



I
我们躯体的细胞对氧的依赖要比我们从外部世界所能得到的任何其他物质都更为密切。职业性饥饿试验者证明,我们可以不依靠食物而靠利用躯体储备的糖元、脂肪和蛋白来生活几个星期,最后再靠我们自己肌肉和腺体的现有组织而不致发生任何可以化验出来的永久性的损伤。有过艰苦经历的人证明,我们在不摄入水的条件下也能过几天;无疑,也还是依靠利用机体所储备的水分。但是,对氧的需要来说,情况就不同了。脑部有重要的神经细胞,如果完全断绝氧气达8分钟以上,则发生无法恢复的严重破坏。可以合理地认为,这种特殊的差异是和人体管理上不存在氧的储备有关。食物和水不是经常地被接纳的,储存要靠充足时的积累而在短缺时加以利用。氧则与此不同,经常存在于我们周围的无限大量的空气中,约占其21%。无论何时需要我们都可以得到它,因而不需要储存。
当氧的供应可能发生障碍时,各种情况自然就出现了。在出血状态下的这种情况我们已经讲过了。坐飞机升空或攀登高山而达到大气中氧分压大大减少的地带时,又出现另一种情况。照明气体或汽车废气的中毒——这种状态下循环红细胞中氧的位置被一氧化碳所夺去并与之牢固地相结合——也同样产生一种相对的缺氧血症。但这些或多或少地可以视为非自然状态。
机体连续不断的活动,诸如心脏的搏动,呼吸的节律性运动,骨骼肌经常的轻微收缩,都要求连续不断地输送氧气以供这些活动所产生的废物的燃烧之用。无论在什么时候,一当这些活动增加,例如在剧烈的体力劳动中,原来在休息状态下足够的最低限度的供氧将远远落后于新的需要。以致这种活动不能再继续下去。机器将因堆积废物的阻塞而很快地被迫停止运转。在此情况下稳态要靠加速(供氧的)连续作用来达到。
II
一个具有平均身材的人,在休息状态下氧的需要量只为每分钟0.25到0.30公升(夸脱)[1]。当他转入很重的体力活动时可增至每分钟15公升以上。但是即使在最有利的情况下,躯体吸入和利用大量氧的最大能力不过每分钟4公升。因此,在高度激烈的运动期间,氧的吸入量可达到休息期间的10到12倍,但仍远不足以满足当时的实际需要。
发生这种情况时,肌肉收缩的伴随产物乳酸就不能氧化成二氧化碳或再转化为其前质——糖元。结果是,乳酸蓄积在肌肉里。收缩可以继续进行,但由于这种酸浓度的增高干扰了收缩过程而使其效能降低。显然,大量的体力活动不能依靠即时的氧化作用。我们在一次剧烈活动中,比如在一次225码赛跑中所产生的乳酸,先是临时在肌肉和液床中得到中和,以后才被氧化,或转化为它的前质。这样我们进行工作的能力就可以远远超出当时输氧能力所规定的能量供给限度。但是乳酸在体内积存起来。这些乳酸是必须予以清除的。以乳酸钠形式存在于血液中的一部分乳酸可通过肾脏排出。但绝大部分是被氧化或再转化为糖元。这种氧化发生在运动停止之后。所以,我们固然能在供氧限度之外支借到继续工作的能力,但必须是在后来吸入足够的氧气以氧化所积存的废物为条件。这样我们就增加了希尔(Hill)所说的“氧债”。我们要在完全停止活动后半小时或更多的时间内仍继续做深呼吸以偿还这个债。一个人在23.4秒内跑完225码后,过27分钟才能回到正常的平静呼吸。这个期间所用的额外的氧,其量超过了同等时间的安静状态下氧的消耗,我们可以以此来测定氧债的多少。
