



10.1.5 其他过氧化物酶
10.1.5.1 细胞色素c过氧化物酶
某些细菌以及酵母线粒体的内膜与外膜间,存在血红素为辅基的细胞色素c过氧化物酶。该酶与H2O2可生成复合物。后者可使还原型细胞色素c转变为氧化型。
![]()
已知细胞色素c过氧化物酶的最大吸收峰为407nm及该酶与H2O2产生的反应率。这种方法已应用于原生动物及实验室中分离线粒体与过氧化体的研究。
10.1.5.2 NADH过氧化物酶与氧化酶
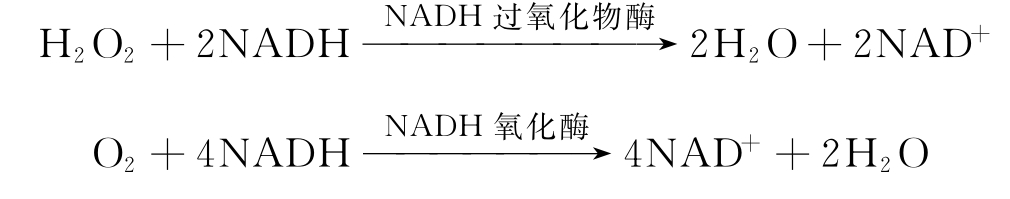
某些细菌如干酪乳杆菌、粪链球菌含有NADH过氧化物酶,后者能利用H2O2氧化NADH成为NAD+。产气荚膜梭菌是一种厌氧菌。其耐氧变种亦含有NADH过氧化物酶。
NADH氧化酶的作用与NADH过氧化物酶虽都可使NADH氧化为NAD+,但NADH氧化酶尚能同时使O2还原为水,而NADH过氧化物酶却只能利用H2O2作为氧化剂,使NADH氧化为NAD+。
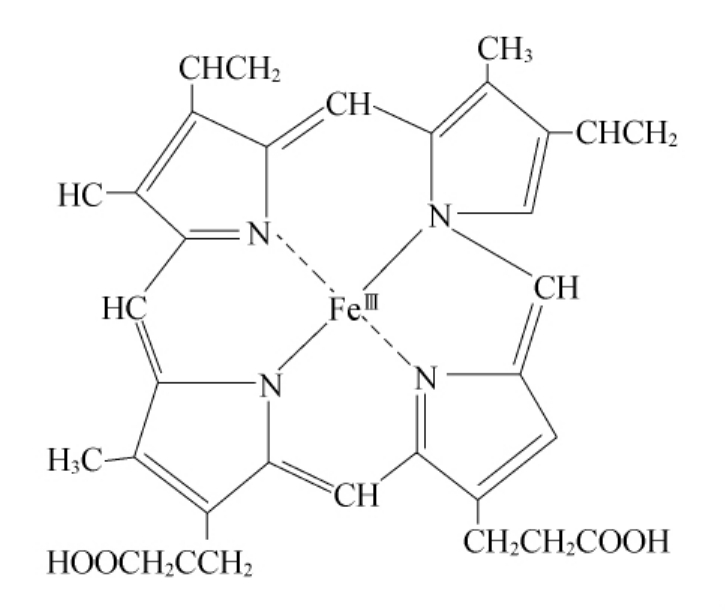
图10-4 辅基血红素(高铁原血卟啉)
在氧存在的条件下某些耐氧的厌氧菌有合成NADH氧化酶的能力,从而可在有限氧气中生存,但实际上尚显示生长受到抑制的效应。这是因为NADH来自糖酵解等代谢途径,本应参与生长有关的生化反应,而由于进入NADH氧化酶或过氧化物酶的反应,遂使生物受到抑制,但这种抑制效应的出现可使细菌不至于O2中毒而死亡。
10.1.5.3 “非专一性”过氧化物酶
在植物与细菌中存在以血红素为辅基的过氧化物酶,其底物多无专一性。人工合成的底物如愈创木脂醇、联苯胺与邻联二茴香胺,在H2O2存在时,经“非专一性”过氧化物酶的作用可转变为有色物质。在生物体内,该酶的底物尚未查明。在少数动物系统中也有这种酶,例如牛乳与唾液中存在的乳过氧化物酶。除了乳过氧化物酶外,还有以下的“非专一性”过氧化物酶。
a)髓过氧化物酶。该酶存在于吞噬细胞。由于它可催化I-为I2,使后者可连接到甲状腺素分子上,它与甲状腺素的生物合成有密切联系。
b)子宫过氧化物酶。已知该酶含量可随雌激素的注射量增加而增加,但对该酶的功能至今尚不清楚。
10.1.5.4 氯化过氧化物酶与溴过氧化物酶
从烟黑酶属(Caldariomyces fumago)中分离出的氯化过氧化物酶,在H2O2与卤族离子如Cl-,Br-或I-存在的条件下,它可催化卤族原子进入许多底物分子。
![]()
但是存在于海产生物中的溴过氧化物酶,只能催化Br-与底物在H2O2的作用下反应成为溴化合物。
10.1.5.5 抗坏血酸过氧化物酶
从眼虫属中可纯化出抗坏血酸过氧化物酶。它可催化抗坏血酸与H2O2的反应,生成脱氢抗坏血酸与H2O,此反应可被CN-与N3-抑制。植物中常含有较高量的抗坏血酸,可能与清除H2O2有关。
虽然在动物的某些组织中存在“非专一性”过氧化物酶,但这类酶并未广泛存在于动物组织中。
生物学研究中常用的过氧化物酶为辣根过氧化物酶,该酶有数种不同形式,但都含有结合型糖类。该酶对底物专一性的要求不高,如愈创木脂醇、邻苯三酚、NADH、巯基化合物、苯酚、吲哚乙酸等底物都可在辣根过氧化物酶催化下被氧化。如以SH2代表底物,则酶反应式如下所示:

在反应(1)中H2O2从辣根过氧化物酶分子中取走两个电子,其中一个电子来自酶的辅基中Fe(Ⅲ),另一个电子的取走可能造成酶蛋白具有额外的氧化能力。所生成的化合物Ⅰ与底物(SH2)反应生成SH自由基与化合物Ⅱ,然后化合物Ⅱ与另一SH2反应生成SH自由基与过氧化物酶。这表明催化反应(1)中生成的化合物Ⅰ需要通过反应(2)与(3)的单电子接受反应才能恢复原来的酶。其中间物为化合物Ⅱ,另一中间物为SH。两个SH可化合成为S及SH2。反应(4)为歧化反应。因为SH有时可使O2还原为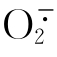 而歧化反应产物之一为H2O2。由此可见,仅需少量H2O2就可启动反应。现以NADH为例,以说明辣根过氧化物酶的催化反应机理。
而歧化反应产物之一为H2O2。由此可见,仅需少量H2O2就可启动反应。现以NADH为例,以说明辣根过氧化物酶的催化反应机理。

这种反应反映出过氧化物酶尚具有氧化酶的效能,因为在正常反应中只需要供给H2O2与SH就可以发生反应,但有些SH自由基如NAD可类似氧化酶,能将O2直接还原为,最后通过歧化反应将转变为H2O2与H2O。
在植物细胞中苯丙氨酸衍生的酚类聚化而成木质,但聚化过程中靠连接在细胞壁上的过氧化物酶,先使这些酚类氧化成酚氧自由基,然后自由基中间物才聚化。在氧化反应中所需的H2O2是结合在细胞壁上的苹果酸脱氢酶产生的NADH在过氧化物酶作用下的氧化产物。此外,植物过氧化物酶尚作用于植物激素吲哚乙酸的降解,从而参与植物生长的调节。
除了上述实例外,要在整体实验中鉴别植物与细菌的非专一性过氧化物酶的底物以及评价该酶清除H2O2的能力比较困难。许多植物根茎部组织所出现的超弱化学发光可能来自过氧化物酶的催化反应。例如吲哚乙酸降解产生的激发态吲哚乙酸可生成dioxetane中间物,然后分解成激发态羰基化合物。当后者转变为基态时就发出光,或者激发O2成为。
在实验室中辣根过氧化物酶可用于检测H2O2的产生。其反应底物为7-羟基-6-甲氧香豆素。后者可在360nm光波激发下发出450nm的荧光;但经辣根过氧化物酶作用后,荧光强度随着H2O2产生量而相应减少,故根据荧光丧失量可推算出H2O2产生量,但其他底物亦可与7-羟基-6-甲氧香豆素竞争,从而造成H2O2的测定结果偏低。此法不适合于叶绿体中的H2O2产量测定,因为叶绿体中含有大量抗坏血酸,而后者恰好可被过氧化物酶氧化。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。










