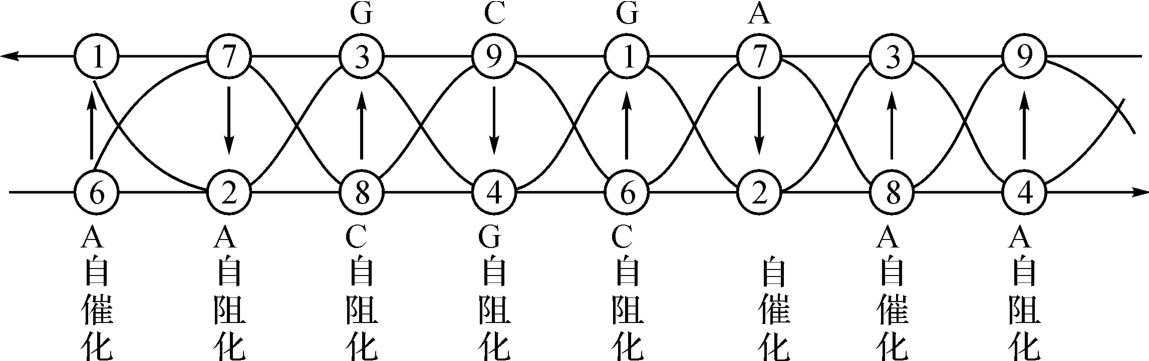
图2‐3‐13
由此可以认为:基因通过DNA分子产生RNA分子的转录过程是确定的。但是由RNA分子指导蛋白质全盛的翻译过程却是不确定的,是多种多样的。说明反应的模式是简单的,但分子(蛋白质、核酸等)是极为复杂的、特殊的。生物学和物理学的区别在这里表现为生物学向复杂方向发展,而物理学则向基本的方向发展。
联系前述的讨论,似可认为生物学的这种复杂性,表现在图2‐3‐13之中,即使是任意的设定所表示的序列是确定的,但在这种确定的序列中却又存在一种套盒机构,可以分为核心(信息)部分。由于我们对64个密码子的编号方式与64个重卦相对应,而64卦是双层结构,比如第一个密码子CCC处于64卦之首。而64卦之首的卦象,所以作如下定量表述:
诸如此类,详见图2‐3‐14和2‐3‐15。
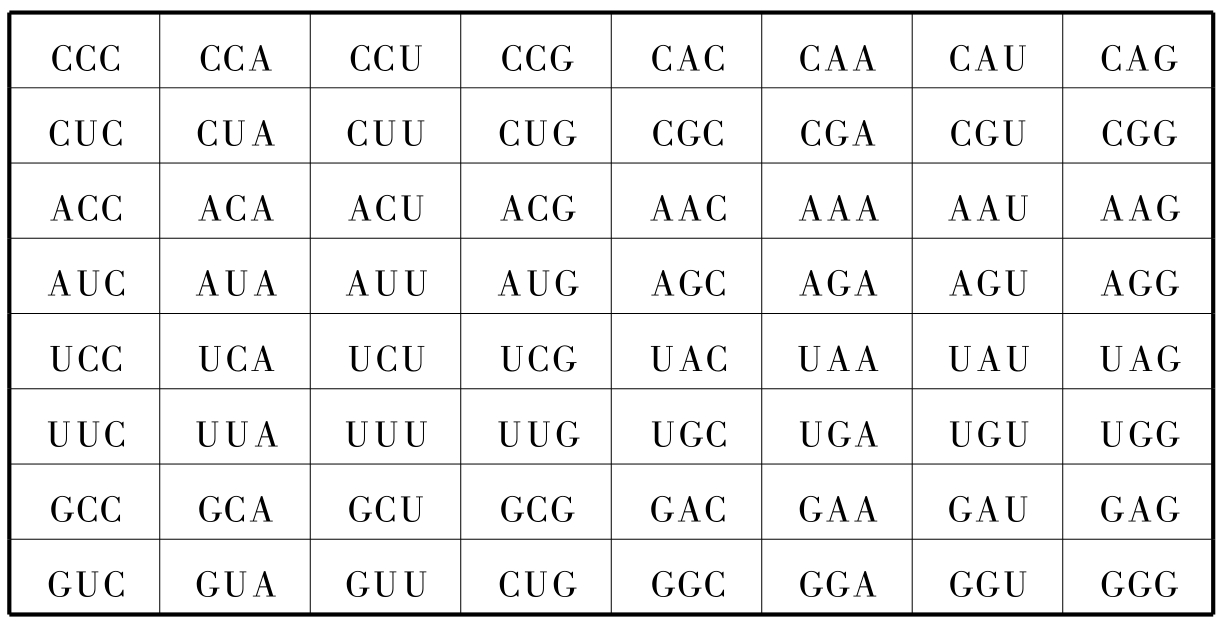
图2‐3‐14

图2‐3‐15
二、64遗传密码与64八卦接轨
这种编码方式对于64个密码子来说,有一个好处,就是对横向的相干性可作定量表示,如图2‐3‐15所示。
此外,还有一个催化效应逐级放大问题,这就要有个动力机制。那就应该到64个遗传密码中去寻长它自身的动力机制。至此,我们还得先说说遗传密码A、U、G、C。按照中国的传统讲法叫四象,而与四维时—空(三维空间加一维时间)相对应。因此,这四种密码必须4×4=16,这又和八卦的64卦之数相合了。不然,如果空间是2维,那么对于任何复杂的有机体是不够用的;超过四维而为五维时空也不行,不然绕原子核旋转的电子的轨道就不稳定,会以螺旋的轨道向中心趋近(S.霍金)。
根据这种法则要求,我们发现64个遗传密码的核心部位是四个密码子:AUG、AGC、UCG、CAG,如图2‐3‐16所示。

图2‐3‐16
这里的所谓相干性,从分维表示看,8个密码子的分维数的“分母”都是1。而它的分子却构成为一个以“5”为结构常数的奇—偶互补对称的整体了。这种整体性的存在形式只是说明这里是一个“数学场,”而并非说明这里存在一种A‐T,G‐C的催化关系。
在理论上,我们知道,物质世界具有某种层次性表现,如图2‐3‐16所示,它和横向的相干性构成一个总体。也就是说,这种层次性结构是由若干要素经横向的相干性而构成的。换言之,正因系统中的横向相干性的存在,才导致纵向层次间质的差异。
而在图2‐3‐15的纵向层次性中,它的表现形式仍然有另一种奇—偶互补关系存在。比如图2‐3‐15左侧第一行的密码子的数量表示,从上于是上,把它转换为由左到右,也仍然和图2‐3‐16那样以“5”为结构常数所作出的奇—偶互补对称组合,如图2‐3‐17所示。
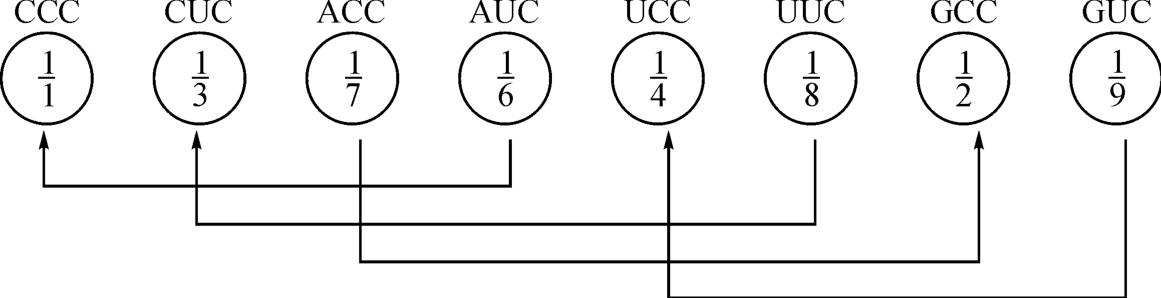
图2‐3‐17
联系我们这里的讨论主题,是要从图2‐3‐15中找到某种动力机制,以便让我们看到看似分散的64个遗传密码自身必然也存在着一种自组织动力,变静态的遗传密码为动态的东西,进而产生某种催化关系,如图2‐3‐18所示。