从以上的讨论中可以明了,只有在中等的活动状态下才能保证对机体内远距离细胞供氧的稳态。无论何时只要从事高度用力的活动或长时间的剧烈运动,单靠呼吸和循环系统的调节就不足以防止液床中产生过多的酸,因而需要一种能够维持其中性反应的装置发生作用。所以,本章中关于氧的供给和下一章关于血液的中性这两个问题是密切相关的。
III
为了供给肌肉作工时额外所需的氧,肺、心脏和血管的机能要以种种不同的和复杂的方式进行调整,其各自的目标都是为了保障足够的供氧量以满足工作器官的需求,或者是用来偿还氧债,如果在肌肉活动的当时氧的需要未能得到满足的话。
首先是,呼吸加深并加快了。这种变化在体力活动的一开始就立即发生,其原因除了那些引起体力活动的并来自大脑皮质的神经冲动之外,还有一种别的作用——伴随主要作用而自动产生的一种附加作用。随之而来的是我们在剧烈运动时都看到的更大量的呼吸——更深更快的呼吸运动,这是由于血中二氧化碳增加所致。这种增加主要是工作肌肉中所产生的乳酸被氧化为二氧化碳(CO2),使肌肉大量生成CO2的结果。肌细胞中CO2较淋巴液中的浓度高,这使前者具有更高的扩散压(即更大的扩散倾向),从而使它扩散入淋巴,因此,由于同样的道理,使它扩散到流经毛细血管的血液中。血液把它输送到肺。
在肺内,血液分布在细小的毛细血管网中,后者位于小气囊即肺泡的极其细薄的壁上。这些(肺泡)是树状呼吸管道中最小分支的顶端,其树干是气管,其树枝就是支气管。据估计,如果把一个人肺中所有的肺泡展平成一个连续的平面,将构成一个大约90平方米,亦即大约1100平方英尺的表面积,其中800平方英尺以上与肺毛细血管接触。显然,这为血液和肺泡腔之间进行气体交换提供了一个非常巨大的面积。
肺泡气中二氧化碳的百分率约为5.25。当过量的二氧化碳被带入肺内静脉血液时,后者所含的二氧化碳就有了比肺泡内更高的扩散压,因此二氧化碳就由血液透出而进入肺泡腔。然而这个过程在气体由血液入肺的倾向和与此相反的倾向间达到平衡时就不再进行。在此过程中,肺泡内CO2百分率当然上升,结果是,气体从血液中的逸出就不比正常处于安静状态下那样多。因此,当血液通过并离开肺部时运出的CO2比平时更多。于是这种二氧化碳扩散压增高了的动脉血来到位于大脑下面延髓中的“呼吸中枢”。在这里,或因血液中的气体扩散到中枢细胞内,或因血液中较高的分压阻止了该部细胞活动所产生的二氧化碳的向外扩散,于是细胞内的二氧化碳含量增高。这就等于一个刺激。这时细胞把神经冲动释放到呼吸肌而使后者的动作变得比原来更加强有力。这种加强了的肺换气作用自然导致肺泡内过多的二氧化碳的呼出。当此过程持续到百分率下降至平静状态下的正常水平的程度时,对呼吸中枢的额外刺激就消失掉了,这样又开始了平静呼吸。对于维持液床相对恒定的连续作用的自动调节来说,没有比脑部的这种精巧装置更好的例子了,它通过保持自身状态的均衡来保护机体所有其他部分免于酸性废物的蓄积。这种敏感性的若干指征可在海登和泼瑞斯特里所证明的下述事实中看到:即肺泡中二氧化碳仅仅增加0.22%就能使肺部换气作用增加100%!
