



第一节 生态学的层次和生态因子
一、生物与环境关系是相互作用的
生物和环境的关系很密切,生物离不开环境,环境也少不了生物。生物所需要的氧气、水分和营养物质等均来自于环境;生物作为环境的重要成分又存在于环境之中,并在环境改造中发挥积极的作用。
生物的生活环境(Environment)是指生物周围一切生物的和非生物因素的总和。通常环境指的是人类环境,它是环绕着人群的空间,及其中可以直接或间接影响人类生活和发展的各种自然因素和社会因素的总称。包括大气、水、土地、矿藏、森林、草原、野生动物、野生植物、水生生物、名胜古迹、风景游览区、温泉、疗养区、自然保护区、生活居住区等。
生物与其生活环境之间是相互联系、紧密相关的。环境从根本上决定着生物的分布和生活等习性。但是生物也并非完全依附于环境而生存。在长期的生存斗争中,生物都在不同程度上获得了适应环境的能力和潜力。不同的生物适应环境的能力是不一样的,能够在较宽限度范围内生活的生物称为广栖生物,如老鼠等。只能在小范围内生活的生物称为狭栖生物,如造礁珊瑚,它只能生活在温暖、清澈的热带、亚热带浅海环境。生物适应环境的方式也是多种多样的,有些生物具有独特的生活方式如冬眠、夏眠、迁移、保护色及警戒色等来适应各自特殊的生活环境。生物适应环境的能力很强,从两极至赤道,从高原至深海平原都有生物的足迹。但是生物适应环境的能力毕竟还是有限的,当环境条件在生物所能忍受的一定限度内变化时,生物可以通过调节自身的生理机能、改变生活方式或主动地寻找新的生活环境来适应环境的变化,但当环境变化超出一定的限度时,生物就会因无法适应新的环境而导致死亡。
故而,研究生物与环境的关系具有重要的理论和现实意义。弄清生物与环境的关系、生物与环境的变化规律,对于我们了解生物的进化规律和地史时期地球的变迁具有重要的理论价值。当今社会、人口、环境和资源等全球性的问题正困扰着世界各国的和平与发展,通过对环境与生态的研究将有助于我们正确处理好经济建设和环境保护的关系,维护整个地球自然生态环境的平衡,以保证人类社会能够快速持续地向前发展。
二、生态学的层次
研究生物与环境之间相互关系及其作用机理的科学即生态学(Ecology)。“生态学”(Oikologie)一词是1865年由勒特(Reiter)合并两个希腊字oikos(房屋、住所)和logs(研究)构成。1866年德国动物学家赫克尔(Ernst Heinrich Haeckel)初次把生态学定义为“研究动物与其有机及无机环境之间相互关系的科学”,特别是动物与其他生物之间的有益和有害关系。从此,揭开了生态学发展的序幕。在1935年英国的Tansley提出了生态系统的概念之后,美国的年轻学者Lindeman在对Mondota湖生态系统详细考察之后提出了生态金字塔能量转换的“十分之一定律”。由此,生态学成为一门有自己的研究对象、任务和方法的比较完整和独立的学科。后来,在生态学定义中又增加了生态系统的观点,把生物与环境的关系归纳为物质流动及能量交换;20世纪70年代以来则进一步概括为物质流、能量流及信息流。