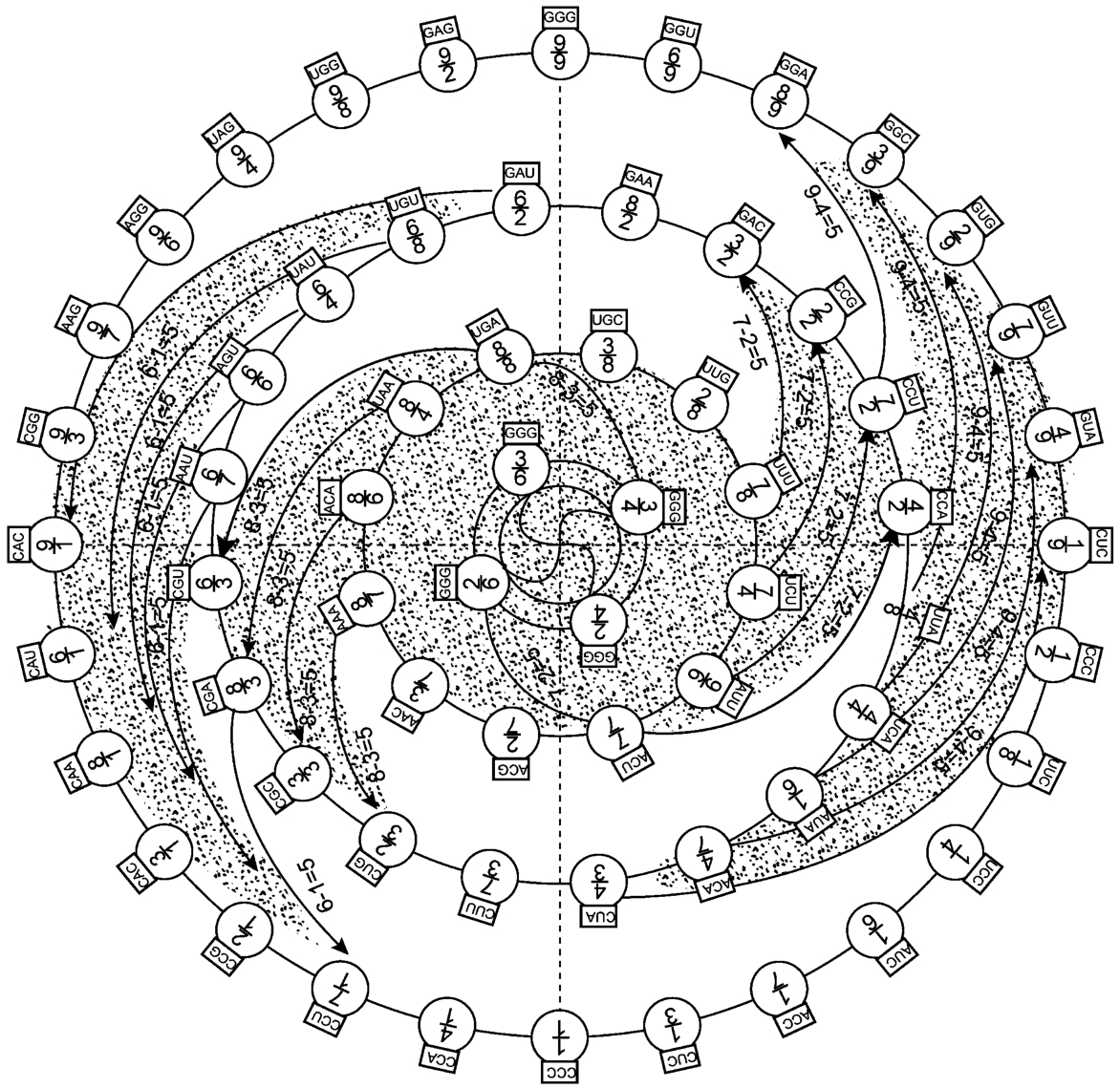
图2‐3‐18
三、DNA数字之流
由此,我们可以认为图3‐5‐1所示的用英文字母表述的遗传密码,在《洛书》、《八卦》的结构原理的观照下,显示出了离散有64个密码子,实际上存在一种自组织原理,本然地有一种旋涡动力机制在其中。而这一点,也似乎只有具自然自发自组织原理的《洛书》矩阵才能做到。
④64八卦中有一个动态的华达哥拉斯定理。量子色动力学(QCD)与八卦接轨。
资料告诉我们,遗传物质虽然只有A、U、G、C四种,但在地球历史上,在DNA之流上,曾经有过3000万条支流。而据估计,现在地球上所存在的物种只不过曾经存在过的1%。
论述至此,面对图3‐5‐4这样的空间形式,笔者联想到西文物理学家有所谓蒙特卡洛法(Monte Carlo Method)简易量子色动力学(QCD)计算的新技术,将会告诉人们许多早期的宇宙情况。
这种计算的数学框架为每边8个点的网络。由于我们的时空是四维的,所以网络总共包含84=4096个点。因此,这样的网络作为一个矩阵就包含4096×4096,大约1600万个数据。
理论认为,粒子相互作用所处的宇宙的四维时空首先被划分为不连续的网格。某一程序能够记录下网格中每个点的费米子,在这种程序中,把费米子聚集起来的规范场被当作网络中连接各点的连线。这种程序考虑的是网络的各种可能的构型出现的概率,并根据它们的概率加权。
笔者之所以在这里插述这种新技术的用意,在于说明这种每边8个点的矩阵的实质竟是我国上古早已有之的64卦框架(见图2‐3‐15)。而且,西文的物理学家为什么要考虑使用这样的点阵的原因是由于考虑到一个严谨的规则即泡利不相容原理。因为宇宙中不允许存在2个同样状态的费米子。任何2个同类的费米子,要么位置不同,要么其他量子数不同。这样一来,就无疑要给物理学家增计算上的麻烦,所以设想这种每边8个点的矩阵,可以获得1600万个数据而避开泡利不相容原理[6]。而在图2‐3‐15中,就无相同的卦象同在一个方格内。
这个每边8个点的点阵还隐藏着一个毕达哥拉斯定理,如图2‐3‐19所示。

图2‐3‐19
1820年,德国著名数学家高斯(C.F.Gauss)曾建议在西伯利亚的原始森林中伐出一片直角三角形的空地种植小麦,再在三角形的边上各种一片正方形的松树,以此表达毕达哥拉斯定理,让途径太阳系的外星人一眼可以断定,地球上存在智慧生物,可遗憾的是高斯的几何形设计未能像本图2‐3‐19这样作出定量表述。
这是一个变维空间(Ernst Mach),它表现为82≠72,但有一个共同的耦合常数“5”在其中。根据这个数学框架,可以作出图2‐3‐20所示的趋向无穷的数量运动。

图2‐3‐20