在剧烈的体力活动中,人体肺部的换气可以从每分钟6公升增加到60公升以上。这意味着支气管中气体的进出必须比原来快很多,而产生的摩擦也要大得多。在支气管的管壁上有环状肌肉来控制管腔大小。这些肌肉和小动脉上的肌肉一样,也受来自交感神经系统的神经的支配。布利顿和我按照前面已经说明过的方法,利用去神经心脏证明,即使在轻微的肌肉运动时,交感—肾上腺系统也是起作用的。图27就是表明一只动物仅在走过地板时
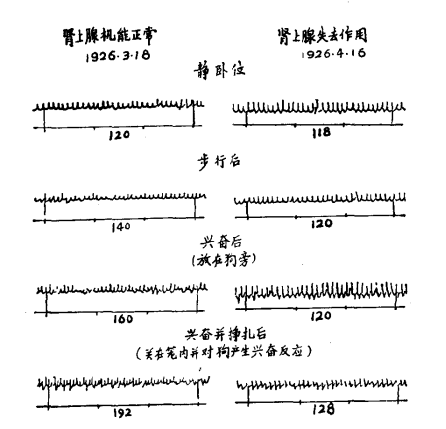
图27:27号猫去神经心脏在肾上腺分泌失去作用(4月6日)及在此以前(3月18日)和以后(4月16日)分别记下的心率原始记录,记录时间间隔5秒。动物在卧位安静状态下的基础心率,分别用来和步行后、面对狗发生兴奋后、被关在笼中并对吠叫的狗发生兴奋后各种状态下的心率进行比较。
去神经心脏速率加快了。在此例中增加了20次。经过这种活动的27例中,脉率平均增加为每分钟17次。在这些情况下,脉率的增加是单独由肾上腺分泌引起的,因为如读者在图27中所看到的,在肾上腺失去活性后同样的肌肉运动所引起的增加只有微不足道的两次。要看到,在动物挣扎时为了使去神经心脏的心率进一步加快,交感—肾上腺系统参与的作用也更大。在不同动物中的大量观察中,其平均增加次数为每分钟49次。交感—肾上腺系统对细支气管的效应是使它们扩张。用肾上腺素治疗支气管哮喘时病人所体验的那种明显的轻快感,就是对这种效应的一个很好的说明。因此,细支气管在大量肌肉活动中比在休息时更为舒张看来是完全可能的。这种舒张对激烈活动时所需的大量空气的进出提供了更大的通道,并能因此减少为摩擦所要作的功。
在呼吸作用管理上又一个出色的特点是,通过深呼吸呼出二氧化碳以防止它在肺中——自然也就是防止它在血液中——的蓄积,同时又吸入比平时更多的氧。即使在激烈的体力活动时,肺泡内氧的利用可增加5倍或10倍,但由于这种巧妙的自动调节作用,肺泡气中氧的百分率及其进入血液的扩散压都能维持在正常或稍高于正常的水平。
IV
由于呼吸运动的双重效能,使血液既能排除其挥发性废物——二氧化碳,又能在通过肺毛细血管时立即运载氧气。为了了解继在肺内气体交换之后,存在于循环系统中的调节装置,我们必须记住氧和二氧化碳的运载要靠血液中的红细胞,其数量虽然能在紧急时刻增加,但仍是有限的。我们在讨论严重失血后所发生的代偿作用时已经知道,在此情况下增加气体运输的唯一方法就是增加载体的利用率;换言之,就是增加载体在肺和活动器官间的循环次数。实际情况也就是如此,但还要加上一个作用,就是保证两头“装”和“卸”的过程都能顺利进行。我们现在就来详细考察一下这些调节作用。
首先,为达到增加心脏每分钟搏出量的目的,必须增加通过静脉回心的血量。我们从事肌肉运动时就是通过循环系统的种种改变来达到这个效应的。控制血管口径的血管舒缩神经使分布在胃肠上的大面积的血管产生收缩。结果,这些血管中的大量血液被驱向肌肉血管,我们将看到,后者在肌肉活动时能有一个高度扩大的容量。
再来说肌肉,它在用力时有不同程度的节律性收缩,对血管产生不同程度的节律性压力,尤其是对位于肌肉束内部及其间的小静脉。因静脉中有瓣膜使血流以向心的方向单向运行,这种节律性的压力必然地要促进这种回流。这种效果能够立即加以证明;用右手紧握左腕,然后迅速并反复地握紧和松开左手,注意这只手背上的静脉很快地就发生了充血。对静脉的另一种泵式动作见于巨大的形如屋顶的呼吸肌,它就是把胸腔和腹腔隔开的横膈,或者称之为横膈的机能动作。这个肌肉在收缩时压低其屋顶形的坡度,即当向腹腔下压时其外缘多少变平。它借这种动作对那条大静脉——下腔静脉增加压力,这条静脉把从下肢汇集起来的血液经腹腔向上回送。腔静脉分支内的瓣膜在这里也防止血液流回下肢。因此,腹内压增高有利于血液的向心流动。