生态学是研究以种群、群落和生态系统为中心的宏观生物学。生态学的一般规律大致可从种群、群落、生态系统和人与环境的关系四方面说明。
(一)种群
地球上任何一种动物或植物都是由许多个体组成,这些个体在地表总是占据着一定的地区。我们把占据着一定环境空间的同一种生物的个体集群叫做种群(Population)。换句话说,种群就是特定空间同种有机体的集合体,其基本构成成分是有潜在互配能力的个体。
种群是由个体组成的,但是当生命组织进入到种群水平时,生物的个体已成为较大和较复杂生物体系中的一部分,此时,作为整体的种群出现了许多不为个体所具有的新属性,如出生率、死亡率、年龄结构、分布格局和某些动物种群独有的社群结构等特征。在自然界,种群是物种存在、物种进化和表达种内关系的基本单位,是生物群落或生态系统的基本组成部分,同时也是生物资源开发、利用和保护的具体对象。因此,种群已成为当前生态学中一个重要研究对象。
种群的结构通常是指种群的年龄结构,此外,也可考虑其性别结构或世代结构等。年龄结构是指种群中的个体按其生命史不同阶段的分组。种群的年龄结构是种群的重要特征之一,它影响着种群的出生率、死亡率和增长率,同时还对其他物种与环境发生影响。
出生率和死亡率是影响种群增长的重要因素:出生率是指种群中单位时间内每1000个个体的出生数;死亡率则是指种群中单位时间内每1000个个体的死亡数。
任何种群都是由老、中、青、少不同年龄个体组成的,这就形成了特定的年龄结构。研究种群的年龄结构,对于了解种群的密度、预测未来发展趋势和采取相应管理措施等具有很重要的意义。种群年龄结构可分成以下三种类型:
增长型:表示种群中有大量幼体和极少数的老年个体,其出生率大于死亡率,是一个迅速增长的种群。
稳定型:出生率与死亡率基本持平,是一个数量稳定的种群。
衰退型:出生率小于死亡率,是一个数量下降的种群。
另外性比率也是与种群动态有关的重要结构特征之一,它是指雌雄两性个体数量在种群中所占的比率。
种群增长是种群动态的主要表现形式之一,它是指随着时间的变化,一个种群个体数目的增加。自然界中存在几种不同的种群增长曲线,如图8-1所示。左图“J”形曲线是一个单独的种群在食物和空间充足、无天敌与疾病、无个体的迁入与迁出等因素存在,按恒定的瞬时增长率连续地增殖时,种群的总个体数随时间而变化的曲线图,该种群便表现为指数式地增长。而实际上,由于限制生物增长的生物因素、非生物因素(即环境阻力的存在,如有限的生存空间和食物,种内和种间竞争,天敌的捕食,疾病和不良气候条件等)和生物的年龄变化等,必然影响到种群的出生率和存活数目,故而这种按生物内在增长能力即生物潜力呈几何级数或指数方式的增长,在自然界是不可能实现的。通常是当种群侵入到一个新地区后,开始时增长较快,随后逐渐变慢,最后稳定在一定水平上,或者在这一水平上下波动,如图8-1右图的“S”形曲线图所示。此时个体数目接近或达到环境所能支持的最大容量或环境的最大负荷量,这个固定值被称为“环境容量”。