如图2‐3‐20所示,由色三股四弦五而展开的一个平滑连续筒上,出现了以“92=81”和“42=16”为指数的数值运动,一方面向无穷大迈进,同时也表现为以这样的指数作周期叠加于原点,意味着这是一种引力坍缩,其结果必须会出现曲率无限而高密、高温的奇点,然后逆转而为时空膨胀。从而可以认为热大爆炸前宇宙已经存在,而与2000年6月3日英国《新科学家》周刊所发表的,根据欧洲粒子研究中心(CERN)的加布里埃尔·维尼齐亚诺(Gabrelle Veneziano)的理论而发表的“欧洲学者提出新理论——大爆炸以前宇宙已经存在”的文章的论点相合。
笔者又复从图2‐3‐20中只提取其中弦五一项而作出宇宙旋涡图,如图2‐3‐21所示。
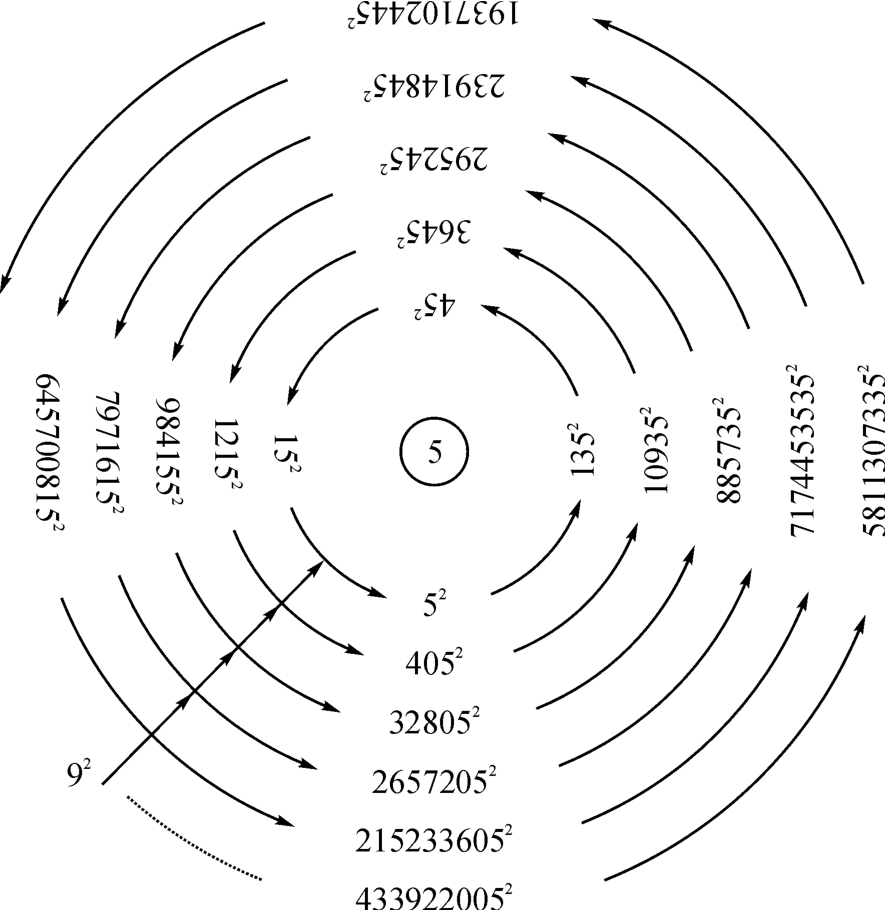
图2‐3‐21
此图结构原理和第二篇图2‐2‐11相同,银河系模型(俯视)如图2‐3‐22所示。

图2‐3‐22
至此,我们的讨论好像在随心所欲地侃谈,把生命起源拉扯到宇宙起源。第一,我们所引据的数学模式相同,第二,DNA既可说明生命的起源,也可以说明生命的复制。在宇宙论上,在我的理论中,类星体就是一种复制宇宙的样本,问题是这里无法展开,而俄罗斯物理学家安德烈·林德(Andrei Linde)的宇宙论,就被称为混沌的、分形的、永远自复制的暴涨宇宙论。
四、遗传密码QCD计算得出的构型
至于前述关于QCD计算中可能出现的各种构型,要是在遗传密码中,就可接着提出如下的一个图(见图2‐3‐23):
这只能说是一种十分简单的数学游戏,却使文字表述的密码子形成一种直观的几何图像,当然没有操作人员把各个分子进行配对。在A、C凯恩斯的书中所说的那样,各种各样的连接器和其他分子一个接着一个与空着的插座相撞。最后一个载有核苷酸的CCC终于来到,咔哒一声插在GGG里面。

图2‐3‐23
有人说数论只能用高等数学推导,而不能用初等数学证明,而在我们这里,这种十分简单的算术操作的实践价值却可用十分优美的、直观的空间结构作出证明;并且启示我们生命世界是一个双层结构的世界,在S.霍金的理论中即为“膜的世界”。
至此,我们要问所谓外显子和内含子的理论是否与这里的双层结构有关呢?再看图2‐3‐24。
本图所示遗传因子A‐U,G‐C总数64,G、C、A、U各16个,它们的出处来自图3‐4‐4中每个遗传密码中的中间这个字母。如图2‐3‐24所示的关系,是以《洛书》结构原理|6-1|=|7-2|=|8-3|=|9-4|=5来分的。
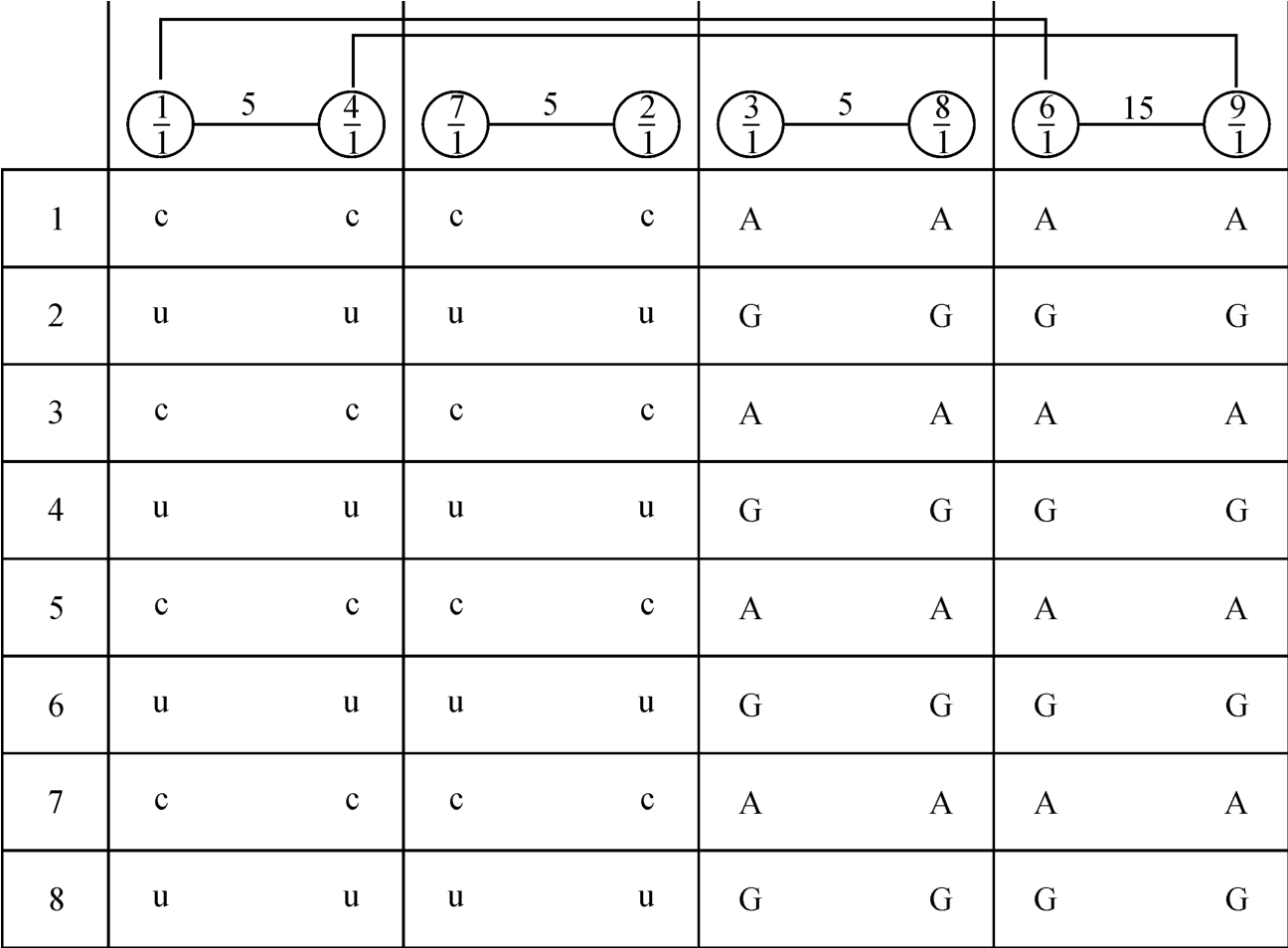
图2‐3‐24
如果联系如图2‐3‐9所示的神圣三角的音律意义,即利用这四个数可以表示三个基本和谐音:4/3,3/2,2/1和一个双八度的和谐音4/1。又假如把这个神圣三角的数变换1~4的数轴而定为图2‐3‐28这样的关系,那么“1+4=5”和“2+3=5”相协,如图2‐3‐25所示。
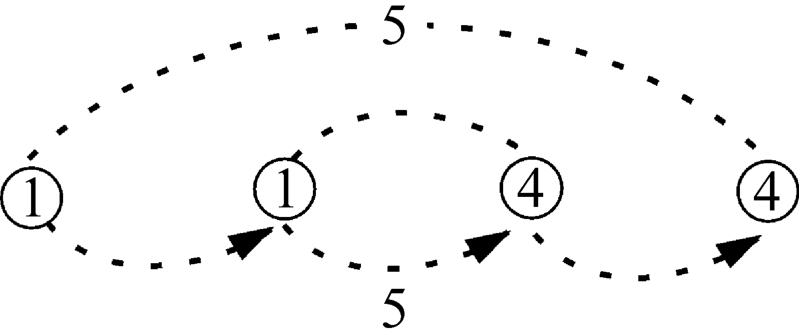
图2‐3‐25
照此说来,我们的DNA是否也存在一种外象—内象关系与前述的外显子—内含子相对应呢(见图2‐3‐26)?