但胸部条件也有助于这个目的。位于胸腔的肺是处在一种被拉长的状态中;这使肺内的空气向内压缩。因此,胸腔内肺外的器官,例如静脉,其所受压力就低于大气压。每次吸气的动作都使胸腔扩大,肺就进一步被牵引,从而它总是使静脉所受的压力减低。所以,在腹内压增高的同时,胸内压也就降低,这不但进一步推动了来自下肢部分的血流,也同样促进了上肢和头部血液的回流。在下一个呼气动作中,正在回流的静脉血液自然淤积在胸腔之外——即上肢、颈部和腹部的静脉里。但在下一个吸气中又发生刚才提到的那种状态而把淤积的血液驱向心脏。就这样,在四肢肌肉的泵式动作外,再加上横膈的泵式动作,成为促进血液利用的一种因素。
应当注意到躯体中的这种构造的巧妙管理方式。收缩肌肉因其本身的收缩而需要额外的氧,这些肌肉使带氧血液回流,这又自动地帮助了肌肉所需氧的获得。至于横膈,则如我们所知,在运动中会更加有力地发挥着“泵”的作用,不仅满足肺中氧气载体的氧气供应,还要帮助加速载体的循环,从而增加对缺氧组织的氧气供应。
V
心脏输出的血液不可能多于其所接受的血液,而且必须和回流的血液保持同步,否则就会产生反压并导致静脉内的血流停滞。简言之,心脏的排出量是受到从静脉输入的血量的控制的。由于在肌肉工作开始后回心血量大大增加,所以产生了一个问题:即心脏将如何完成任务?这有两个方法:心脏可以在每次搏动中推出更多的血液,其次可以增加搏动速度。实际上,这两个方法都被使用了。
心脏每搏的输出量,也就是扬代尔·亨德森(Yandell Henderson)所称“搏出量”,必须通过间接测定法来确定。不同的研究者各用不同方法在较少的对象身上所得到的结果,其数据只在它们的一般特征上才是一致的。显然,当一个人处于休息状态时,心脏的慢速搏动不会使本身接近于排空;每次收缩后,心室中剩余的血量可为原有量的25%或更多些。当心脏在舒张期放松时肌性的心壁是柔软和松弛的,尤其是接受静脉血的右心部分。因此它接受从大静脉涌入的血液时变得比平时更加膨胀。英国的生理学家施塔林发现了一个高度引人兴趣的重要事实:在一个合理限度内,正常心脏的肌肉纤维越是被牵引,其收缩就越有力!因此涌入心脏的血量越多,就十分自动地引起越加有力的喷射。所以心脏在进行强力收缩时,在其收缩终末,心室中的余血量要比在平静节律下搏动时为少。由于这些调节作用,重劳动时每搏输出量约为处于休息状态下的两倍。
仅仅靠每次搏动——因而也就是每分钟血液的搏出量——的加倍,显然还不能满足必须把氧供给增加到10倍以上这种状态的要求。心脏容量增加所受到的限制是通过加快红细胞的利用速度而得到代偿的。在安静时,我们的心率为每分钟70次左右,而在激烈运动时,可以升高至140次以上。再加上心搏出量加倍,就大大提高了满足肌肉工作中氧额外需要的能力。
有各种不同的动因引起并保持心脏更快的收缩。实际上在工作量达到高潮之前,心率就可以开始加快。在呼吸调节作用中,我们已经注意到,只要有行动的开始动作,就能引起呼吸的加快,因为伴随此动作的神经冲动使脑部呼吸中枢兴奋起来。与此相似,在我们开始行动时,就由于一直对心脏保持阻遏作用的迷走神经受到不同程度的抑制而使脉搏加快。这些作用都是机体面对需要时作出迅速反应的一组机制,这些机制在器官上表现为两种不同的系统,但无需说明,这两个系统在彼此的协同作用中是互相密切关联的。
四肢肌肉和横膈推动静脉血进入右心房、心室的泵式动作,引起静脉内压力增高,这一点可在用力时紧贴皮肤下面的静脉怒张得到证明。这种压力的增高使刚才提到的神经效应得以继续并加强,因为这种压力施加于心腔内时能引起右心房与大静脉连接末端部分的扩张,从而产生一个反射,即所谓“班布利奇反射”(Bain-bridge reflex),它进一步抑制迷走神经对心率的阻遏作用,因而使心搏变得更快并一直保持下去。