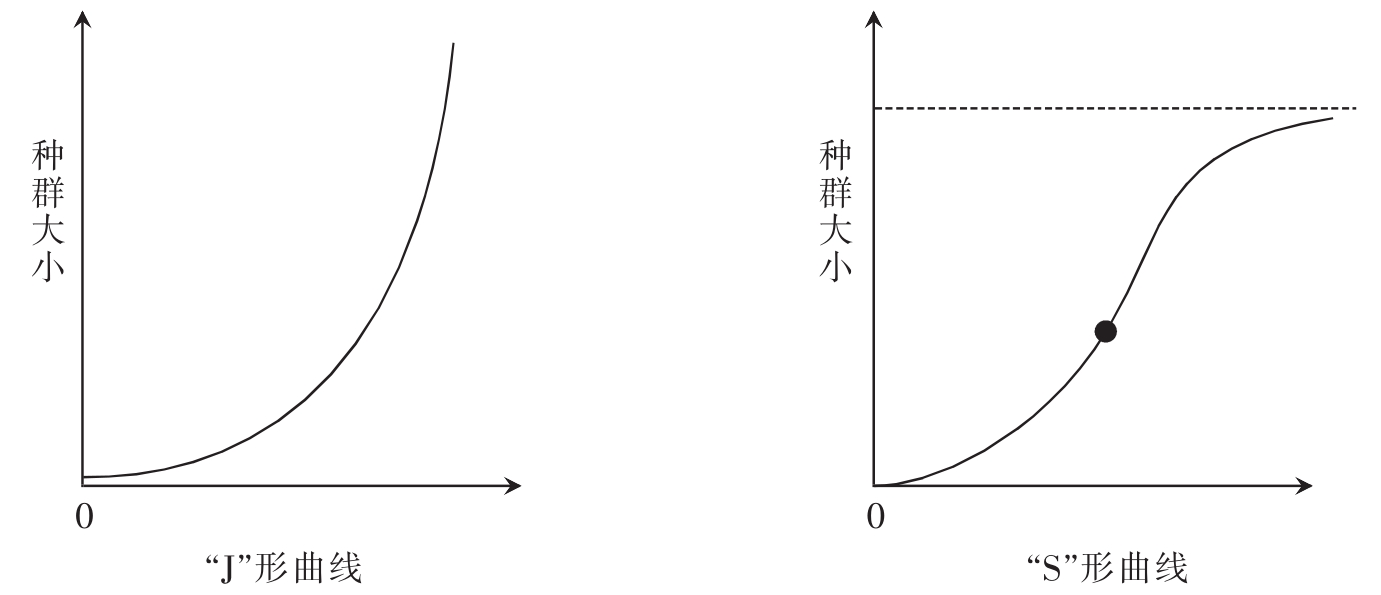
图8-1 种群增长曲线
这种向着环境负荷量极限逼近的“S”形增长曲线反映了一定时期内生物、自然种群增长的普遍规律。但寿命长的、世代重叠多的种群会产生较大的偏差,只有低等的生物,如细菌、酵母或浮游生物等才与之比较吻合。
(二)群落
在自然界,任何生物种都不是孤立地生存,总有许多其他生物种与之同群共居,形成一个完整的生物群体。正如种群是个体的集合体一样,群落(Biological Community)是种群的集合体,是一个比种群更复杂、更高一级的生命组织层次。群落因其组成成分中生物类别不同而有不同名称。如果在一定地段上,共同生活在一起的植物种以多种多样的方式彼此发生作用,形成一种有规律的组合,这种多植物种的组合就叫做植物群落。它是不同种类植物松散地组织起来的单位。河漫滩上的一块草地,山坡上的一片松林,湖岸浅水处的一片芦苇丛,乃至一块人工管育的稻田都是植物群落。其类型繁杂多样,其面积差别悬殊,彼此之间的边界明显或不明显。一个地区全部植物群落的总体,叫做该地区的植被(Vegetation)。如北京的植被、秦岭山地的植被等,都是指该地区范围内分布的全部植物群落。
动物同植物一样,也常常是以群落的形式组合在一起共同生活着。只是由于动物的流动性很大,群落的组合更松散,在科学研究上多以种群为对象而很少应用“动物群落”一词。
(三)生态系统
在自然界,任何生物群落都不是孤立存在的,它们总是通过能量和物质的交换与其生存的环境不可分割地相互联系、相互作用着,共同形成一种统一的整体,这样的整体就是生态系统。换句话说,生态系统(Ecosystem)就是在一定地区内,生物和它们的非生物环境(物理环境)之间进行着连续的能量和物质交换所形成的一个生态学功能单位。
按照生态系统的上述定义,我们既可以从类型上去理解,例如森林、草原、荒漠、冻原、沼泽、河流、海洋、湖泊、农田和城市等;也可以从区域上理解它,例如分布有森林、灌丛、草地和溪流的一个山地地区,或是包含着农田、人工林、草地、河流、池塘和村落与城镇的一片平原地区等,都是生态系统。整个地理壳便是由大大小小各种不同的生态系统镶嵌而成。生态系统是地理壳的基本组成单位,它的面积大小很悬殊,其中最大的生态系统就是生物圈,它实质上等于地理壳。
任何一个能够维持其机能正常运转的生态系统,必须依赖外界环境提供输入(太阳辐射能和营养物质等)和接受输出(热、排泄物等),其行为经常受到外部环境的影响,所以它是一个开放系统。但是生态系统并不是完全被动地接受环境的影响,在正常情况下,或者说在一定限度内,其本身都具有反馈机能,使它能够自动调节,逐渐修复与调整因外界干扰而受到的损伤,维持正常的结构与功能,保持其相对平衡状态。因此,它又是一个控制系统或反馈系统。