那也就是说,DNA为什么会构成双螺线结构的原因是否也就是这里呢?说明如图2‐3‐26所示的静态密码子,却是存在着变动不居的玄机了(见图2‐3‐27)。
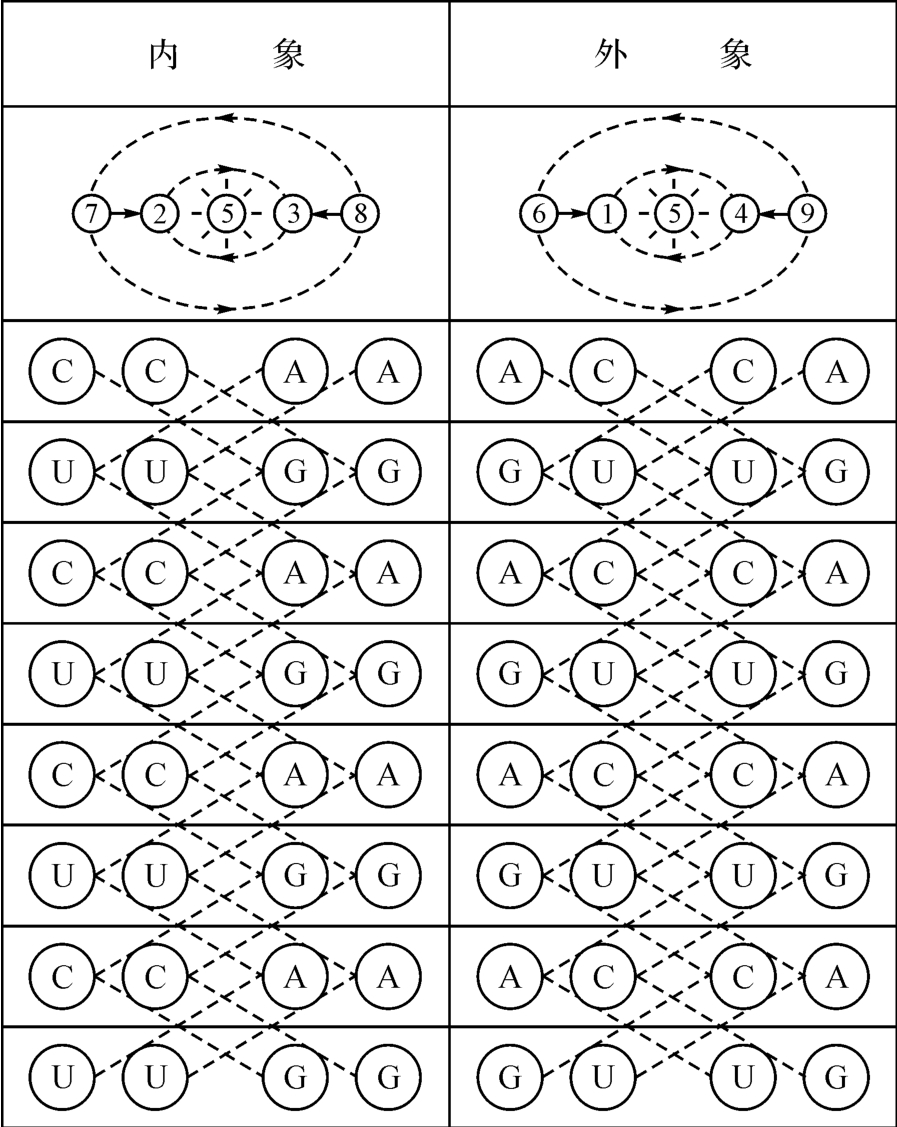
图2‐3‐26

图2‐3‐27
五、雏菊状64遗传密码
虽然,图2‐3‐18的层次性,那种矩阵构型被转换为这里像雏菊一样的放射形,由内而外分为4—12—24—28好像是4季—12月—28宿—俨然中国古天文的赤极坐标线的味道。在这里,任何相对两个遗传密码也照样获得A—U和G—C的互补配对绝无例外。它告诉人们:地球上生命形态的多样性主要存在于表面。越往里面,多样性愈少。在中心,多样性就不存在。鲸与蝙蝠之间,它们在设计上的雷同,可能大大超出它们的外表所显出的不同,说明地球上的生命出自一个共同的祖先(A.G.凯恩斯)。卡尔·萨根则风趣地说:“橡树和我竟是用相同的材料制成?”
F.Bacon说:“一切绝妙的美都显示出奇异的均衡关系。”那么我要说,自然乃是“均衡数学”的鼻祖。也就是说,只有自然自组织的《洛书》数理才能作出如此绝妙的“均衡关系,”但在均衡中有自发破缺。
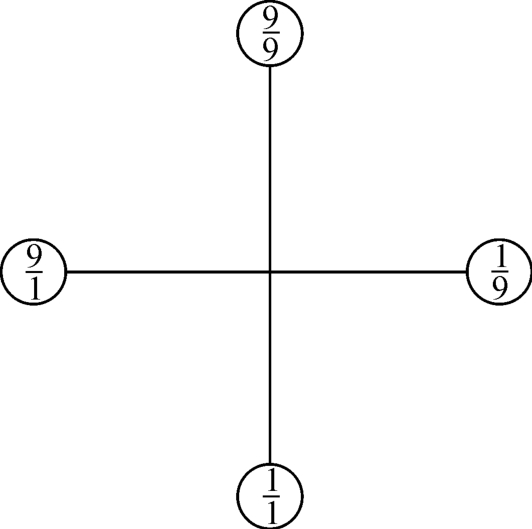
图2‐3‐28
六、基因的断裂
在图2‐3‐28中,又有一个直角坐标。
要横坐标左右两端的基因表示,可能认为这里出现了基因断裂。CAGGUC资料告诉我们:“断裂基因的剪接问题有一个GU—AG法则”,如图2‐3‐29所示。
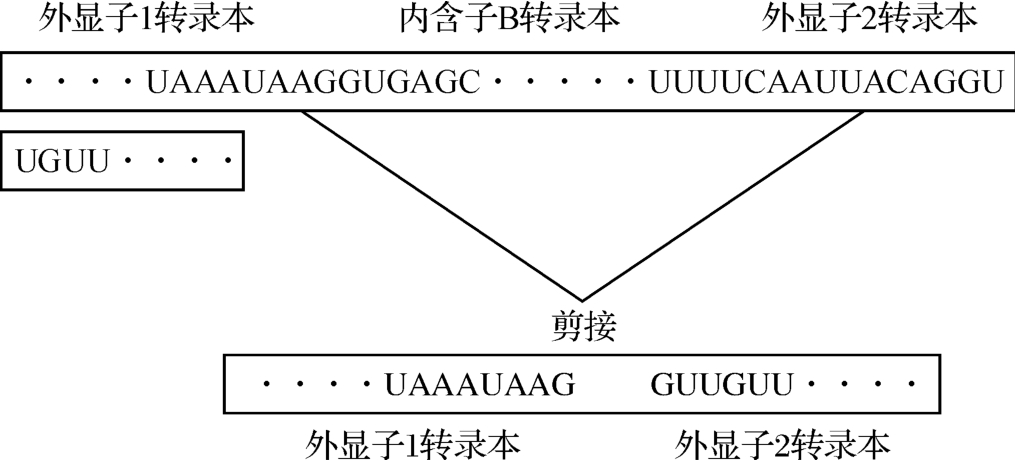
图2‐3‐29
这就惊人地说明《洛书》矩阵具有明察秋毫的数学创造力,因为断裂基因的发现,在时间上是20世纪70年代利用电子显微镜才得以发现的;而其存在的环境,那简直就像在一片迷雾或者落花流水之中,如图2‐3‐30所示。
图2‐3‐30
这就怪不得,这一发现被认为是改变了我们对高等生物基因在进化中是如何发展的观点,从而对生物学的基础研究有着十分重要的意义。因此这项发现在1993年获得了诺贝尔化学奖,获此殊荣的是英国遗传学家理查德·罗伯茨和美国遗传学家菲利普·夏普。而其实,中国的学者周芷,早在1970年就从她的研究中已经发现DNA转录为RNA时,少了一段遗传密码,推论出基因可能断裂的假说,并在她后来的实验中证实了这一点。而罗伯茨那时正与周芷同在美国冷泉港实验室工作,1977年发表论文时,只好共同署名。1933年11月12日中国中医学报为此发表了题为《今年诺贝尔医学奖》的文章,令人不解——最先发现断裂基因的周芷竟榜上无名,这自然是笔者带便的话。
七、S.考夫曼的生命自组织理论
斯图亚特·考夫曼(Stuart Kaufman),美国宾夕法亚大学生物化学家,1993年出版了一本巨著《秩序的起源:进化中的自组织与自然选择》[7],认为达尔文不知道自组织理念,在达尔文的一生中,对孟德尔(Gregor Mende)的研究,一直懵无所知。
1990年2月5日在美国新墨西哥州的圣菲(SantaFe)召开的第二次人工模拟生命专题讨论会上,S.考夫曼指出:人的每个细胞中包含有大约十万个基因。其中包含数量不详的调节基因。它们相互之间的开和关都是通过一个巨大的、复杂的相互作用网来实现的。看起来似乎一片混乱,因为很多人认为如此复杂的系统应该是始终如一地循着同一个方向循环进行的,然后,事实并非如此,也许除癌细胞以外,所有的基因对应于特定的细胞类型通过自组织而形成其稳定的运动模式,例如,一些基因的活动对应于白血球,另一些基因的活动则对于平滑的肌细胞等等。那么,这些活动的模式是怎样形成的呢?