人们已经知道,支配心脏的交感神经同样能被右心房静脉压的增高所兴奋。此外,在肌肉高度用力时,该神经也转入兴奋,特别是像在竞赛中那样伴有情绪兴奋时更是如此。我们已知道这些神经的作用在于加速心搏。示意图27的心搏真实记录表明,甚至轻微活动都能唤起交感—肾上腺系统产生作用,而且活动越大,该系统参与作用越强。另外,有证据说明交感性支配的神经中枢可能受到某种程度的像呼吸中枢所受到的那种影响;作为二氧化碳过多的结果,该部位酸性可以提高,而其主要后果将是产生刺激。这种设想得到了马蒂松(Mathison)所做实验的支持,他证明,在呼吸空气中——因而也在血液中——有过多二氧化碳时能使动脉压升高。林顿(Linton)兄弟和我本人所做的一些观察同样说明肌肉活动产生的废物能使交感—肾上腺系统发生作用。我们发现,对完全断绝了与躯体其他部分神经联系的大肌肉群给以人工刺激,能引起去神经心脏速率的加快,这一点肯定了瑞典生理学家约翰松(Johansson)之说。这只能是由于肌肉向循环血液放出化学物质所引起,因为此时活动肌肉与躯体其他部分唯一的联系就是通过血流。但是,如事先使肾上腺失去作用,则在此实验中重复同样刺激并不产生效应,或反使心率多少有些减慢。显而易见,心率的加速并非血中变化的直接效果,而是通过交感—肾上腺装置的途径所产生的效应。
所以,在剧烈的肌肉活动中,为了机体的需要,呼吸系统和循环系统之间的调节明显地存在着密切的关联就可得到解释了;虽然这两个系统都通过随意运动所引起的神经冲动而开始产生心率和呼吸的加速,但它们在执行其额外任务时,则是通过动脉血中二氧化碳浓度的增高而继续保持其活动的。随后,由于它们额外的活动使二氧化碳又减少到休息状态的水平,这两个系统又渐渐地回到自己安静时的日常机能状态。
所有这些协调进行的作用提供了如下的条件,首先,使心脏对静脉流回的大量血液具备充分的接受能力;其次,在肺进行深呼吸以利于呼吸气体(氧和二氧化碳)的更大量交换时保证其有效的血液供应;最后,将带氧血液有力地输出到大面积的“动脉网”中去。
VI
体力运动伴有动脉血压的升高。我们业已知道,动脉内压力是向内流入的推力与向外流出的阻力二者之间相互作用的结果。在其他条件相同时,从心脏的输出量的大量增加本身就能使血压升高。但有证据说明,特别是腹部器官的大面积血管在交感系统血管舒缩神经作用下所发生的收缩,也引起外周阻力的增高。心脏和血管这两个因素的共同作用,把推动血流穿行毛细血管的压力提到相当的高度。让一个人骑在固定脚踏车上进行的试验中,动脉血压从开始时130毫米汞柱升高到180,并且在继续运动过程中此高度保持在165—170毫米之间,这个高度相当于一个大约8英尺的血柱高度,而在休息时其压力只相当于五又二分之一英尺。
值得注意的是,动脉中血压在运动实际进行之前就已升高,恰如呼吸和心率在活动开始时的增加是由于引起该活动的神经冲动所伴随的一种间接效应一样,血压在开始活动时迅速而突然的升高可能也是相同原因所造成。的确,根据威伯(Weber)的说法,仅仅从事运动的念头就能引起腹部内脏的缩小和四肢的胀大。
关于血压升高所具有的意义,我们只要想到活动肌肉中出现小动脉和毛细血管扩张这一点就能最恰当地作出评价。如果动脉压仅仅足以使血循环速度保持在适应躯体静止状态的水平,比如刚超过临界水平,那么任何特殊部分的血管扩张都将为血液流入静脉提供一个方便途径,以致血液转而流向这条较宽的通路,而离开了没有得到充分供血的其他部位。在剧烈活动中升高的动脉压,不仅能防止不活动部分任何这样的供血不足,而且还保证大量血液能快速流过那些最需要带氧血液的活动部分中扩张的血管。