生态系统概念的提出,使我们对生命自然界的认识提到了更高一级水平。它的研究为我们观察、分析复杂的自然界提供了有力的手段,并且成为解决现代人类所面临的环境污染、人口增长和自然资源的利用与保护等重大问题的理论基础之一。
生态系统具有以下特征:
1.具有自我调节能力。
2.能量流动、物质循环和信息传递是生态系统的三大功能。
3.生态系统中营养级数目一般不会超过4~5个。
4.生态系统是一个半开放的动态系统,要经历一个从简单到复杂、从不成熟到成熟的演变过程,其早期阶段和晚期阶段具有不同特性。
(四)人与环境
由人类社会与环境组成的世界系统在基本层面上可以概括为三种生产——物质生产、人的生产和环境生产——的联系。三大生产呈环状结构,物质流在它们之间的畅通程度决定着世界系统的和谐程度,进而决定着人类社会是否可持续发展。
人的生产环节其基本参量是人口数量、质量与消费方式,其中人口数量和消费方式共同决定了社会总消费(对物质生产产生的生活资料的消费与对环境生产产生的生活资源的消费)。在传统的生存方式或模式中,社会的总消费决定着自然资源的消耗。社会总消费的无限性提高与自然环境有限性的供给(产出)的矛盾日益严重,导致世界系统失控。人口素质包括人的科技知识水平和文化道德修养,它不但决定人参加物质生产、环境生产的能力,而且表现为调节自我生产和消费方式的能力。因此,人口素质的提高,不仅会体现在单种生产,如物质生产和环境生产的提高和人的生产的改善上,还会体现在调节三种生产的能力的提高上。由此可见,人既是社会可持续发展的动力与源泉,在某种意义上又是社会可持续发展的负担。因此,人口规模适度、人口素质优良与人口结构合理的状态是实现人类社会可持续发展的必要条件。
人口(Population),通常是指一个地理区域的人的数目。对人口进行研究的科目有社会学和地理学,它是人口学研究的最基本的范畴。一个地域的承载能力是有限的,这限制了人口的数目的增长。一些人类社会学观察员建议这个理论同样适用于人口,并且认为未受抑制的人口增长有可能导致马尔萨斯学说的灾难。但是也有人表示不同意这个观点,人口具有自然的和社会的两重属性。
截至到2011年1月,全球人口总数为6,930,686,985,且人口数量每年还在不断地增长。下面两图分别为根据美国人口普查局2011年公布的人口调查结果统计所得的从1950年到2010年的世界人口的总数和年增长率(图8-2,图8-3)。联合国预计2012年全球人口将达到70亿,而到2050年时,全球人口将达91.5亿。中国和印度是世界上人口最多的两个国家。联合国人口基金发表《2010年世界人口状况报告》,预测印度的人口在2050年将增至16.14亿,成为世界第一人口大国。
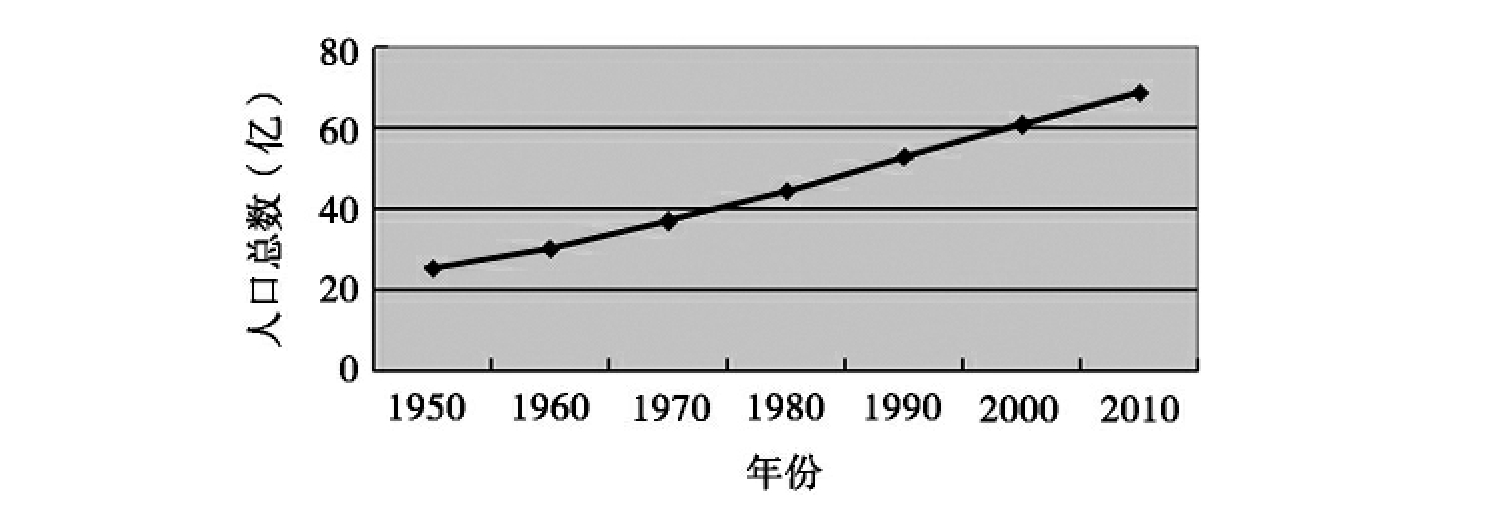
图8-2 1950~2010年世界人口总数

图8-3 1950~2010年世界人口的年增长率