……
考夫曼惊人地发现:一个稳定细胞类型的形成,并不是进化过程中的一个偶然事件。在他模拟的基因中,与具有自催化特性的、稳定的基因活动模式相对应的细胞类型就相当于数学家们所称的“吸引子”利用数学中的动力学系统理论。他并且已经证明不管系统网络开始时是如何的无组织,这种模式的形成,几乎是不可逆的。简而言之,似首基因间相互作用的原动力可促使基因组织产生某种形式自发性的自组织。
这种自发性的自组织运动,在考夫曼的理论中,他建立一个数学模型把遗传调节系统理想化为一个纯粹的相互作用网:基因A开启基因B;基因B开启基因C而抑制基因D的表达;等等。
笔者把他的话再重复一句:这里主要阐发了这样一点,在细胞中有这样一种动力模式:某一种稳定的运动模式,是受控于数量不详的调节基因的。它们相互之间的开和关都是通过一个巨大、复杂的相互作用网来实现的。
作为一个读者,遗憾的是,考夫曼所建立的这种理想化的相互作用网,笔者至今未能看到,也许早已有了,但我尚未看到。而按我的设想,他所说的一个相互作用网,它的控制中心,就由密码子UCG—AGC,AUG—UAC联合组成,如图2‐3‐31所示。
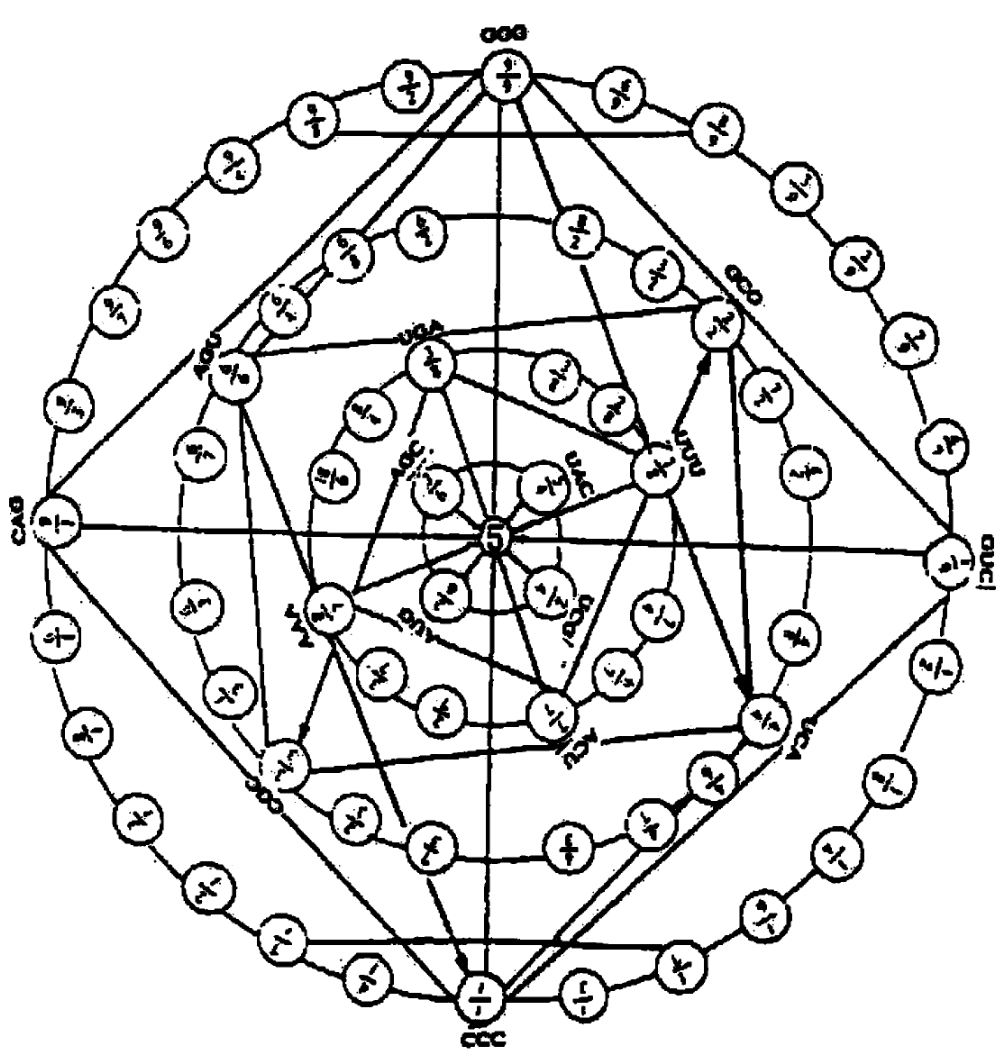
图2‐3‐31
八、昏迷77年醒后青春依旧
1994年美国《太阳报》曾经报道一个耸人听闻的奇事:蒙古公主米莉嘉昏睡了77年后,不久前一朝醒来,仍保持当年的25岁的青春娇美。但发现全世界已发生了翻天覆地的变化。为了追赶时代,她正马不停蹄地到世界各地旅游。
1917年12月3日,她为了逃避俄国大革命的滚滚洪流,策马南下时坠马受伤,从此,一直昏迷,到去年底国际学研究小组精英高手,通过医艺妙术才使她苏醒过来。
从生理年龄说,这位蒙古公主是102岁,但肉体却仍和77年前25岁一样年轻漂亮。
医生指出,米莉嘉一睡70多年,主要由于她坠马时碰撞了头部,使她沉沉昏睡外,还引致大脑产生一种化学的反应,终止了她的老化进程。
后来一份杂志披露了米莉嘉苏醒的消息,据称她一直在莫斯科克里姆林宫一所秘密医院的病房里。70多年来,日夜有医护人员治疗和照料。去年秋天,在美国、德国和澳大利亚脑科专家的努力下,她终于恢复了神经上的联系,苏醒过来,据医生说,米莉嘉坠马被碰撞昏迷时,其“死亡基因”的大脑部门功能,也一并终止。
当时她是蒙古王室中的成员,出事时正出访莫斯科,却碰上了“十月革命”。苏联认为她是成吉思汗后裔,怕事件国际化,于是把她救起,但却一直否认知道公主的下落。
直到苏联解体后,俄罗斯才承认不为岁月所动的米莉嘉,已在克里姆林宫接受治疗77年。当医生批准米莉嘉出外旅行时,她首先返回乌兰巴托,但只逗留了几个星期,便开始她的环游世界。她已先后到过法国南部和爱琴海温泉,也畅游了美国迪斯尼世界。
话已说得十分清楚,故事主人在坠马受伤时,死亡基因的大脑活动即已停止,就是说,衰老的活动模式的开关关上了。
接着我们不妨把上述四个密码子的物质性质作出某种剖析,如图2‐3‐32所示。
这可能是闻所未闻的做法,它需要加以如下说明:
这里的四个遗传密码:AUG、AGC、UCG和UAC就是图2‐3‐15,亦即2‐3‐31中心的四个密码子。
每个密码子都有一个分维符合,比如AUG的分维符号为(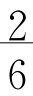 )。
)。
已知:上夸克电荷为2/3,下夸克电荷为-1/3。
又知2个上(u)夸克和1个下(d)夸克相结合而为1个质子:P=uud,它的电荷为1=2/3+2/3+-1/3。
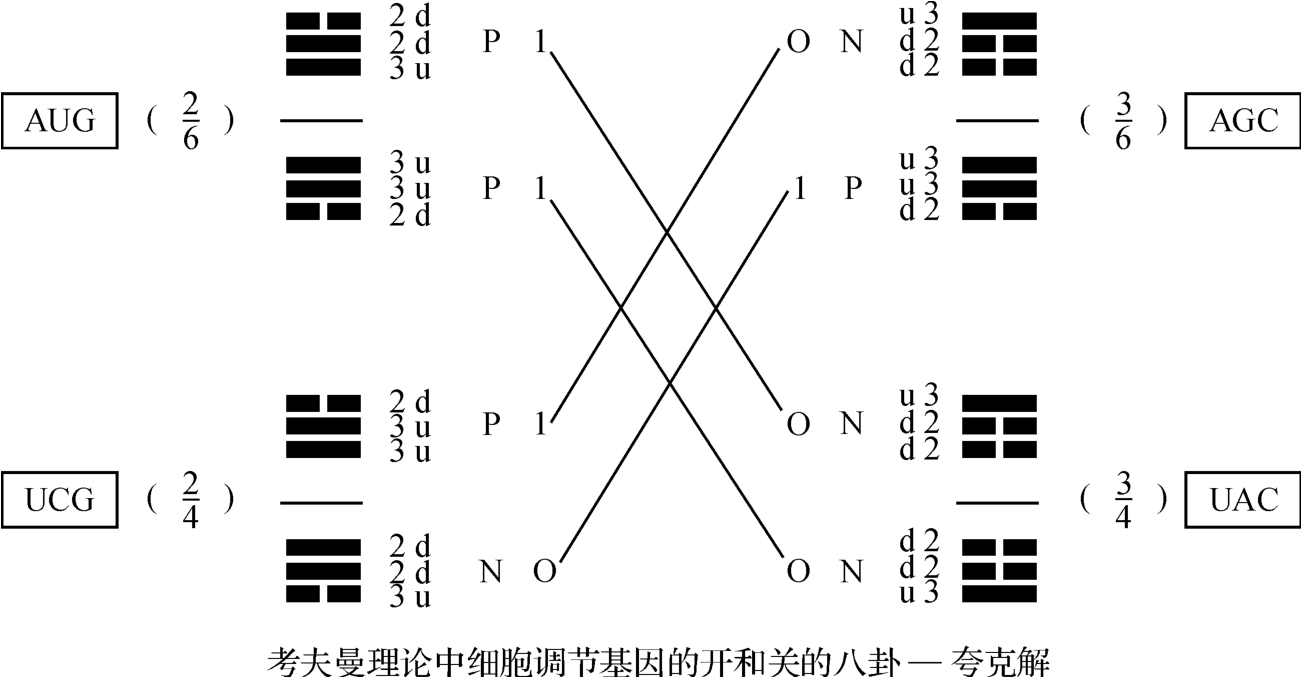
图2‐3‐32
又知2个下(d)夸克和1个上(u)夸克相结合而为1个中子N=ddu,它的电荷为0=-1/3+-1/3+2/3。
八卦和夸克具同构性,主要在于:
阳爻 (3)和上(u)夸克同构。
(3)和上(u)夸克同构。
阴爻 (2)和下(d)夸克同构。
(2)和下(d)夸克同构。