活动肌肉中小动脉和毛细血管的扩张,是为供养细胞并运走废物的最显著的应急措施之一。周密的研究证明,在肌肉休息时,其中许多毛细血管并未被使用,也就是说它们是交替地让血液通过的;某些血管开放一段时间,然后关闭,不让血液流过,同时其他附近的毛细血管开放,以代替相邻部分的工作。在显微镜下只能看到含有血液的毛细血管。丹麦生理学家克罗格(Krogh)对躯体一侧活动肌肉与对侧不活动肌肉作了比较后发现了一个惊人的事实:在活动肌肉中开放的毛细血管数目为静止肌肉中的40倍到100倍。还不了解是什么因素引起毛细血管扩张。人们作过推测:局部缺氧或二氧化碳增加是引起血管开放的原因,即利用这种办法来摆脱自己的不利状态。也可能这种扩张是由于肌肉受牵拉时的磨损和撕裂所产生一种微妙的物质作用所引起的。
无论毛细血管是怎样得到开放的,都不能忽视这种开放的高度重要性。我在前面强调过这一点,即血液和固定细胞间的物质交换是在毛细血管部分发生的。循环系统对体力工作的所有适应作用在此都有自己的意义。血液运给工作中的肌肉所需要的糖和氧,并能把收缩过程中氧化所生成的二氧化碳和水带走。流动血液越是能靠近具有这两种需要的肌细胞,肌肉工作就越能有效地进行。在肌肉开始行动时,肌肉中那些未使用的毛细血管明显的开放,保证了细胞与血流间紧密的联系。
我们现在可以对循环系统中的适应性变化的过程作个结束了。显然,当肌肉有节律地收缩并压迫肌内和肌间的血管时,它所推动的血液量要比在它静止状态下的为多。总之,工作中肌肉所起的作用宛如设置在另外部位的心脏,在工作时接受较多的血液,而又把这些血液送回中心部位的心脏以及肺内,使血液更新以便完成新的任务。
VII
我们现在已注意到,为满足对氧的额外需求,肺每分钟换气量可能增加6倍,心脏搏出量约增加1倍,心率增加1倍,并增加动脉压,使得每分钟内有更大量的血液通过增加了需氧量的活动器官中的已扩张的毛细血管。另外还有两个相当值得注意的机制有待提及。第一个是,旨在保持稳态而运用加速作用过程的另一例证。它从肺和收缩肌肉中毛细血管内气体的便利交换可以看出。
上面概要谈到的循环调节,其作用在于增加红细胞在肺与肌肉间在给定时间内的循环次数。虽然血流通过毛细血管时比通过循环中任何其他部分要慢得多——这是为了保证血液与组织间进行交换所需要的时间,但在循环速度加快时,毛细血管内的速度也能和别处一样得到加快。这意味着载体在肺部释放二氧化碳和装载氧以及在肌肉做工部分进行相反作用所能得到的时间都将减少。小井沼(Oinuma)[2]以出色的观察所获事实证明,二氧化碳的增加,还有体温的升高,都能促进氧的解离。由此可知,在肌肉反复牵引中不仅产生额外的二氧化碳,还产生额外的热量,这些新的条件促使载体更快、更完全地抛出氧。甚至在中等程度的运动中也可因CO2的作用再加上活动肌肉中温度升高的作用而使氧的释放速度加倍。同时,细胞以更快的速度利用氧气,而与其直接接触的淋巴液中的氧量则减少。这造成了从血液向淋巴扩散倾向的增加,其结果是发生了更快的扩散。值得注意的是,这些作用间存在着共同变化的关系,即肌肉工作强度越大,则对氧从血细胞解离并进入细胞速度所产生的效应也越大。因此血流通过肌肉毛细血管速度的加快和流量的加大,给机体的适应带来了益处。
氧的解离速度越快,它在血浆中的扩散压将越高,因此按照顺序,它将更快地进入淋巴,因而也更快地进入活动细胞。作为这些作用的连锁反应的结果,细胞按照其不断变化着的需要来接受氧气。
在肺的毛细血管中,血液处在氧的相对高的扩散压和二氧化碳的相对低的扩散压之下,因为这两种气体都和肺泡有关联。其结果是,二氧化碳穿过组成毛细血管壁和肺泡壁的极薄的膜进入肺泡腔。但二氧化碳不受帮助就不能自行离去。从肺泡进入红细胞的氧能比其他方式更快地帮助驱出二氧化碳。因此在循环途中的每一个转换站,一种气体赶出其他气体后就为自己占有了后者在载体中的空位,直到下一个交替中自己又被照样赶出。在机体中任何部分都找不到比这种过程更为生动的相互作用了。