人类的人口增长和任何生物种群一样,是有限度的,因为环境容纳量是有限的。虽然人类可以改造环境,增加环境容纳量,但这也是在一定限度之内,人类所居住的地球资源毕竟是有限的,因此人口无节制增长,必将带来灾难。
人口问题是当今世界面临的难题之一。它既包括由于人口过度增长带来的教育普及、就业困难、住房紧张、社会保险不足、人均收入和健康状况下降等困难(主要表现在亚、非、拉三大洲发展中国家),也包括由于人口零增长或负增长带来的劳动力不足、老龄人口福利水平下降等新难题(主要表现在欧美的发达国家),同时还包括由于经济发展不平衡造成的人口迁移等问题。站在全球的角度来讲,人口的过度增长是所有人口问题的根源之一。主要表现有:
1.归因于人口过度增长带来的人均耕地、人均资源、人均粮食、人均能源、人均国民生产总值、人均收入、人均住房面积等不断下降的形成机制、地区差异及解决措施等。其中以中国和印度等亚、非、拉发展中国家表现最为突出。
2.归因于人口零增长甚至负增长带来的人口老龄化问题,包括老龄化的形成机制,对社会福利制度的影响,劳动力的短缺,社会伦理观念的转化,以及政府相应的政策等。其中以西欧、北欧、北美诸国表现比较突出。
3.与人口结构有关的问题。如人口的年龄结构、性别结构、学历结构、就业结构、城乡结构等。与现实密切相关的是就业结构,与人口增长相关的是年龄结构,与未来发展相关的是学历结构。
4.农村地区人口过度增长带来的过度开垦,造成森林草场破坏、水土流失、土地沙化、河湖淤塞等相关的生态问题。另外,还有农村年轻劳力外出打工造成的劳力短缺问题,农村剩余劳力大量拥入城市(即城市化过程),民工的盲目流动带来的交通紧张、社会治安变差、住房紧张等其他社会问题。
5.国际人口迁移过程中带来的文化交流、种族融合问题,以及对迁入地经济繁荣的促进、社会环境的破坏等问题。尤其要注意国际人口迁移的原因、特点、路线及后果。
中国社会科学院人口与劳动经济研究所研究员张翼博士认为“十二五”期间中国必须关注的三大人口问题是:
1.中国人口收缩且增速放缓。人口收缩主要是指人口金字塔底部的缩小,即在计划生育政策或社会与经济发展的约束之下,由于出生率的降低而导致的少儿人口年出生率和新生人口数量的逐步稳定下降的态势。中国人口金字塔收缩会引发人口老化速度加快,且各地分布极不平衡和人口出生性别比在人口收缩中趋于上升等问题。
2.中国人口的流动与流动人口的结构转变。中国当前的人口流动主要由劳动力人口的就业需求所引发,因为子女教育、养老照料、移民搬迁等引起的流动等开始增加。中国当前的人口流动与发达国家的人口流动有所不同,但与发展中国家向发达国家的移民具有某种程度的相似性,既体现着非农化与城市化的“拉力”,又体现着城市人口金字塔底部收缩所带来的“吸力”。尤其是乡—城人口流动,与中国人口收缩态势具有极其强烈的相关性——城市的人口规模越大,城市户籍人口的收缩态势越明显,其独生子女率越高,对流动人口的需求也就越旺盛。