根据以上说明,图2‐3‐32可以化简为图2‐3‐33。
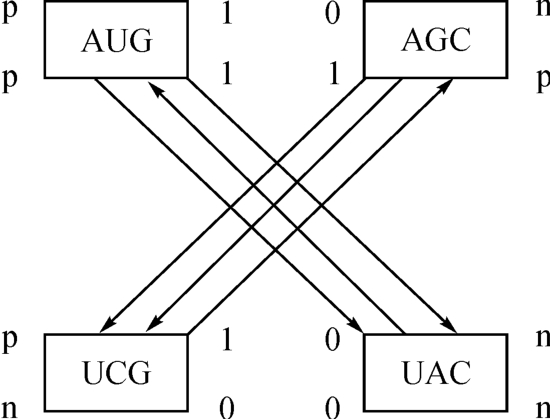
图2‐3‐33
这也就是说,图2‐3‐32的中心,确是一个由调节基因组成的调控中心,其间有“开—关”装置,这种装置的开启,会使一种稳定的活动模式持续下去。如果相反而为关闭,那么相关的活动模式也就停止不再进行。上述故事的女主人,犯的毛病是“死亡基因”被关上了,稳定的生命衰老的活动模式停顿了77年。这种病,生得好,77年后她又活过来了,而且青春依旧;并且躲过了一场战乱。展望将来,医药科学更加发达,人的年寿长短,或可由自己来设定了。俄罗斯《论据与事实》周报题为《100年后醒来》的文章就是讲一种冷冻法的探索。
那么,能够调控生命活性的物质是什么呢?据悉,磷元素就具有调控生命分子活性的功能。蛋白质最小分子的基因转移、信息转移都是受磷调控的。
在考夫曼的自组织生命理论中并认为自催化系统可以形成的条件是足够的多样性。例如,系统中所有分子都具有相同的催化强度时,形成的较长聚合物的浓度就相当低,而当系统中分子具有不同的催化强度时,形成的长链分子就较多。对于这一点,笔者认为同时并存在一个动力机制问题。
这种动力机制,从《洛书》矩阵看来是自洽自足的,如图2‐3‐33所示。它既能作相对内旋(旋紧),又可作相对外旋(旋松)。
下面,先按《洛书》结构原理的要求,把图2‐3‐22中的8个密码子纳入一条《洛书》式的DNA双螺线结构中,如图2‐3‐34所示。
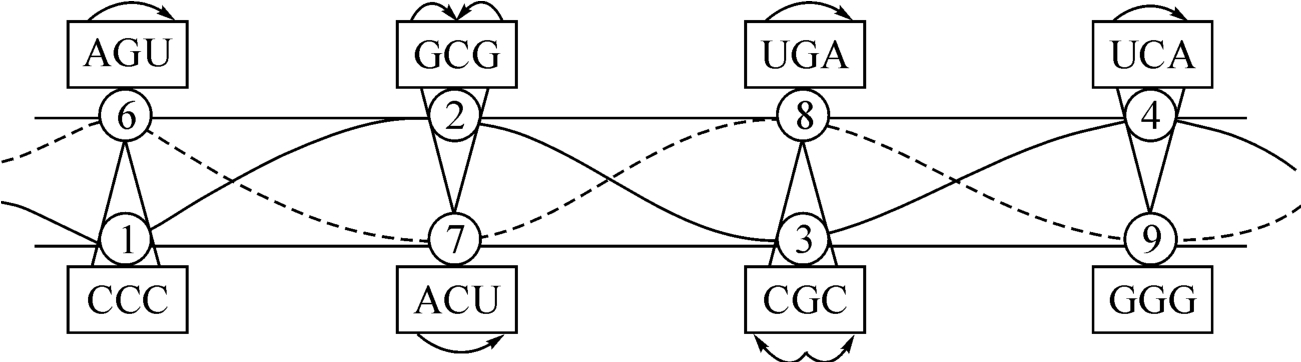
图2‐3‐34
图2‐3‐33,好像一张充满对称的装饰画。首先表现在整体上,它是一个双层结构。其间有两条由6-7-8-9和1-2-3-4互相缠绕的双螺线。在画的左右两端,左边遗传密码子AGU,右为密码子UCA,两者各有一个A催化U的关系,而方向像互相叩头。其间的距离为6+4=10,与此同时,双层结构的底层,左为CCC三个细胞嘧啶组成的密码子,和右端的GGG,即三个鸟嘌呤组成的密码子互相对待。好像CCC会走路,前来和GGG组装在一起(见图2‐3‐6)。
单是上述左右两端的结构关系的数学实质,就是电磁力数学场U(1)和引力数学场的一左一右关系。由此可以结合说明这里存在一种内含子的关系如AGU,同时又存在一种外显子的关系如图2‐3‐34左端的 ,一个好像是散投奔,一个好像是激光的聚合,在数量结构上,6—1与9—4的关系又正是互补统一的,如图2‐3‐35所示。
,一个好像是散投奔,一个好像是激光的聚合,在数量结构上,6—1与9—4的关系又正是互补统一的,如图2‐3‐35所示。
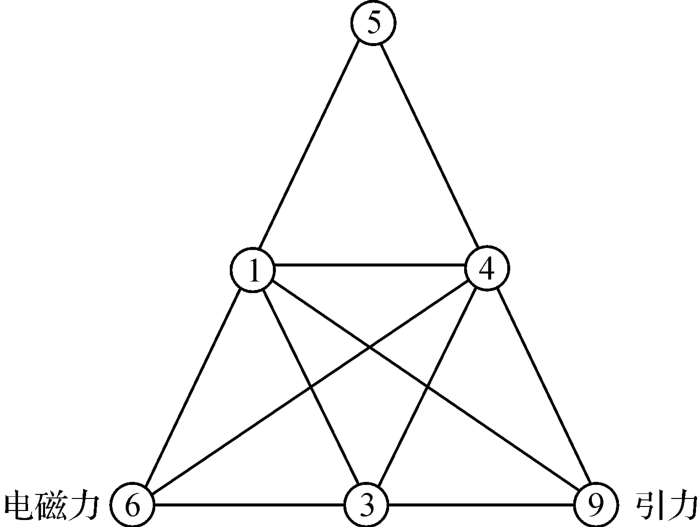
图2‐3‐35
此外,我们还在图2‐3‐34的四个密码子上看到同样有超弦的对称关系。密码子GCG相对于密码子CGC,如果把前者看成两种力向中心集中,那么,后者表示中心向左右发散,前者为阖。“往来不究,谓之通。”(《周易·上传》)跟这两个密码子结对的AGU和UGA,彼此表示:前者的AU和后者的U—A好像两对互相揖让作礼,十分友好。
如果也照弱力场和强力场来理解,那么,这两种力,也是互补统一的,如图2‐3‐36所示。
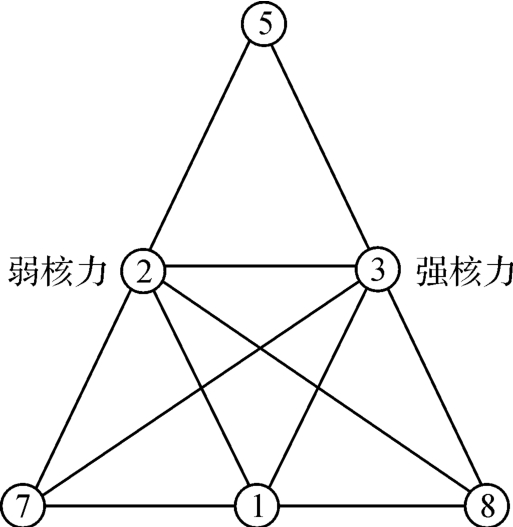
图2‐3‐36
以上所论,可能是故弄玄虚,夸夸其谈,但有一点令人百思不得其解:为什么我们所看到的包括人类在内的一切生物,会构造得如此巧妙、动人?这里头存在什么样的动力机制?如果离开了四种自然力(电磁力、引力、强力和弱力)难道另有一种万能的力在起作用吗?理论又告诉我们,一切物质的最小单位叫夸克。而夸克又被说成是纯粹的数学构造,离开了它的数学定义,便没有任何意义,这种讲法又何尝不是玄虚之谈吗?因为在实际上,大自然的所作所为要比人类能够想象到的美妙还来得美妙。