血液在离开肺时,甚至在剧烈体力活动下的快速流动中也能带走占其全部载氧量95%的氧,也就是大约每百立方厘米血液中18立方厘米的氧。在安静的休息状态下,每100cc静脉血在返回心脏时可带回14或15cc的氧。在外周毛细血管中只放出了3或4cc。由于我们刚才讨论过的那些因素作用的效果,在活动部位中血液氧容量的下降大大增加,因而即使是混合的静脉血[3],在其返回时,氧含量不到5cc%。因此血细胞从肺到需氧组织间循环次数的增加是必须依靠携带它的氧气在利用率上的进一步提高来扩大效果。
VIII
在遇到需要时保证充足供氧的一个更明显的措施见于肌肉运动期间红细胞数量的骤然增加。这种主要由巴克洛夫特所阐明的现象,在低等动物中比在人类更为突出。马在5分钟重度活动后每立方毫米红细胞数量可增加20%或更多一些。这种显著变化是机体对氧的需要所产生的唯一类似储存方式的适应作用,关于这种储存方式我已在氧以外其他物质的稳态中叙述过了。在繁重和长时间的肌肉劳动中,如我们所知,葡萄糖从肝储备中被动员出来并通过血液输送到任何需要的部位以供利用。从机体对失血的反应这个角度来看,脾被认为是浓缩集存红细胞的一个贮藏所。如遇到引起交感—肾上腺系统产生作用的处境,例如一旦发生缺氧,而使脾肌肉产生收缩,则其中内容物就被挤出。在猫身上,运动可使脾在一次释放后其重量从26克减至13克,可知有13克是特别富含红细胞的体液。自然,这些血细胞在面临要求发挥作用的时刻立即就变成了氧和二氧化碳的载体。
为保持对机体中固定细胞和远处细胞的氧的输送的稳态,存在着上述这些多种多样的精密安排的机制,这一点反映了保持一个不断适应需求的输送作用的重要性。在所有中度和长时间的劳动中,在参与这种所必需的职能的多数因子的密切合作下,才有效地维持了充足供氧。但在剧烈和极重的劳动中,这种供给可能尚不足以满足其要求。这时非挥发性乳酸开始在细胞内蓄积并向外扩散入淋巴和血液。但血液的酸碱度应处于酸性和碱性反应之问并接近中性的状态,这一点是不允许改变的。关于保护机体免于遭到这种危险的一些防备措施,我们将在下面加以考察。
参考文献
Bainbridge. Journ. Physiol., 1914, xlviii, 332.
Barcroft. Ergebn. d. Physiol., 1926, xxv, 818.
Cannon and Britton, Am. Journ. Physiol., 1927, lxxix, 433.
Cannon, Linton and Linton. Ibid., 1924, lxxi, 153.
Henderson. Physiol. Rev., 1923, iii, 165.
Hill, Long and Lupton. Proc. Roy. Soc., London, 1924, xcvi, 442.
Johanson. Skand. Arch. f. Physiol., 1895, v, 59.
Krogh. The Anatomy and Physiology of the Capillaries. New Haven,1929, 64.
Mathison. Journ. Physiol., 1911, xlii, 283.
Oinuma. Ibid., 1911, xliii, 364.
Starling. The Law of the Heart. London, 1918.
Weber. Arch. f. Physiol., 1907, 300.
[1] 1夸脱(quart)约等于1.14公升。——译者
[2] 小井沼为原书上Oinuma的日语音译。——译者
[3] 指来自活动部位与非活动部位混合的静脉血。——译者
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。