3.中国人口开始迅速集聚。当前,中国人口流动带来了经济活跃地区人口数量的增加,导致了人口开始了工业化与非农化拉动的迅速集聚。这种集聚表现为城市化带来的大集聚与小城镇建设带来的小集聚及这两种集聚的并行发展。
解决好人口问题,简单地从人口增长模式本身去寻找解决问题的途径和办法是远远不够的,必须把人口问题放在国民经济和社会发展的全局中考虑和处理,正确处理好人口、资源、环境的关系,才能促进人类社会的和谐发展。
我国将以科学发展观为指导,强调全面、协调和可持续发展,促进人与自然、人与社会、人与人之间的和谐发展,统筹解决人口数量、素质、结构、迁移和安全问题。由通过行政控制手段为主,转向用宣传教育、依法行政、技术服务、促进健康、社会保障等方法,形成统筹解决人口问题的新局面。
目前,我国正采取的五项措施是:积极预防出生缺陷发生,促进人口素质和健康水平全面提高;加快推进农村计划生育家庭奖励扶助制度,探索与经济发展水平相适应、有利于计划生育的农村社会保障体系;对人口性别比升高问题实行综合治理,完善政策法规体系,开展“关爱女孩行动”,依法保护女孩的生存发展权;积极促进人口分布和就业结构比例合理,努力实现城乡共同发展和进步;创建环境生态良好的人居环境。
三、生态因子
生态因子(Ecological Factor)是指环境中对生物的生长、发育、生殖、行为和分布有着直接或间接影响的环境要素,如温度、阳光、湿度、大气及其相关生物等。生态因子是生物生存所不可缺少的环境条件,也称生物的生存条件。生态因子也可认为是环境因子中对生物起作用的因子,而环境则是指生物体外部的全部环境要素。
在任何一种生物的生存环境中都存在着很多生态因子,它们彼此之间相互制约、相互组合,构成了多种多样的生存环境,为各类生物的生存和进化创造了不计其数的生境类型。生态因子据其性质可归纳为:
1.非生物因子
气候因子:如温度、湿度、光、降雨、风、气压和雷电等。
土壤因子:包括土壤结构、土壤有机和无机成分的理化性质和土壤生物等。
地形因子:如地面的起伏、山脉的坡度和阴坡阳坡等。
2.生物因子
生活环境中的生物因素主要是指生活在一起的各类生物之间的相互关系,如捕食、寄生、竞争和共生等(有时为了强调人的作用的特殊性和重要性,把人为因子从生物因子中分离出来)。但产生这种关系的最重要的因素是食物链。所谓食物链(Food Chain)是指通过处于不同营养水平的生物之间的食物传递形成一环套一环的链条式关系结构。它是一定的环境范围内各种生物通过捕食方式而产生的直接或间接的联系,是生物界普遍存在的现象。而在生态系统中,各种食物链相互交错、互相联系所形成的错综复杂的网状结构叫食物网(Food Web)。
四、生态位
在对大自然的研究中,人们发现这样一种奇妙现象:即亲缘关系接近的、具有同样生活习性或生活方式的物种,一般不会在同一区域出现。如果它们在同一区域出现,那么严酷的竞争将不可避免,并且最终大自然会用空间把它们各自隔开。自然界这一奇妙现象,生态学家将其称为生态位现象。生态位(Ecological niche),又称小生境、生态区位、生态栖位或是生态龛位,是一个物种所处的环境以及其本身生活习性的总称。每个物种都有自己独特的生态位,借以跟其他物种作出区别。生态位包括该物种觅食的地点,食物的种类和大小,还有其每日的和季节性的生物节律等。