下面,我们就把图2‐3‐34变换成一个环球形,对前述催化系统作出逐级放大的表示,如图2‐3‐37所示。
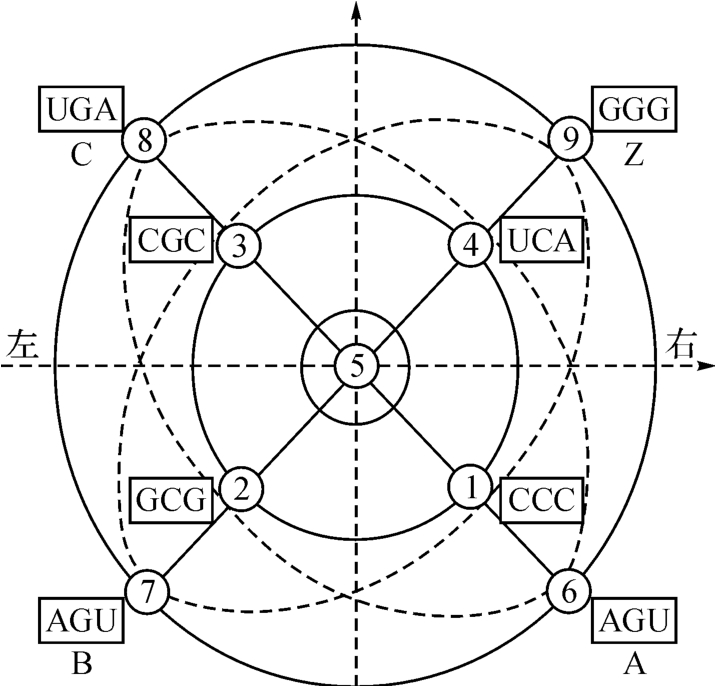
图2‐3‐37
之所以可以这样变,是因为图2‐3‐34存在着一致的波长“2”,即首尾是相连的,所以它的波长是量子化的:
|4-2|=|8-6|=|9-7|=|3-1|=2。
原来,在考夫曼的理论中,他曾说从一般的实验观察中得到启示,氨基酸和其他小分子化合物混在一起偶尔会自发地连接成一些短的聚合分子,而且有很多小聚合分子对其他反应具有弱的催化作用,因此假设聚合物A催化另一种分子B的形成,而B又可催化另一种反应生成C等。依次类推产生的终产物Z又可催化A的产生,那么就自然形成了可称之为自催化系统的一组逐级放大的反应系列。此类系统一旦形成,它即具有类似于原始代谢的功能。系统即可从氨基酸和其他简单化学分子中获得比较稳定的“食物”及能量供给……考夫曼的合作者(这次人工模拟生命专题讨论会的组织者之一)法默(DoyneFarmer)还认为生命起源与新陈代谢起源很可能是同一回事。
为了充分说明这种逐级放大的反应机理,我们再引述由国家教委社会科学研究与思想政治工作司组编、供理工农医类硕士研究生使用的高等学校教材《自然辩证法概论》[8]一书的图式,如图2‐3‐38所示。
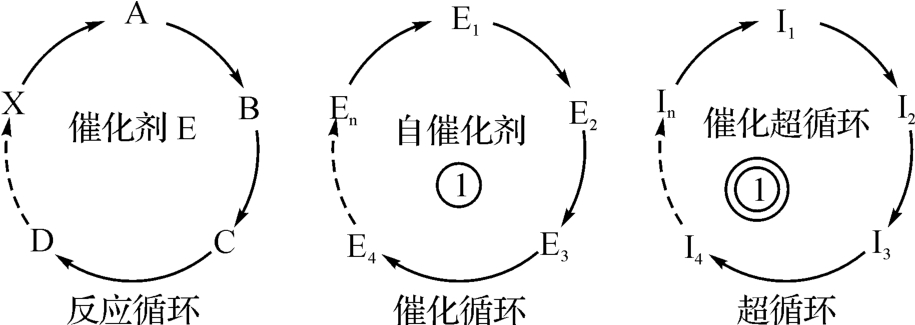
图2‐3‐38
文章说:反应循环虽不改变最后生成物的性能,但却改变了反应的速度,这对于许多化学反应都是重要的。通常不易发生的化学反应在反应循环建立之后就可以进行。在这个意义上可以认为,反应循环的组织形式提供了物理运动向化学运动转化的一个条件。
催化循环可以造成亲代和子代之间的“通信”,使一定量的信息在逐代传递中稳定不变。超循环更进一步,不仅能使单元信息保持不变,而且能使不同的功能单元的联合稳定保持下来。这对于生命运动是极端重要的。无论简单或复杂的生命系统,都是功能单元所结合起来的有机体。
不难领会,上述图式和引文对于图2‐3‐37的理论意义得到了更多的发挥。比如说,物理运动向化学运动的转化。从而进一步说明《洛书》《八卦》的数学创造力,带有某种普适性。但从本文的主题而言,仅在于生命的起源,如果又要对图2‐3‐4至2‐3‐5所示的四种相互作用力作出统一的论述,那就无法在这里展开了。
而且事实上,在讨论图2‐3‐33所示的遗传密码AUG、AGC、UCG和UAC的物理意义时,运用了夸克的结构原理,从而导出了质子电荷1,中子电荷0而为一个是“开”一个是“关”的控制机理,说明这里的生物学与物理学已经装到一起了。这从图2‐3‐7所示的1~9九个自然数的自发自组织原理来说当是情理之中的事。换句话说,要想推翻它,并不比建立它来得容易。
长铎先生的以上所论,就算是一份答卷,用以回应“普里戈津教授深信用古老的东方《易经》有助于进一步了解自然”这句话。
至于为什么选定生命起源问题作为本文论述的主题,原因在于1984年也即普—刘会晤的同一年,联合国教科文组织(UNESCO)的《信使》杂志11月号发表了[美]著名科学家卡尔·萨根(Carl Sagan)的文章——《生命的奥秘》中,竟用我国2000年前的西汉帛画——伏羲—女娲人身蛇尾作为DNA双螺旋线的参照物。而这一点,却又正好可和普里高津教授以下的话相联系。他说:科学的每个伟大时期,都引出某个自然界的模型,对经典科学来说,这个模型是钟表:对19世纪的科学,即工业革命时代来说,这个模型是一个逐渐慢下来的发动机;对于我们来说,标志可能是什么呢?我们头脑里的东西也许可以用那些从古印度或前哥伦布的艺术直到我们时代为止的雕刻最好地表达出来……
不过我想要提出的我们这个时代的标志不是出自古印度而是我们中国远古即已有之的《洛书》矩阵以及由《洛书》推演而出的八卦,还有已如上述的伏羲—女娲人身蛇尾像(见图2‐3‐1)。
因为21世纪的前沿科学之一是生命科学,那么这张图像也就与DNA双螺线结构相呼应的艺术品了!
这也许是普里高津教授头脑里所设想的东西居然由卡尔·萨根唯独在中国新疆的博物馆里找到了。古语说“心有灵犀一点通”,莫非这就是一个现实的例子。
天地默默,但不是无话与我们人类对话。不过她用的“手语”是自然自组织的数学符合。
我们知道,自组织这个理念,是普里高津理论的核心。他的闪光大作《从混沌到有序》一书所表述的中心思想,如果借用美国未来学家托夫勒(Alvin Toffler)的话来说,就是“有序和组织可以通过一个‘自组织’的过程真的从无序和混沌中‘自发地’产生出来”。因此,《从混沌到有序》这本书,在托夫勒看来,可以具体地说,“就是把生物学和物理学重新装到一起,把必然性和偶然性重新装到一起,把自然科学和人文科学重新装到一起”。
综上所论,考夫曼的生命自组织论,普里戈津的《从混沌走向有序》的理论,以及托夫勒上述未来学理论,好像都在同一条线上跑向中国古老的《洛书》面前,把他们的理论和1~9九个自然数的自发、自组织原理接轨。不然他们的理论就缺少无可置疑的数理基础。
人类性状色体X、Y的交换如图2‐3‐39所示。
已知男性性状色体为X、Y各1;女性性状染色体为2个X;而且,在女性的卵中,已天然地储存着一个X.这也就是说,这个卵将化育为一个男孩,抑或一个女孩.取决于它将从妻子的丈夫的精子中所取得的染色体是X还是Y而时定.如果是X,则为女孩;如果是Y,则为男孩。
这也许是牵强附会的,但是,在数学面前,生物学和物理学是平权的。

图2‐3‐39
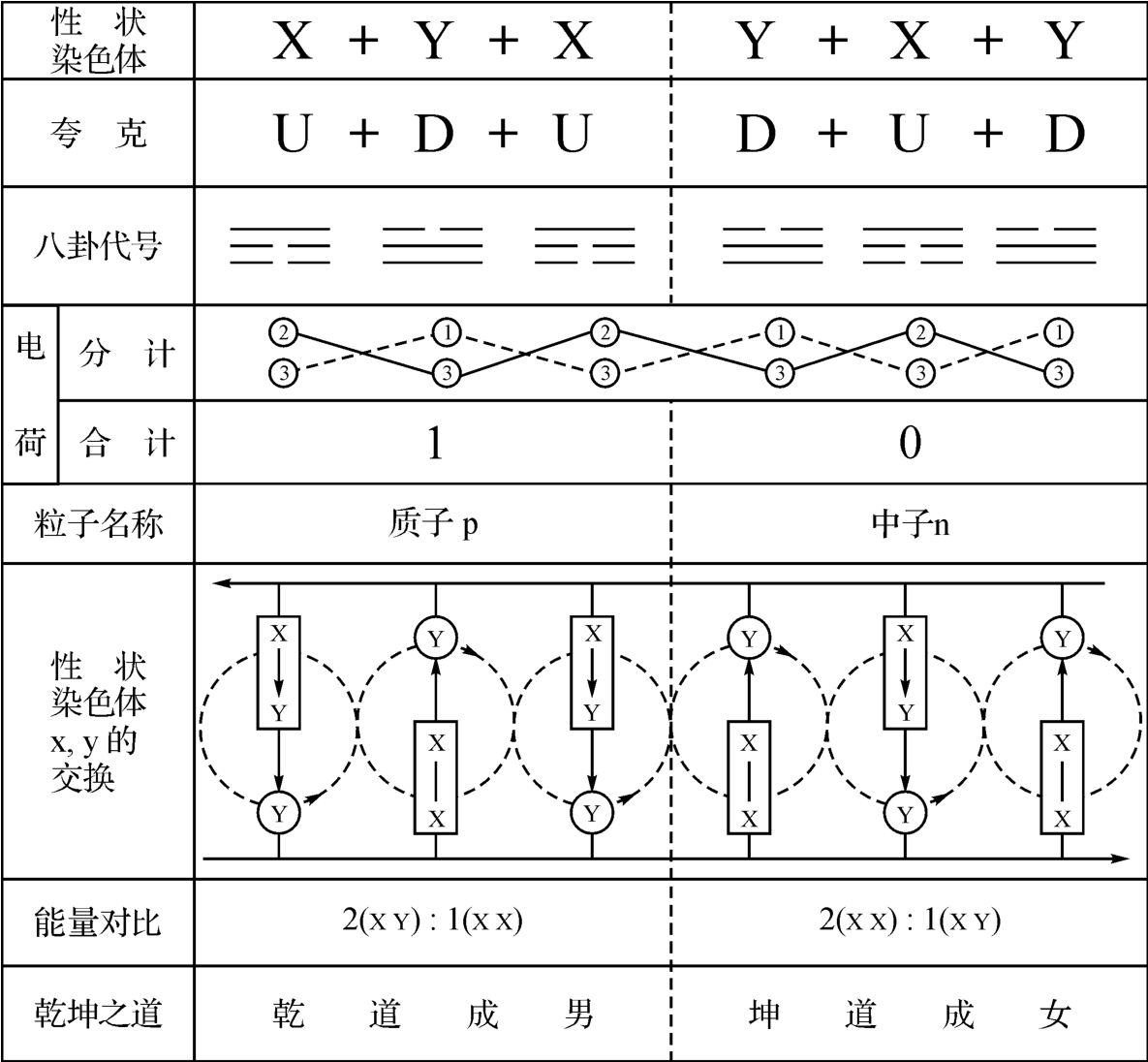
图2‐3‐40
【注释】
[1][美]M.克莱因著,李宏魁译:《数学:确定性的丧失》,湖南科学技术出版社,1997年,第357页。
[2]引自A.G.Cairns Smith.Seven Clues to The Originoflife。
[3][美]S.钱德拉塞卡著,杨建邺、王晓明译:《莎士比亚、牛顿和贝多芬——不同的创造模式》(第2版),湖南科学技术出版社,2007年。
[4][比]伊·普里戈金,[法]伊·斯唐热著,曾庆宏、沈小峰译:《从混沌到有序:人与自然的新对话》,上海译文出版社,2005年。
[5]M.Mitchell Walrop,Science,1990-4-27.
[6]Greg Pendle booth:《网状的世界》,New Scientist,1993年12月4日。
[7][美]S.考夫曼著,李绍明译:《秩序的起源:进化中的自组织和自然选择》,湖南科学技术出版社,2004年。
[8]黄顺基主编:《自然辩证法概论》,高等教育出版社,2007年。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