在现代生态学中,生态位不仅已成为了一个重要的专有名词,而且生态位理论也已成为重要的基础理论之一。由于生态位理论在探讨种间关系、生物多样性、群落结构及演替和种群进化等研究方面有着广泛应用,因此生态位理论在现代生态学中占有的位置,就显得愈来愈重要。
群落生境(栖息地)是生态位这个概念的一部分。生态位的含义远不止是“生活空间”(温度、空气湿度等环境因素的综合,它是生物生存的依据)的一个抽象概念,它描述了一个物种在其群落生境中的功能作用,而且带有构成群落生境的自然因素所留下的烙印,是一个物种为求生存而所需的广义“资源”。例如:蝙蝠需要在某地夜间捕食蚊子。这里面某地的自然因素(例如空气质量等其他关系到蝙蝠栖息地的因素),蝙蝠夜间运动的可行性,蚊子都是蝙蝠的生态位的一部分。一个物种只能占有一个生态位。
生态位分两个层次:
基本生态位:是生态位空间的一部分,一个物种有在其中生存的可能。这个基本生态位是由物种的变异和适应能力决定的,而并非其地理因素。或者说基本生态位是实验室条件下的生态位,里面不存在捕食者和竞争。
现实生态位:是基本生态位的一部分,但考虑到生物因素和它们之间的相互作用。或者说是自然界中真实存在的生态位。
两个地区,虽然地理上被分隔,但却有着相似的非生物因素,在这两个地区生活的物种会占有相似的生态位。这会导致趋同演化的发生,即两个物种虽然无亲缘关系,但却各自独立地发展出相似的身体构造去适应环境。南极的捕鱼能手、不会飞的鸟——企鹅和已灭绝的欧洲的大海雀一样占有相似的生态位;澳洲的袋鼹和欧洲的田鼠都有挖土铲的前肢,它们在泥土中挖掘通道,捕食细小的动物,它们占有相似的细小地下肉食性动物生态位。
对于一个地理上不存在分隔的生态系统中,在生态平衡时,各个生物的生态位原则上不重合。若有重合,那么必然是不稳定的,它必然会通过物种间的竞争来削减生态位的重叠,直到平衡为止。竞争,比如需要相似生态位的入侵物种的进入,会导致土著物种存在区域减少。如果存在区域太小,会导致一个物种的灭绝。这就是竞争排除原则。
进化导致的是两个有亲缘关系的物种去占据不同的生态位,减少竞争的机会。一个很好的例子是拉帕戈斯群岛的达尔文雀的进化(适应辐射)。
换句话说,进化以及新的物种出现的重要原因是空余的生态位出现了,而这种情况大多是由于灾难或者地理分隔导致的。
生态位的数量与生态系统的气候、地理和生物因素有关。相应的物种数会因这些因素的差异导致有很大的不同。极地(例如格陵兰的冰川)、南极洲或是高原的生态位数就不及热带的原始森林或是珊瑚礁的多。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。











